
Host Non-Coding RNA Regulates Influenza A Virus Replication

 ,
,
Abstract
:1. Introduction
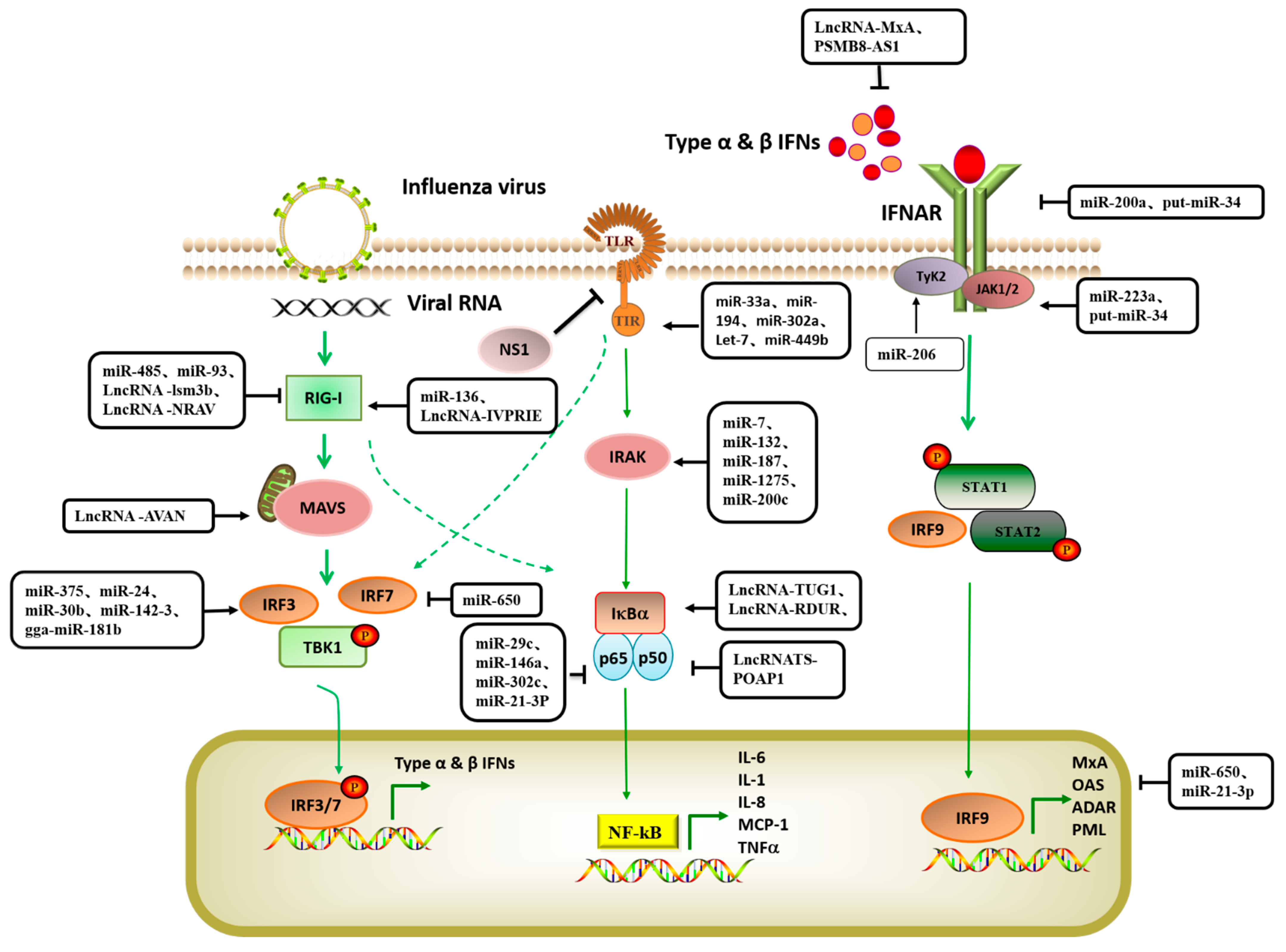
2. Host miRNA Regulates IAV Replication
2.1. Host miRNA Directly Inhibits IAV Gene Expression

{kind=link}
{kind=link}
| miRNA | Target a | Cell Type | Virus | Induced Expression b | Binding Site of Target c | References |
|---|---|---|---|---|---|---|
| miR-323 miR-491 miR-654 | PB1 | MDCK | H1N1 (A/WSN/33) | UP | CCACC | [28] |
| miR-3145 | A549 | H5N1 (A/Thailand/NK165/2005) H1N1 (A/Thailand/104/2009) H3N2 (A/Thailand/ CU-H1817/2010) | UP | UAUGGAGCUGCCC GCUUUGGAGUGUC UUUGGAGUGUCU | [27] | |
| miR-485 | HEK293T | H5N1 (A/duck/India/02CA10/2011) | UP | CAGCCUC | [31] | |
| miR-324-5p | A549 | H5N1 (A/duck/India/02CA10/2011) | DOWN | GAGGGGAT | [39] | |
| gga-miR6675 | HEK293T | H9N2 (A/NJO2/2009) | UP | — | [26] | |
| miR-4753 | A549 | H5N1 (A/Thailand/NK165/2005) | UP | AGAGAAAGAGAA | [27] | |
| miR-584-5p | PB2 | A549 | H1N1 (A/Beijing/501/2009) H5N1 (A/goose/Jilin/hb/2003) | DOWN | GCAAACCA GGAGGGC | [30] |
| miR-1249 | ||||||
| miR-188-3p | A549 | H1N1(A/FM/1/47) H5N6 (A/chicken/Hubei/XY918/2016) H7N9 (A/quail/Hebei/CH06–07/2018) | UP | TGTGGGA ATGTGGGA ATGTGGGA | [29] | |
| hsa-miR-1307-3p | NS1 | A549 | H1N1 (A/California/04/2009) | UP | CGCCGAG | [33] |
| miR-3682 | A549 | H1N1 (A/Thailand/104/2009) H3N2 (A/Thailand/ CU-H1817/2010) | UP | GUAUCGUC AUGAUAACACAG | [27] | |
| miR-4331 | NPTR | H1N1 | DOWN | TGGCCT ACAGCCAC | [40] | |
| miR-584-5P | M1 | A549 | H5N1 (A/goose/Jilin/hb/2003) | UP | GCAAACCA | [30] |
| let-7c | A549 | H1N1 (Jing Fang/86-1) | UP | ACTACCT | [37] | |
| miR-204 | HA | NPTR | H1N1 (A/swine/Nanchang/F9/2010) | DOWN | AAAGGGA | [40] |
| miR-192 | A549 | H5N1 (A/PR8) | UP | — d | [41] | |
| ssc-miR-221-3p | PAM | H5N1 (A/duck/An-hui/1/2006) | DOWN | AAUGUGGUA | [38] | |
| miR-1249 | NA | A549 | H5N1 (A/goose/Jilin/hb/2003) H1N1 (A/Beijing/501/2009) | UP | GGAGGGC | [30] |
| miR-216b | A549 | H5N1 (A/Thailand/NK165/2005) H3N2 (A/Thailand/ CU-H1817/2010) | UP | UGCAGGGAU UGUCUGCAGAGA | [27] | |
| miR-4513 | PA | A549 | H1N1 (A/Thailand/104/2009) H3N2 (A/Thailand/ CU-H1817/2010) | UP | UCGUCAGUC | [27] |
| miR-5693 | A549 | H5N1 (A/Thailand/NK165 /2005) H3N2 (A/Thailand /CU-H1817/2010) | UP | UAGAGCCACUG AGAGAAAGAGAA | [27] | |
| ssc-miR-222 | HEK293T | H5N1 (A/duck/An-hui/1/2006) | UP | GAUGUGGUA | [38] |
2.2. Host miRNAs Indirectly Inhibit IAV Replication by Regulating Intracellular Signaling
2.3. Host miRNAs Promote the Replication of IAV
| Signaling Pathways a | Target Gene b | miRNA | Cell Type | Regulation Direction c | Virus | Induced Expression d | References |
|---|---|---|---|---|---|---|---|
| RLRS | RIG-1 | miR-485 | HEK293T | Promote | H5N1 (A/duck/India/02CA10/2011) | UP | [31] |
| RIG-1, IL-6 | miR-136 | A549 | Inhibition | H5N1 (A/duck /Hubei /hangmei01/2006) | UP | [56] | |
| RIG-1 | miR-93 | A549 | Promote | H1N1 (A/ Puerto Ri-co/8/1934) | DOWN | [64] | |
| TLRS | ARCN1 | miR-33a | A549, Hela | Inhibition | H1N1 (A/WSN/33) H9N2 (A/Chicken/Liaoning/1/100) | UP | [66] |
| FGF2 | miR-194 | A549 | Inhibition | H1N1 (IAV/ Beijing/501/2009) | UP | [67] | |
| IRF5 | miR-302a | A549 | Inhibition | H1N1 (A/FM/1/47) | DOWN | [59] | |
| IRAK1, MAPK3 | miR-7, miR-132, miR-187, miR-1275, miR-200c | A549 | Inhibition | H1N1 (A/WSN/33) | UP | [68] | |
| HDAC1 | miR-449b | A549 | Inhibition | H1N1 (A/WSN/33) | UP | [69] | |
| TOLL 4 | Let-7e | THP-1 | Inhibition | H7N9 (A/Anhui/1/2013) | UP | [42] | |
| NF-κB | IRAK-1, TRAF6 | miR-146a | Hela | Inhibition | H1N1 (A/Jing fang/01/1986) H3N2 (A/Lu fang/09/ 1993) | UP | [70] |
| TRAF6 | miR-144 | HBE | Inhibition | H1N1 (A/ Puerto Ri-co/8/34) | UP | [71] | |
| NFKBIB | miR-4776 | MDCK | Inhibition | H1N1 (A/WSN/33) | UP | [72] | |
| BCL2L2 | miR-29c | A549 | Inhibition | H1N1 (A/Jing fang/01/1986) | UP | [44] | |
| NIK | miR-302c | A549 | Promote | H3N2 (A/Hong Kong/498/97) | DOWN | [57] | |
| TRAF6 | miR-146a | A549 | Promote | H1N1 (A/Jing fang/01/ 1986) H3N2 (A/Lu fang/09/1993) | UP | [70] | |
| A20 | miR-29c | A549 | Promote | H1N1 (A/Jingfang/01/1986) | UP | [44] | |
| HDACB | miR-21-3P | A549 | Promote | H5N1 (A/goose/Ji-Lin/ hb/2003) H1N1 (A/Beijing/501/2009) | DOWN | [63] | |
| FGF2 | Promote | H5N1 (A/Hong Kong/156/97) | DOWN | [62] | |||
| IRF3, IRF7 | miR-24, gga-miR-30b, miR-142-3 | A549 | Inhibition | H9N2 (environment/HN/1-18/2007) | UP | [73] | |
| miR-375, gga-miR-181b | Chicken | Inhibition | H9N2 (environment/HN/1-18/2007) | UP | |||
| USP3 | miR-26a | MDCK | Inhibition | H1N1 (A/WSN/33) | UP | [74] | |
| Jak-STAT | STIM1 | miR-223a | A549 | Inhibition | H1N1 (A/Puerto Rico/8/34) | UP | [60] |
| IFNAR1, STAT2 | miR-200a | A549 | Promote | H1N1 (r1918 and A/Texas/36/91) | DOWN | [43] | |
| CUEDC2 | miR-324-5p | A549 | Inhibition | H5N1 (A/duck/India/02CA10/2011) | DOWN | [39] | |
| IRF3, IFIT2, MxA | miR-650 | NK | Promote | H1N1 (A/Puerto Rico/8/34) | DOWN | [45] | |
| JNK/c-Jun | miR-206 | A549 | Inhibition | H1N1 (A/ Puerto Rico/8/34) H3N2 (A/Oklahoma/309/2006) | DOWN | [46] | |
| STAT3 | put-miR-34 | HBE | Inhibition | H9N1 (1WF10) | DOWN | [75] | |
| Apoptosis | C0X6C | miR-4276 | A549 | Inhibition | H1N1 (A/WS/33) H3N2 (A/Aichi/2/68) | UP | [76] |
| HMBOX1 | ssc-miR-221-3p ssc-miR-222 | PAM | Inhibition | H5N1 (A/duck/Anhui/1/2006) | UP | [38] | |
| MCPIP1 | miR-9 | A549 | Promote | H1N1 (A/PR/8/34) H3N2 (A/Lufang/9/93) | UP | [77] | |
| IL-6 | miR-let-7b-MRE | HBE | Inhibition | H1N1 (A/Nanjing/NJU-108/2009) | UP | [78] | |
| MAPK/ERK | Vimentin | miR-1290 | A549 | Promote | H1N1 (A/Taiwan/126/2009) | UP | [79] |
| Muscleblind | Mbnl3 | miR-674 | Dendritic | Inhibition | H9N2 (A/duck/ Nanjing /01/1999) | UP | [80] |
| Wnt/β-catenin | frizzled 5 gene | miR-29a | HEK293T A549 | Inhibition | H3N2 (A/Oklahoma//309/2006) | UP | [61] |
| DC/TCell | DR1 | miR-203 | A549 | Inhibition | H5N1 (A/Vietnam/1194/2004 | UP | [81] |
| mTOR | mTOR | miR-101 | A549 | Inhibition | H5N1 (A/Hatay/2004) | UP | [82] |
3. Host LncRNAs Regulates IAV Replication
3.1. Host LncRNAs Directly Act on Viral Genes to Regulates Influenza Virus Replication
3.2. Host LncRNAs Indirectly Inhibit IAV Replication by Regulating Intracellular Signals
3.3. Host LncRNAs Indirectly Promote IAV Replication by Regulating Intracellular Signaling
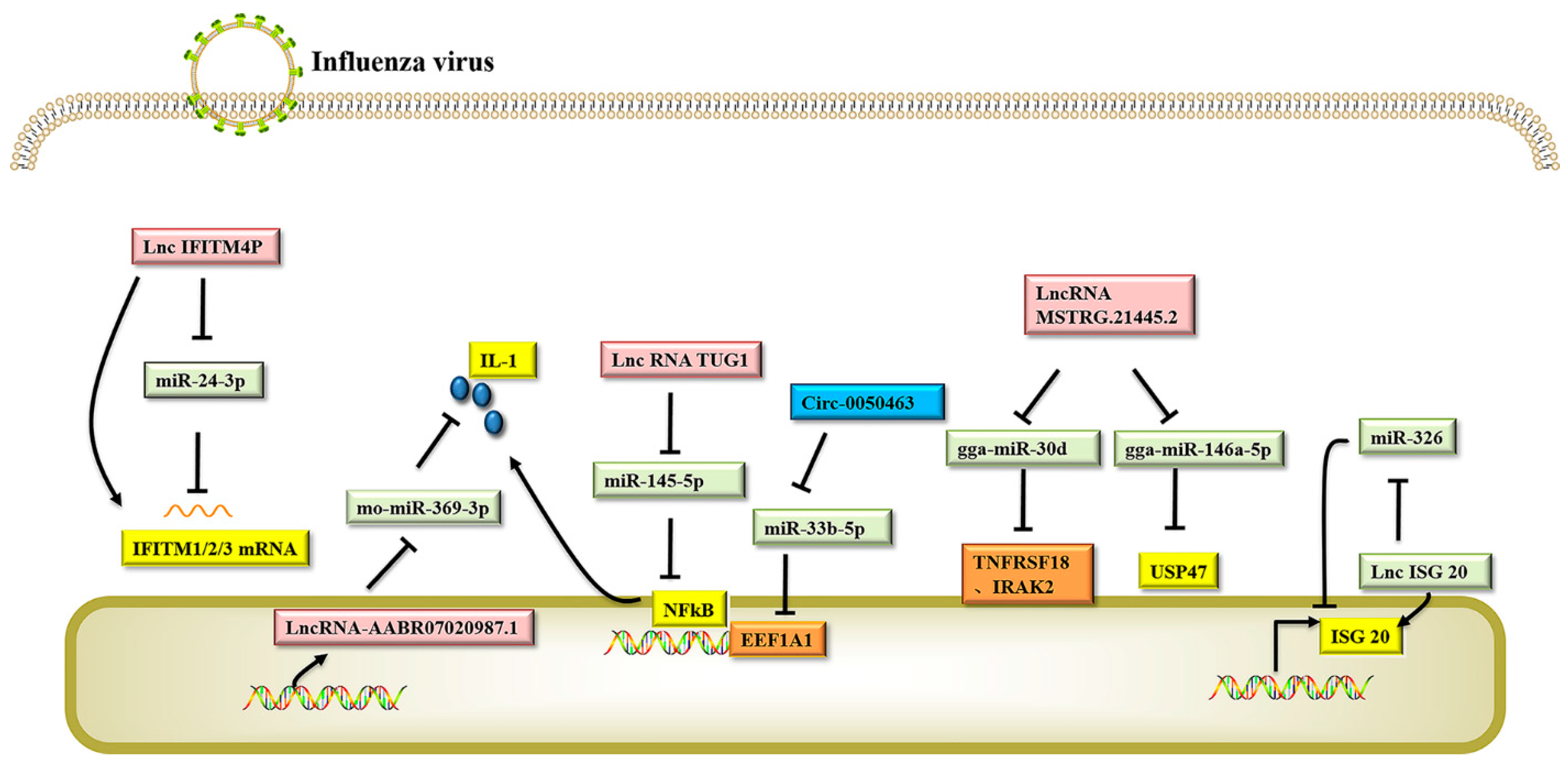
4. Other Host Non-Coding RNAs Regulate the Replication of Influenza A Virus
| LncRNA | Target Gene a | Cell Type | Regulation Direction b | Virus | Induced Expression c | References | |
|---|---|---|---|---|---|---|---|
| LncRNA IPAN | PB1 | A549 | Promote | H1N1 (A/WSN/33) | UP | [86] | |
| LncRNA PAAN | PA | A549 | Promote | H1N1 (A/PR/8/34) | UP | [85] | |
| Signaling pathways d | LncRNA | Target gene e | Cell type | Regulation direction b | Virus | Induced expression c | References |
| RLRs | LncRNA NRAV | ZONAB | A549 | Promote | H1N1 (A/WSN/33) | DOWN | [115] |
| LncRNA Lsm3b | RIG-I | L929 | Promote | H1N1 (A/ Puerto Rico /8/1934) | UP | [104] | |
| LncRNA VIN | Nuclear | A549 | Promote | H1N1 (A/WSN/33) | UP | [103] | |
| LncRNA-155 | PTP1B | A549 | Inhibition | H1N1 (A/Puerto Ri-co/8/1934) | UP | [89] | |
| LncRNA IVPRIE | RIG-I | A549 | Inhibition | H1N1 (A/Puerto Ri-co/8/1934) | UP | [93] | |
| LncRNA NEAT1 | SFPQ | Hela | Inhibition | H1N1 (A/WSN/33) | UP | [116] | |
| LncRNA ISG20 | miR-326 | A549 HEK293T | Inhibition | H1N1 (A/Puerto Ri-co/8/34) | UP | [117] | |
| LncRNA ISR | IFN-β | A549 | Inhibition | H1N1 (A/California/04/2009) | UP | [90] | |
| LncRNA AVAN | TRIM25 | A549 | Inhibition | H7N9 (A/Hebei/01/2013) | UP | [92] | |
| lncRNA IFITM4P | miR-24-3p | A549 | Inhibition | H1N1 (A/Shanghai-Jiading/SWL1970/2015) | UP | [118] | |
| TLRs | LncRNA TSPOAP1 | NF-κB | A549 | Promote | H1N1 (A/PR/8/34) H3N2 (A/Lufang/9/93) | UP | [119] |
| LncRNA-MxA | IFN-β | A549 | Promote | H1N1 (A/WSN/33) | UP | [100] | |
| PSMB8-AS1 | IFN | A549 | Promote | H1N1 (A/Puerto Ri-co/8/34) | UP | [99] | |
| Cell metabolism | LncRNA-ACOD1 | GOT2 | A549 | Promote | H1N1 (A/WSN/33) | UP | [106] |
| NF-κB | LncRNA TUG1 | miR-145-5p | DHBE | Inhibition | H3N2 | UP | [120] |
| LncRNA RDUR | RIG-I/MAVS /NF-κB | A549 | Inhibition | H1N1 (A/Shanghai-Jiad-ing/SWL1970/2015) | UP | [94] | |
| Other ncRNAs in the host regulate influenza A virus replication | |||||||
| Signaling pathways d | NcRNA | Target gene e | Cell type | Regulation direction b | Virus | Induced expression c | References |
| Post transcriptional regulation | Circ_0050463 | microRNA-33b-5p | A549 | Promote | H1N1 (A/Lufang/9/93) | UP | [121] |
| Autophagy regulation | Circ-GATAD2A | VPS34 | A549 | Promote | H1N1 (A/ Puerto Ri-co /8/34) | UP | [114] |
| PKR | vtRNAs | NS1 | A549 | Promote | H1N1 (A/WSN/33) | UP | [108] |
5. Host Non-Coding RNAs Regulate the Replication of Other Influenza Virus
| miRNA | Target Gene a | Cell Type | Regulation Direction b | Virus | Induced Expression c | Binding Site of Target d | References |
|---|---|---|---|---|---|---|---|
| hsa-miR-30e-3p | NA, NP | A549 | Inhibition | IBV (Influ-enza B virus B/Thailand/CU-B5522/2011 representing the Vic-toria lineage) | UP | GAUGUCU GAACUGAAA | [122] |
| Signaling pathways e | miRNA | Target gene f | Cell type | Regulation direction b | Virus | Induced expression c | References |
| TLRs | cfa-miR-197 | KPNA6 | MDCK | Inhibition | IBV Victoria lineage (B/Thailand/CU-B5522/2011), or Yamagata lineage (B/Massa Chu-setts/2/2012) | UP | [128] |
| gga-miR-30d | USP47 | HD11 | Inhibition | IBV (Beaudette strain) | DOWM | [124] | |
| gga-miR-146a-5p | IRAK2, TNFRSF18 | chickens | promote | IBV (Beaudette strain) | UP | [123] |
6. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Ciminski, K.; Chase, G.P.; Beer, M.; Schwemmle, M. Influenza A Viruses: Understanding Human Host Determinants. Trends Mol. Med. 2021, 27, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Dadonaite, B.; Gilbertson, B.; Knight, M.L.; Trifkovic, S.; Rockman, S.; Laederach, A.; Brown, L.E.; Fodor, E.; Bauer, D.L.V. The structure of the influenza A virus genome. Nat. Microbiol. 2019, 4, 1781–1789. [Google Scholar] [CrossRef]
- Harrington, W.N.; Kackos, C.M.; Webby, R.J. The evolution and future of influenza pandemic preparedness. Exp. Mol. Med. 2021, 53, 737–749. [Google Scholar] [CrossRef]
- Podstawka, J.; Sinha, S.; Hiroki, C.H.; Sarden, N.; Granton, E.; Labit, E.; Kim, J.H.; Andonegui, G.; Lou, Y.; Snarr, B.D.; et al. Marginating transitional B cells modulate neutrophils in the lung during inflammation and pneumonia. J. Exp. Med. 2021, 218, e20210409. [Google Scholar] [CrossRef]
- Jiang, Z.; Wei, F.; Zhang, Y.; Wang, T.; Gao, W.; Yu, S.; Sun, H.; Pu, J.; Sun, Y.; Wang, M.; et al. IFI16 directly senses viral RNA and enhances RIG-I transcription and activation to restrict influenza virus infection. Nat. Microbiol. 2021, 6, 932–945. [Google Scholar] [CrossRef]
- Vogel, O.A.; Han, J.; Liang, C.; Manicassamy, S.; Perez, J.T.; Manicassamy, B.; Bukreyev, A. The p150 Isoform of ADAR1 Blocks Sustained RLR signaling and Apoptosis during Influenza Virus Infection. PLoS Pathog. 2020, 16, e1008842. [Google Scholar] [CrossRef]
- Kandasamy, M.; Suryawanshi, A.; Tundup, S.; Perez, J.T.; Schmolke, M.; Manicassamy, S.; Manicassamy, B. RIG-I Signaling Is Critical for Efficient Polyfunctional T Cell Responses during Influenza Virus Infection. PLoS Pathog. 2016, 12, e1005754. [Google Scholar] [CrossRef]
- Sun, N.; Li, C.; Li, X.; Deng, Y.; Jiang, T.; Zhang, N.; Zu, S.; Zhang, R.; Li, L.; Chen, X.; et al. Type-IInterferon-Inducible SERTAD3 Inhibits Influenza A Virus Replication by Blocking the Assembly of Viral RNA Polymerase Complex. Cell Rep. 2020, 33, 108342. [Google Scholar] [CrossRef]
- Jung, H.J.; Park, S.H.; Cho, K.M.; Jung, K.I.; Cho, D.; Kim, T.S. Threonyl-tRNA Synthetase Promotes T Helper Type 1 Cell Responses by Inducing Dendritic Cell Maturation and IL-12 Production an NF-κB Pathway. Front. Immunol. 2020, 11, 571959. [Google Scholar] [CrossRef]
- Hornick, E.E.; Dagvadorj, J.; Zacharias, Z.R.; Miller, A.M.; Langlois, R.A.; Chen, P.; Legge, K.L.; Bishop, G.A.; Sutterwala, F.S.; Cassel, S.L. Dendritic cell NLRC4 regulates influenza A virus-specific CD4 T cell responses through FasL expression. J. Clin. Investig. 2019, 129, 2888–2897. [Google Scholar] [CrossRef]
- Mohapatra, S.; Pioppini, C.; Ozpolat, B.; Calin, G.A. Non-coding RNAs regulation of macrophage polarization in cancer. Mol. Cancer 2021, 20, 24. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Jie, X.; Gao, Y.; Nie, B.; Jiang, H. Long non-coding RNA CACNA1G-AS1 promotes proliferation and invasion and inhibits apoptosis by regulating expression of miR-205 in human keloid fibroblasts. Biosci. Rep. 2020, 40, BSR20192839. [Google Scholar] [CrossRef]
- Lou, M.; Tang, X.; Wang, G.; He, J.; Luo, F.; Guan, M.; Wang, F.; Zou, H.; Wang, J.; Zhang, Q.; et al. Long noncoding RNA BS-DRL1 modulates the DNA damage response and genome stability by interacting with HMGB1 in neurons. Nat. Commun. 2021, 12, 4075. [Google Scholar] [CrossRef]
- Xie, C.; Zhang, L.Z.; Chen, Z.L.; Zhong, W.J.; Fang, J.H.; Zhu, Y.; Xiao, M.H.; Guo, Z.W.; Zhao, N.; He, X.; et al. A hMTR4-PDIA3P1-miR-125/124-TRAF6 Regulatory Axis and Its Function in NF kappa B Signaling and Chemoresistance. Hepatology 2020, 71, 1660–1677. [Google Scholar] [CrossRef]
- Liu, R.; Lu, Z.; Gu, J.; Liu, J.; Huang, E.; Liu, X.; Wang, L.; Yang, J.; Deng, Y.; Qian, J.; et al. MicroRNAs 15A and 16-1 Activate Signaling Pathways That Mediate Chemotaxis of Immune Regulatory B cells to Colorectal Tumors. Gastroenterology 2018, 154, 637–651. [Google Scholar] [CrossRef] [PubMed]
- Gaur, A.B.; Holbeck, S.L.; Colburn, N.H.; Israel, M.A. Downregulation of Pdcd4 by mir-21 facilitates glioblastoma proliferation in vivo. Neuro-oncology 2011, 13, 580–590. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, B.; Bai, Q.; Wang, P.; Wei, G.; Li, Z.; Hu, L.; Tian, Q.; Zhou, J.; Huang, Q.; et al. The lncRNA Snhg1-Vps13D vesicle trafficking system promotes memory CD8 T cell establishment via regulating the dual effects of IL-7 signaling. Signal Transduct. Target. Ther. 2021, 6, 126. [Google Scholar] [CrossRef]
- Khatun, M.; Sur, S.; Steele, R.; Ray, R.; Ray, R.B. Inhibition of Long Noncoding RNA Linc-Pint by Hepatitis C Virus in Infected Hepatocytes Enhances Lipogenesis. Hepatology 2021, 74, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Fan, H.; Chen, X.; Zhao, X.; Wang, X.; Feng, Y.; Liu, M.; Li, S.; Tang, H. Hepatitis B Virus DNA Polymerase Restrains Viral Replication Through the CREB1/HOXA Distal Transcript Antisense RNA Homeobox A13 Axis. Hepatology 2021, 73, 503–519. [Google Scholar] [CrossRef]
- Sui, B.; Chen, D.; Liu, W.; Wu, Q.; Tian, B.; Li, Y.; Hou, J.; Liu, S.; Xie, J.; Jiang, H.; et al. A novel antiviral lncRNA, EDAL, shields a T309 O-GlcNAcylation site to promote EZH2 lysosomal degradation. Genome Biol. 2020, 21, 228. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Xu, X.; Jiang, Y.; Liu, Y.; Sun, W.; Guo, Y.; Ren, Y.; Zuo, W.; Hu, X.; Huang, S.; et al. The endogenous retrovirus-derived long noncoding RNA TROJAN promotes triple-negative breast cancer progression via ZMYND8 degradation. Sci. Adv. 2019, 5, t9820. [Google Scholar] [CrossRef] [Green Version]
- Xue, P.; Huang, S.; Han, X.; Zhang, C.; Yang, L.; Xiao, W.; Fu, J.; Li, H.; Zhou, Y. Exosomal miR-101-3p and miR-423-5p inhibit medulloblastoma tumorigenesis through targeting FOXP4 and EZH2. Cell Death Differ. 2021, 1–14. [Google Scholar] [CrossRef]
- Yu, S.; Kim, V.N. A tale of non-canonical tails: Gene regulation by post-transcriptional RNA tailing. Nat. Rev. Mol. Cell Biol. 2020, 21, 542–556. [Google Scholar] [CrossRef]
- Hanouskova, B.; Vavrova, G.; Ambroz, M.; Bousova, I.; Karlsen, T.A.; Skalova, L.; Matouskova, P. MicroRNAs mediated regulation of glutathione peroxidase 7 expression and its changes during adipogenesis. Biochim. Biophys. Acta Gene Regul. Mech. 2021, 194734. [Google Scholar] [CrossRef]
- Zhang, X.; Chapat, C.; Wang, P.; Choi, J.; Li, Q.; Luo, J.; Wiebe, S.; Kim, S.; Robichaud, N.; Karam, I.F.; et al. microRNA-induced translational control of antiviral immunity by the cap-binding protein 4EHP. Mol. Cell 2021, 81, 1187–1199. [Google Scholar] [CrossRef]
- Lin, J.; Xia, J.; Zhang, T.; Zhang, K.; Yang, Q. Genome-wide profiling of microRNAs reveals novel insights into the interactions between H9N2 avian influenza virus and avian dendritic cells. Oncogene 2018, 37, 4562–4580. [Google Scholar] [CrossRef]
- Khongnomnan, K.; Makkoch, J.; Poomipak, W.; Poovorawan, Y.; Payungporn, S. Human miR-3145 inhibits influenza A viruses replication by targeting and silencing viral PB1 gene. Exp. Biol. Med. 2015, 240, 1630–1639. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Liu, H.; Gao, S.; Jiang, W.; Huang, W. Cellular MicroRNAs Inhibit Replication of the H1N1 Influenza A Virus in Infected Cells. J. Virol. 2010, 84, 8849–8860. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Zhang, C.; Zhao, Z.; Zhang, C.; Fu, Y.; Li, J.; Chen, G.; Lai, M.; Li, Z.; Dong, S.; et al. Identification of cellular microRNA miR-188-3p with broad-spectrum anti-influenza A virus activity. Virol. J. 2020, 17, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Zhang, Y.Y.; Lu, J.S.; Xia, B.H.; Yang, Z.X.; Zhu, X.D.; Zhou, X.W.; Huang, P.T. The highly pathogenic H5N1 influenza A virus down-regulated several cellular MicroRNAs which target viral genome. J. Cell. Mol. Med. 2017, 21, 3076–3086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingle, H.; Kumar, S.; Raut, A.A.; Mishra, A.; Kulkarni, D.D.; Kameyama, T.; Takaoka, A.; Akira, S.; Kumar, H. The microRNA miR-485 targets host and influenza virus transcripts to regulate antiviral immunity and restrict viral replication. Sci. Signal. 2015, 8, a126. [Google Scholar] [CrossRef]
- Jiang, W.; Wang, Q.; Chen, S.; Gao, S.; Song, L.; Liu, P.; Huang, W. Influenza A virus NS1 induces G0/G1 cell cycle arrest by inhibiting the expression and activity of RhoA protein. J. Virol. 2013, 87, 3039–3052. [Google Scholar] [CrossRef] [Green Version]
- Bavagnoli, L.; Campanini, G.; Forte, M.; Ceccotti, G.; Percivalle, E.; Bione, S.; Lisa, A.; Baldanti, F.; Maga, G. Identification of a novel antiviral micro-RNA targeting the NS1 protein of the H1N1 pandemic human influenza virus and a corresponding viral escape mutation. Antivir. Res. 2019, 171, 104593. [Google Scholar] [CrossRef]
- Zhou, Z.; Li, X.; Liu, J.; Dong, L.; Chen, Q.; Liu, J.; Kong, H.; Zhang, Q.; Qi, X.; Hou, D.; et al. Honeysuckle-encoded atypical microRNA2911 directly targets influenza A viruses. Cell Res. 2015, 25, 39–49. [Google Scholar] [CrossRef]
- Williams, G.D.; Townsend, D.; Wylie, K.M.; Kim, P.J.; Amarasinghe, G.K.; Kutluay, S.B.; Boon, A.C.M. Nucleotide resolution mapping of influenza A virus nucleoprotein-RNA interactions reveals RNA features required for replication. Nat. Commun. 2018, 9, 412–465. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Pang, M.; Liang, S.; Lin, Y.; Zhao, Y.; Qiu, D.; Liu, J.; Dong, Y.; Liu, Y. Cellular microRNAs influence replication of H3N2 canine influenza virus in infected cells. Vet. Microbiol. 2021, 257, 109083. [Google Scholar] [CrossRef]
- Ma, Y.J.; Yang, J.; Fan, X.L.; Zhao, H.B.; Hu, W.; Li, Z.P.; Yu, G.C.; Ding, X.R.; Wang, J.Z.; Bo, X.C.; et al. Cellular microRNA let-7c inhibits M1 protein expression of the H1N1 influenza A virus in infected human lung epithelial cells. J. Cell. Mol. Med. 2012, 16, 2539–2546. [Google Scholar] [CrossRef]
- Song, J.; Sun, H.; Sun, H.; Jiang, Z.; Zhu, J.; Wang, C.; Gao, W.; Wang, T.; Pu, J.; Sun, Y.; et al. Swine MicroRNAs ssc-miR-221-3p and ssc-miR-222 Restrict the Cross-Species Infection of Avian Influenza Virus. J. Virol. 2020, 94, e01700-20. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, A.; Ingle, H.; Kumar, S.; Mishra, R.; Verma, M.K.; Biswas, D.; Kumar, N.S.; Mishra, A.; Raut, A.A.; et al. MicroRNA hsa-miR-324-5p Suppresses H5N1 Virus Replication by Targeting the Viral PB1 and Host CUEDC2. J. Virol. 2018, 92, e01057-18. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wang, R.; Su, H.; Wang, B.; Sizhu, S.; Lei, Z.; Jin, M.; Chen, H.; Cao, J.; Zhou, H. Sus scrofa miR-204 and miR-4331 Negatively Regulate Swine H1N1/2009 Influenza A Virus Replication by Targeting Viral HA and NS, respectively. Int. J. Mol. Sci. 2017, 18, 749. [Google Scholar] [CrossRef] [PubMed]
- Langlois, R.A.; Albrecht, R.A.; Kimble, B.; Sutton, T.; Shapiro, J.S.; Finch, C.; Angel, M.; Chua, M.A.; Gonzalez-Reiche, A.S.; Xu, K.; et al. MicroRNA-based strategy to mitigate the risk of gain-of-function influenza studies. Nat. Biotechnol. 2013, 31, 844–847. [Google Scholar] [CrossRef]
- Zhang, S.; Gu, D.; Ouyang, X.; Xie, W. Proinflammatory effects of the hemagglutinin protein of the avian influenza A (H7N9) virus and microRNAmediated homeostasis response in THP1 cells. Mol. Med. Rep. 2015, 12, 6241–6246. [Google Scholar] [CrossRef]
- Li, Y.U.; Chan, E.Y.; Li, J.; Ni, C.; Peng, X.; Rosenzweig, E.; Tumpey, T.M.; Katze, M.G. MicroRNA Expression and Virulence in Pandemic Influenza Virus-Infected Mice. J. Virol. 2010, 84, 3023–3032. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Dong, C.; Sun, X.; Li, Z.; Zhang, M.; Guan, Z.; Duan, M. Induction of the cellular miR-29c by influenza virus inhibits the innate immune response through protection of A20 mRNA. Biochem. Bioph. Res. Co. 2014, 450, 755–761. [Google Scholar] [CrossRef]
- Pichulik, T.; Khatamzas, E.; Liu, X.; Brain, O.; Delmiro, G.M.; Leslie, A.; Danis, B.; Mayer, A.; Baban, D.; Ragoussis, J.; et al. Pattern recognition receptor mediated downregulation of microRNA-650 fine-tunes MxA expression in dendritic cells infected with influenza A virus. Eur. J. Immunol. 2016, 46, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Bamunuarachchi, G.; Yang, X.; Huang, C.; Liang, Y.; Guo, Y.; Liu, L. MicroRNA-206 inhibits influenza A virus replication by targeting tankyrase 2. Cell. Microbiol. 2021, 23, e13281. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Wu, W.; Jones, C.; Perwitasari, O.; Mahalingam, S.; Tripp, R.A.; Chan, M.C. MicroRNA Regulation of Human Genes Essential for Influenza A (H7N9) Replication. PLoS ONE 2016, 11, e155104. [Google Scholar] [CrossRef] [Green Version]
- Goncalves-Alves, E.; Saferding, V.; Schliehe, C.; Benson, R.; Kurowska-Stolarska, M.; Brunner, J.S.; Puchner, A.; Podesser, B.K.; Smolen, J.S.; Redlich, K.; et al. MicroRNA-155 Controls T Helper Cell Activation During Viral Infection. Front. Immunol. 2019, 10, 1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Tan, S.; Wu, Z.; Xu, L.; Wang, Z.; Xu, Y.; Wang, T.; Gao, C.; Gong, Y.; Liang, X.; et al. HBV suppresses ZHX2 expression to promote proliferation of HCC through miR-155 activation. Int. J. Cancer 2018, 143, 3120–3130. [Google Scholar] [CrossRef]
- Zhu, Z.; Teng, M.; Li, H.; Zheng, L.; Liu, J.; Yao, Y.; Nair, V.; Zhang, G.; Luo, J. Virus-encoded miR-155 ortholog in Marek’s disease virus promotes cell proliferation via suppressing apoptosis by targeting tumor suppressor WWOX. Vet. Microbiol. 2021, 252, 108919. [Google Scholar] [CrossRef]
- Shen, S.M.; Jiang, H.; Zhao, J.N.; Shi, Y. Down-regulation of miR-155 inhibits inflammatory response in human pulmonary microvascular endothelial cells infected with influenza A virus by targeting sphingosine-1-phosphate receptor 1. Chin. Med. J. 2020, 133, 2429–2436. [Google Scholar] [CrossRef]
- Woods, P.S.; Doolittle, L.M.; Rosas, L.E.; Nana-Sinkam, S.P.; Tili, E.; Davis, I.C. Increased expression of microRNA-155-5p by alveolar type II cells contributes to development of lethal ARDS in H1N1 influenza A virus-infected mice. Virology 2020, 545, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Izzard, L.; Dlugolenski, D.; Xia, Y.; Mcmahon, M.; Middleton, D.; Tripp, R.A.; Stambas, J. Enhanced immunogenicity following miR-155 incorporation into the influenza A virus genome. Virus Res. 2017, 235, 115–120. [Google Scholar] [CrossRef]
- Sayed, N.; Ospino, F.; Himmati, F.; Lee, J.; Chanda, P.; Mocarski, E.S.; Cooke, J.P. Retinoic Acid Inducible Gene 1 Protein (RIG1)-Like Receptor Pathway Is Required for Efficient Nuclear Reprogramming. Stem Cells 2017, 35, 1197–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, J.; Zheng, Y.; Mo, J.; Wang, W.; Luo, Z.; Li, Y.; Chen, X.; Zhang, Q.; Wu, K.; Liu, W.; et al. Sox4 represses host innate immunity to facilitate pathogen infection by hijacking the TLR signaling networks. Virulence 2021, 12, 704–722. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhu, J.; Zhou, H.; Zhao, Z.; Zou, Z.; Liu, X.; Lin, X.; Zhang, X.; Deng, X.; Wang, R.; et al. Identification of cellular microRNA-136 as a dual regulator of RIG-I-mediated innate immunity that antagonizes H5N1 IAV replication in A549 cells. Sci. Rep. 2015, 5, 14991. [Google Scholar] [CrossRef] [Green Version]
- Gui, S.; Chen, X.; Zhang, M.; Zhao, F.; Wan, Y.; Wang, L.; Xu, G.; Zhou, L.; Yue, X.; Zhu, Y.; et al. Mir-302c mediates influenza A virus-induced IFNβ expression by targeting NF-κB inducing kinase. FEBS Lett. 2015, 589, 4112–4118. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhou, L.; Peng, N.; Yu, H.; Li, M.; Cao, Z.; Lin, Y.; Wang, X.; Li, Q.; Wang, J.; et al. MicroRNA-302a suppresses influenza A virus–stimulated interferon regulatory factor-5 expression and cytokine storm induction. J. Biol. Chem. 2017, 292, 21291–21303. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Yu, S.; Ren, P.; Sun, X.; Jin, M. Human microRNA-30 inhibits influenza virus infection by suppressing the expression of SOCS1, SOCS3, and NEDD4. Cell. Microbiol. 2020, 22, e13150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Miao, Y.; Chen, R.; Zhang, Y.; Wu, H.; Yang, S.; Shang, L. STIM1 mediates IAV-induced inflammation of lung epithelial cells by regulating NLRP3 and inflammasome activation via targeting miR-223. Life Sci. (1973) 2021, 266, 118845. [Google Scholar] [CrossRef]
- Yang, X.; Liang, Y.; Bamunuarachchi, G.; Xu, Y.; Vaddadi, K.; Pushparaj, S.; Xu, D.; Zhu, Z.; Blaha, R.; Huang, C.; et al. miR-29a is a negative regulator of influenza virus infection through targeting of the frizzled 5 receptor. Arch. Virol. 2021, 166, 363–373. [Google Scholar] [CrossRef]
- Shi, J.; Feng, P.; Gu, T. MicroRNA-21-3p modulates FGF2 to facilitate influenza A virus H5N1 replication by refraining type I interferon response. Biosci. Rep. 2020, 40, BSR20200158. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Lu, J.; Wang, R.; Yang, Z.; Zhou, X.; Huang, P. miR-21-3p Regulates Influenza A Virus Replication by Targeting Histone Deacetylase-8. Front. Cell. Infect. Microbiol. 2018, 8, 175. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Li, F.; Ji, J.; Liu, Y.; Liu, F.; Zhao, Y.; Li, J.; Han, S.; Wang, Q.; Ding, G. Inhibition of miR-93 promotes interferon effector signaling to suppress influenza A infection by upregulating JAK1. Int. Immunopharmacol. 2020, 86, 106754. [Google Scholar] [CrossRef]
- Liu, S.; Li, H.; Wang, Y.; Li, H.; Du, S.; Zou, X.; Zhang, X.; Cao, B. High Expression of IL-36γ in Influenza Patients Regulates Interferon Signaling Pathway and Causes Programmed Cell Death During Influenza Virus Infection. Front. Immunol. 2020, 11, 552606. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Jiang, L.; Lai, W.; Qin, Y.; Zhang, T.; Wang, S.; Ye, X. MicroRNA-33a disturbs influenza A virus replication by targeting ARCN1 and inhibiting viral ribonucleoprotein activity. J. Gen. Virol. 2016, 97, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Lai, C.; Gu, H.; Zhao, L.; Xia, M.; Yang, P.; Wang, X. miR-194 Inhibits Innate Antiviral Immunity by Targeting FGF2 in Influenza H1N1 Virus Infection. Front. Microbiol. 2017, 8, 2187. [Google Scholar] [CrossRef] [Green Version]
- Buggele, W.A.; Johnson, K.E.; Horvath, C.M. Influenza A virus infection of human respiratory cells induces primary microRNA expression. J. Biol. Chem. 2012, 287, 31027–31040. [Google Scholar] [CrossRef] [Green Version]
- Buggele, W.A.; Krause, K.E.; Horvath, C.M.; Pekosz, A. Small RNA profiling of influenza A virus-infected cells identifies miR-449b as a regulator of histone deacetylase 1 and interferon beta. PLoS ONE 2013, 8, e76560. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Sun, X.; Zhu, Y.; Qin, W. Downregulation of miR-146a inhibits influenza A virus replication by enhancing the type I interferon response in vitro and in vivo. Biomed. Pharmacother. 2019, 111, 740–750. [Google Scholar] [CrossRef]
- Rosenberger, C.M.; Podyminogin, R.L.; Diercks, A.H.; Treuting, P.M.; Peschon, J.J.; Rodriguez, D.; Gundapuneni, M.; Weiss, M.J.; Aderem, A.; Lopez, C.B. miR-144 attenuates the host response to influenza virus by targeting the TRAF6-IRF7 signaling axis. PLoS Pathog. 2017, 13, e1006305. [Google Scholar] [CrossRef]
- Othumpangat, S.; Bryan, N.B.; Beezhold, D.H.; Noti, J.D. Upregulation of miRNA-4776 in Influenza Virus Infected Bronchial Epithelial Cells Is Associated with Downregulation of NFKBIB and Increased Viral Survival. Viruses 2017, 9, 94. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Gao, Q.S.; Zhou, L.; Chen, Z.H.; Lu, S.; Huang, H.J.; Zhan, C.Y.; Xiang, M. MicroRNAs in avian influenza virus H9N2-infected and non-infected chicken embryo fibroblasts. Genet. Mol. Res. 2015, 14, 9081–9091. [Google Scholar] [CrossRef]
- Gao, S.; Li, J.; Song, L.; Wu, J.; Huang, W. Influenza A virus-induced downregulation of miR-26a contributes to reduced IFNalpha/beta production. Virol. Sin. 2017, 32, 261–270. [Google Scholar] [CrossRef]
- Othumpangat, S.; Beezhold, D.H.; Umbright, C.M.; Noti, J.D. Influenza Virus-Induced Novel miRNAs Regulate the STAT Pathway. Viruses 2021, 13, 967. [Google Scholar] [CrossRef] [PubMed]
- Othumpangat, S.; Noti, J.D.; Beezhold, D.H. Lung epithelial cells resist influenza A infection by inducing the expression of cytochrome c oxidase VIc which is modulated by miRNA 4276. Virology 2014, 468, 256–264. [Google Scholar] [CrossRef]
- Dong, C.; Sun, X.; Guan, Z.; Zhang, M.; Duan, M. Modulation of influenza A virus replication by microRNA-9 through targeting MCPIP1. J. Med. Virol. 2017, 89, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Sun, W.; Shi, Y.; Xing, Z.; Su, X.; Cantarini, L. Altered Viral Replication and Cell Responses by Inserting MicroRNA Recognition Element into PB1 in Pandemic Influenza A Virus (H1N1) 2009. Mediat. Inflamm. 2015, 2015, 976512–976575. [Google Scholar] [CrossRef]
- Huang, S.Y.; Huang, C.H.; Chen, C.J.; Chen, T.W.; Lin, C.Y.; Lin, Y.T.; Kuo, S.M.; Huang, C.G.; Lee, L.A.; Chen, Y.H.; et al. Novel Role for miR-1290 in Host Species Specificity of Influenza A Virus. Mol. Ther. Nucleic Acids 2019, 17, 10–23. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Chen, Y.T.; Xia, J.; Yang, Q. MiR674 inhibits the neuraminidase-stimulated immune response on dendritic cells via down-regulated Mbnl3. Oncotarget 2016, 7, 48978–48994. [Google Scholar] [CrossRef]
- Zhang, S.; Li, J.; Li, J.; Yang, Y.; Kang, X.; Li, Y.; Wu, X.; Zhu, Q.; Zhou, Y.; Hu, Y. Up-regulation of microRNA-203 in influenza A virus infection inhibits viral replication by targeting DR1. Sci. Rep. 2018, 8, 6715–6797. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Chatterjee, A.; Kumar, P.; Lal, S.; Kondabagil, K. Upregulation of miR-101 during Influenza A Virus Infection Abrogates Viral Life Cycle by Targeting mTOR Pathway. Viruses 2020, 12, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, T.; Zhang, Q.; Wu, B.; Wang, J.; Li, N.; Zhang, T.; Wang, Z.; Luo, J.; Guo, X.; Ding, X.; et al. LncRNA-encoded microproteins: A new form of cargo in cell culture-derived and circulating extracellular vesicles. J. Extracell Vesicles 2021, 10, e12123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, Y.; Zhou, R.; Zhao, J.; Zhang, Y.; Yi, D.; Li, Q.; Zhou, J.; Guo, F.; Liang, C.; et al. Host Long Noncoding RNA lncRNA-PAAN Regulates the Replication of Influenza A Virus. Viruses 2018, 10, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, Y.; Li, Q.; Zhao, J.; Yi, D.; Ding, J.; Zhao, F.; Hu, S.; Zhou, J.; Deng, T.; et al. Influenza Virus Exploits an Interferon-Independent lncRNA to Preserve Viral RNA Synthesis through Stabilizing Viral RNA Polymerase PB1. Cell Rep. 2019, 27, 3295–3304. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zheng, X.; Li, J.; Wang, G.; Hu, Z.; Chen, Y.; Wang, X.; Gu, M.; Gao, R.; Hu, S.; et al. Long noncoding RNA#45 exerts broad inhibitory effect on influenza a virus replication via its stem ring arms. Virulence 2021, 12, 2443–2460. [Google Scholar] [CrossRef]
- Zong, Z.; Zhang, Z.; Wu, L.; Zhang, L.; Zhou, F. The Functional Deubiquitinating Enzymes in Control of Innate Antiviral Immunity. Adv. Sci. 2021, 8, 2002484. [Google Scholar] [CrossRef]
- Maarouf, M.; Chen, B.; Chen, Y.; Wang, X.; Rai, K.R.; Zhao, Z.; Liu, S.; Li, Y.; Xiao, M.; Chen, J.L. Identification of lncRNA-155 encoded by MIR155HG as a novel regulator of innate immunity against influenza A virus infection. Cell. Microbiol. 2019, 21, e13036. [Google Scholar] [CrossRef]
- Pan, Q.; Zhao, Z.; Liao, Y.; Chiu, S.; Wang, S.; Chen, B.; Chen, N.; Chen, Y.; Chen, J. Identification of an Interferon-Stimulated Long Noncoding RNA (LncRNA ISR) Involved in Regulation of Influenza A Virus Replication. Int. J. Mol. Sci. 2019, 20, 5118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Jiang, M.; Liu, L.; Yang, Z.; Ma, Z.; Liu, S.; Ma, Y.; Zhang, L.; Cao, X. The long noncoding RNA Lnczc3h7a promotes a TRIM25-mediated RIG-I antiviral innate immune response. Nat. Immunol. 2019, 20, 812–823. [Google Scholar] [CrossRef]
- Lai, C.; Liu, L.; Liu, Q.; Wang, K.; Cheng, S.; Zhao, L.; Xia, M.; Wang, C.; Duan, Y.; Zhang, L.; et al. Long noncoding RNA AVAN promotes antiviral innate immunity by interacting with TRIM25 and enhancing the transcription of FOXO3a. Cell Death Differ. 2021, 28, 2900–2915. [Google Scholar] [CrossRef]
- Zhao, L.; Xia, M.; Wang, K.; Lai, C.; Fan, H.; Gu, H.; Yang, P.; Wang, X. A Long Non-coding RNA IVRPIE Promotes Host Antiviral Immune Responses Through Regulating Interferon β1 and ISG Expression. Front. Microbiol. 2020, 11, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Hu, J.; Liu, S.; Chen, B.; Xiao, M.; Li, Y.; Liao, Y.; Rai, K.R.; Zhao, Z.; Ouyang, J.; et al. RDUR, a lncRNA, Promotes Innate Antiviral Responses and Provides Feedback Control of NF-κB Activation. Front. Immunol. 2021, 12, 672165. [Google Scholar] [CrossRef] [PubMed]
- Barriocanal, M.; Prior, C.; Suarez, B.; Unfried, J.P.; Razquin, N.; Hervás-Stubbs, S.; Sangro, B.; Segura, V.; Fortes, P. Long Noncoding RNA EGOT Responds to Stress Signals to Regulate Cell Inflammation and Growth. J. Immunol. 2021, 206, 1932–1942. [Google Scholar] [CrossRef]
- Carnero, E.; Barriocanal, M.; Prior, C.; Pablo Unfried, J.; Segura, V.; Guruceaga, E.; Enguita, M.; Smerdou, C.; Gastaminza, P.; Fortes, P. Long noncoding RNA EGOT negatively affects the antiviral response and favors HCV replication. EMBO Rep. 2016, 17, 1013–1028. [Google Scholar] [CrossRef]
- Liu, S.; Liu, J.; Yang, X.; Jiang, M.; Wang, Q.; Zhang, L.; Ma, Y.; Shen, Z.; Tian, Z.; Cao, X. Cis-acting lnc-Cxcl2 restrains neutrophil-mediated lung inflammation by inhibiting epithelial cell CXCL2 expression in virus infection. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Abak, A.; Shoorei, H.; Talebi, S.F.; Mohaqiq, M.; Sarabi, P.; Taheri, M.; Mokhtari, M. Interaction between non-coding RNAs and Toll-like receptors. Biomed. Pharmacother. 2021, 140, 111784. [Google Scholar] [CrossRef] [PubMed]
- More, S.; Zhu, Z.; Lin, K.; Huang, C.; Pushparaj, S.; Liang, Y.; Sathiaseelan, R.; Yang, X.; Liu, L. Long non-coding RNA PSMB8-AS1 regulates influenza virus replication. RNA Biol. 2019, 16, 340–353. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Guo, G.; Lu, M.; Chai, W.; Li, Y.; Tong, X.; Li, J.; Jia, X.; Liu, W.; Qi, D.; et al. Long Noncoding RNA Lnc-MxA Inhibits Beta Interferon Transcription by Forming RNA-DNA Triplexes at Its Promoter. J. Virol. 2019, 93, e00786-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almatrrouk, S.; Saba, I.; Abozaid, S.; Al-Qahtani, A.A.; Al-Ahdal, M.N. Virus sensing receptors in cellular infectivity of influenza A virus. J. Infect. Dev. Countr. 2021, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, M.; Wang, X.; Sun, M.; Ma, C.; Liang, W.; Gao, X.; Wei, L. Long Noncoding RNA NRAV Promotes Respiratory Syncytial Virus Replication by Targeting the MicroRNA miR-509-3p/Rab5c Axis to Regulate Vesicle Transportation. J. Virol. 2020, 94, e00113-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winterling, C.; Koch, M.; Koeppel, M.; Garcia-Alcalde, F.; Karlas, A.; Meyer, T.F. Evidence for a crucial role of a host non-coding RNA in influenza A virus replication. RNA Biol. 2014, 11, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, S.; Yang, Z.; Lin, H.; Zhu, J.; Liu, L.; Wang, W.; Liu, S.; Liu, W.; Ma, Y.; et al. Self-Recognition of an Inducible Host lncRNA by RIG-I Feedback Restricts Innate Immune Response. Cell 2018, 173, 906–919. [Google Scholar] [CrossRef] [Green Version]
- Runtsch, M.C.; O’Neill, L.A. GOTcha: LncRNA-ACOD1 targets metabolism during viral infection. Cell Res. 2018, 28, 137–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Xu, J.; Wang, Y.; Cao, X. An interferon-independent lncRNA promotes viral replication by modulating cellular metabolism. Science (Am. Assoc. Adv. Sci.) 2017, 358, 1051–1055. [Google Scholar] [CrossRef] [Green Version]
- Amort, M.; Nachbauer, B.; Tuzlak, S.; Kieser, A.; Schepers, A.; Villunger, A.; Polacek, N. Expression of the vault RNA protects cells from undergoing apoptosis. Nat. Commun. 2015, 6, 7030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Chen, Y.; Zhang, Z.; Ouyang, J.; Wang, Y.; Yan, R.; Huang, S.; Gao, G.F.; Guo, G.; Chen, J. Robust expression of vault RNAs induced by influenza A virus plays a critical role in suppression of PKR-mediated innate immunity. Nucleic Acids Res. 2015, 43, 10321–10337. [Google Scholar] [CrossRef]
- Ge, J.; Wang, J.; Xiong, F.; Jiang, X.; Zhu, K.; Wang, Y.; Mo, Y.; Gong, Z.; Zhang, S.; He, Y.; et al. Epstein-Barr virus-encoded circular RNA circBART2.2 promotes immune escape of nasopharyngeal carcinoma by regulating PD-L1. Cancer Res. 2021, 81, 5074–5088. [Google Scholar] [CrossRef]
- Hu, Y.; Xu, Y.; Deng, X.; Wang, R.; Li, R.; You, L.; Song, J.; Zhang, Y. Comprehensive analysis of the circRNA expression profile and circRNA-miRNA-mRNA network in the pathogenesis of EV-A71 infection. Virus Res. 2021, 303, 198502. [Google Scholar] [CrossRef]
- Zhao, W.; Su, J.; Wang, N.; Zhao, N.; Su, S. Expression Profiling and Bioinformatics Analysis of CircRNA in Mice Brain Infected with Rabies Virus. Int. J. Mol. Sci. 2021, 22, 6537. [Google Scholar] [CrossRef]
- Choudhary, A.; Madbhagat, P.; Sreepadmanabh, M.; Bhardwaj, V.; Chande, A. Circular RNA as an Additional Player in the Conflicts between the Host and the Virus. Front. Immunol. 2021, 12, 602006. [Google Scholar] [CrossRef]
- Qu, Z.; Meng, F.; Shi, J.; Deng, G.; Zeng, X.; Ge, J.; Li, Y.; Liu, L.; Chen, P.; Jiang, Y.; et al. A Novel Intronic Circular RNA Antagonizes Influenza Virus by Absorbing a microRNA That Degrades CREBBP and Accelerating IFN-β Production. MBio 2021, e01017-21. [Google Scholar] [CrossRef]
- Yu, T.; Ding, Y.; Zhang, Y.; Liu, Y.; Li, Y.; Lei, J.; Zhou, J.; Song, S.; Hu, B. Circular RNA GATAD2A promotes H1N1 replication through inhibiting autophagy. Vet. Microbiol. 2019, 231, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.; Zhu, X.; Chen, Y.; Wei, H.; Chen, Q.; Chi, X.; Qi, B.; Zhang, L.; Zhao, Y.; Gao, G.F.; et al. NRAV, a Long Noncoding RNA, Modulates Antiviral Responses through Suppression of Interferon-Stimulated Gene Transcription. Cell Host Microbe 2014, 16, 616–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imamura, K.; Imamachi, N.; Akizuki, G.; Kumakura, M.; Kawaguchi, A.; Nagata, K.; Kato, A.; Kawaguchi, Y.; Sato, H.; Yoneda, M.; et al. Long noncoding RNA NEAT1-dependent SFPQ relocation from promoter region to paraspeckle mediates IL8 expression upon immune stimuli. Mol. Cell 2014, 53, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Chai, W.; Li, J.; Shangguan, Q.; Liu, Q.; Li, X.; Qi, D.; Tong, X.; Liu, W.; Ye, X.; Gallagher, T. Lnc-ISG20 Inhibits Influenza A Virus Replication by Enhancing ISG20 Expression. J. Virol. 2018, 92, e00539-18. [Google Scholar] [CrossRef] [Green Version]
- Xiao, M.; Chen, Y.; Wang, S.; Liu, S.; Rai, K.R.; Chen, B.; Li, F.; Li, Y.; Maarouf, M.; Chen, J. LncRNA IFITM4P regulates host antiviral responses by acting as a ceRNA. J. Virol. 2021, I27721. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, D.; Feng, W.; Guo, Y.; Sun, X.; Zhang, M.; Guan, Z.; Duan, M. Long noncoding RNA TSPOAP1 antisense RNA 1 negatively modulates type I IFN signaling to facilitate influenza A virus replication. J. Med. Virol. 2019, 94, 557–566. [Google Scholar] [CrossRef]
- Tu, Y.H.; Guo, Y.; Ji, S.; Shen, J.L.; Fei, G.H. The Influenza A Virus H3N2 Triggers the Hypersusceptibility of Airway Inflammatory Response via Activating the lncRNA TUG1/miR-145-5p/NF-kappaB Pathway in COPD. Front. Pharmacol. 2021, 12, 604590. [Google Scholar] [CrossRef]
- Shi, N.; Zhang, S.; Guo, Y.; Yu, X.; Zhao, W.; Zhang, M.; Guan, Z.; Duan, M. CircRNA_0050463 promotes influenza A virus replication by sponging miR-33b-5p to regulate EEF1A1. Vet. Microbiol. 2021, 254, 108995. [Google Scholar] [CrossRef] [PubMed]
- Khongnomnan, K.; Saengchoowong, S.; Mayuramart, O.; Nimsamer, P.; Pisitkun, T.; Poovorawan, Y.; Payungporn, S. Hsa-miR-30e-3p inhibits influenza B virus replication by targeting viral NA and NP genes. Exp. Biol. Med. 2020, 245, 1664–1671. [Google Scholar] [CrossRef]
- Liu, H.; Yang, X.; Zhang, Z.; Zou, W.; Wang, H. miR-146a-5p promotes replication of infectious bronchitis virus by targeting IRAK2 and TNFRSF18. Microb. Pathog. 2018, 120, 32–36. [Google Scholar] [CrossRef]
- Li, H.; Li, J.; Zhai, Y.; Zhang, L.; Cui, P.; Feng, L.; Fu, X.; Tian, Y.; Wang, H.; Yang, X.; et al. Gga-miR-30d regulates infectious bronchitis virus infection by targeting USP47 in HD11 cells. Microb. Pathog. 2020, 141, 103998. [Google Scholar] [CrossRef]
- Li, H.; Cui, P.; Fu, X.; Zhang, L.; Yan, W.; Zhai, Y.; Lei, C.; Wang, H.; Yang, X. Identification and analysis of long non-coding RNAs and mRNAs in chicken macrophages infected with avian infectious bronchitis coronavirus. BMC Genom. 2021, 22, 67. [Google Scholar] [CrossRef]
- Carrique, L.; Fan, H.; Walker, A.P.; Keown, J.R.; Sharps, J.; Staller, E.; Barclay, W.S.; Fodor, E.; Grimes, J.M. Host ANP32A mediates the assembly of the influenza virus replicase. Nature 2020, 587, 638–643. [Google Scholar] [CrossRef]
- Sato, K.; Hayashi, H.; Shimotai, Y.; Yamaya, M.; Hongo, S.; Kawakami, K.; Matsuzaki, Y.; Nishimura, H. TMPRSS2 activates hemagglutinin-esterase glycoprotein of influenza C virus. J. Virol. 2021, 95, I129621. [Google Scholar] [CrossRef]
- Saengchoowong, S.; Khongnomnan, K.; Poomipak, W.; Praianantathavorn, K.; Poovorawan, Y.; Zhang, Q.; Payungporn, S. High-Throughput MicroRNA Profiles of Permissive Madin-Darby Canine Kidney Cell Line Infected with Influenza B Viruses. Viruses 2019, 11, 986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, H.; Zhang, X.; Liu, B.; Jia, G.; Li, W. Circular RNA circ-ABCB10 promotes breast cancer proliferation and progression through sponging miR-1271. Am. J. Cancer Res. 2017, 7, 1566–1576. [Google Scholar] [PubMed]
- Qiu, F.; Chen, H.; Ying, H.; Yu, W.; Guo, M.; Zhou, W.; Yu, C. A Biological Insight into the Susceptibility to Influenza Infection in Junior Rats by Comprehensive Analysis of lncRNA Profiles. Biomed Res. Int. 2021, 2021, 8112783. [Google Scholar] [CrossRef]
- Fukuyama, H.; Shinnakasu, R.; Kurosaki, T. Influenza vaccination strategies targeting the hemagglutinin stem region. Immunol. Rev. 2020, 296, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Mohan, T.; Chang, T.Z.; Gonzalez, G.X.; Wang, Y.; Kwon, Y.M.; Kang, S.-M.; Compans, R.W.; Champion, J.A.; Wang, B.-Z. Influenza NP core and HA or M2e shell double-layered protein nanoparticles induce broad protection against divergent influenza a viruses. Nanomed. Nanotechnol. Biol. Med. 2021, 9, 102479. [Google Scholar] [CrossRef]
- Chen, J.; Yuan, L.; Fan, Q.; Su, F.; Chen, Y.; Hu, S. Adjuvant effect of docetaxel on the immune responses to influenza A H1N1 vaccine in mice. BMC Immunol. 2012, 13, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, S.; Wang, J.; Wei, S.; Li, C.; Zhou, K.; Hu, J.; Ye, X.; Yan, J.; Liu, W.; Gao, G.F.; et al. Endogenous Cellular MicroRNAs Mediate Antiviral Defense against Influenza A Virus. Mol. Ther. Nucleic Acids 2018, 10, 361–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Yang, T.; Liu, X.; Xiong, F.; Luo, J.; Yi, Y.; Fan, J.; Chen, Z.; Tan, W.-S. MiRNA Targeted NP Genome of Live Attenuated Influenza Vaccines Provide Cross-Protection against a Lethal Influenza Virus Infection. Vaccines 2020, 8, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Y.; Guo, S.; Liu, G.; Qiu, Z.; Wang, J.; Yang, D.; Tian, X.; Qiao, Z.; Ma, Z.; Liu, Z. Host Non-Coding RNA Regulates Influenza A Virus Replication. Viruses 2022, 14, 51. https://doi.org/10.3390/v14010051
Liao Y, Guo S, Liu G, Qiu Z, Wang J, Yang D, Tian X, Qiao Z, Ma Z, Liu Z. Host Non-Coding RNA Regulates Influenza A Virus Replication. Viruses. 2022; 14(1):51. https://doi.org/10.3390/v14010051
Chicago/Turabian StyleLiao, Yuejiao, Shouqing Guo, Geng Liu, Zhenyu Qiu, Jiamin Wang, Di Yang, Xiaojing Tian, Ziling Qiao, Zhongren Ma, and Zhenbin Liu. 2022. "Host Non-Coding RNA Regulates Influenza A Virus Replication" Viruses 14, no. 1: 51. https://doi.org/10.3390/v14010051
APA StyleLiao, Y., Guo, S., Liu, G., Qiu, Z., Wang, J., Yang, D., Tian, X., Qiao, Z., Ma, Z., & Liu, Z. (2022). Host Non-Coding RNA Regulates Influenza A Virus Replication. Viruses, 14(1), 51. https://doi.org/10.3390/v14010051