


An Optimized FI-RSV Vaccine Effectively Protects Cotton Rats and BALB/c Mice without Causing Enhanced Respiratory Disease

 , , and
, , and 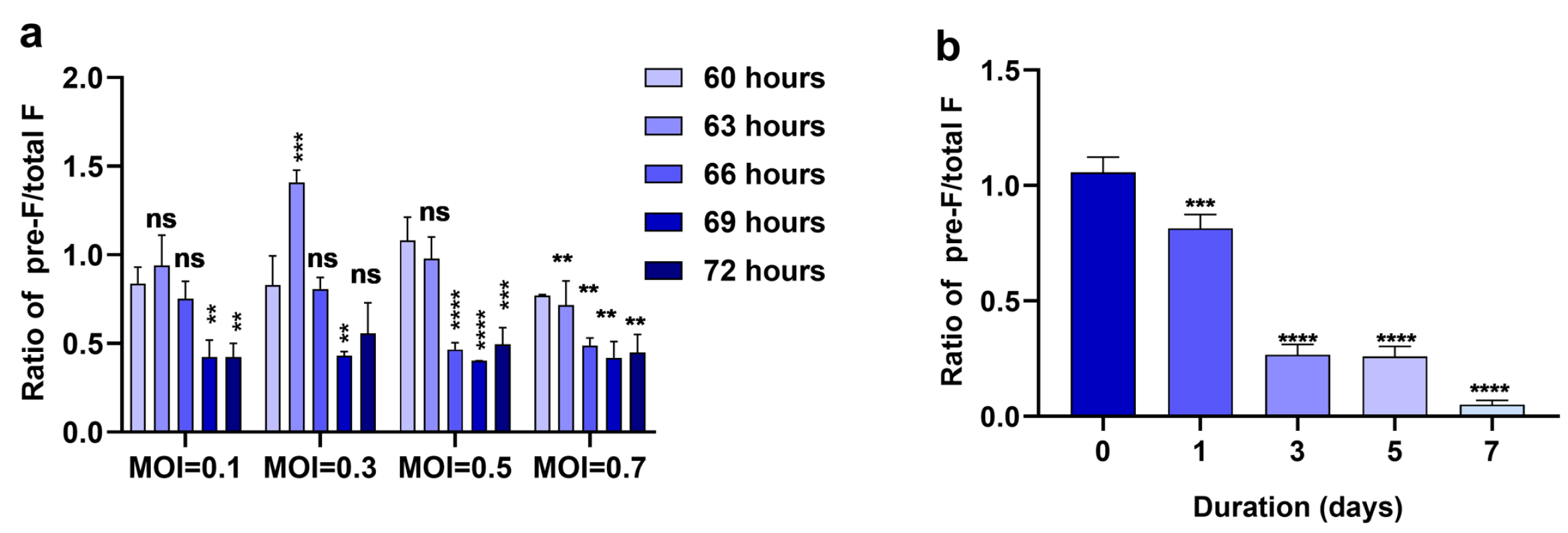{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Virus and Animals
2.2. Antibodies and Proteins
2.3. Vaccine Preparation
2.4. Thermal Stability
2.5. Biotin–Streptavidin-Based Sandwich ELISA
2.6. Animal Vaccination and RSV Challenge
2.7. IgG Subclass and Avidity Immunoassays
2.8. RSV F Competitive ELISA
2.9. Neutralization Assay
2.10. Plaque Assay
2.11. Statistical Analysis
3. Results
3.1. Pre-F on RSV A2 Virions Is Rapidly Lost within 3 Days
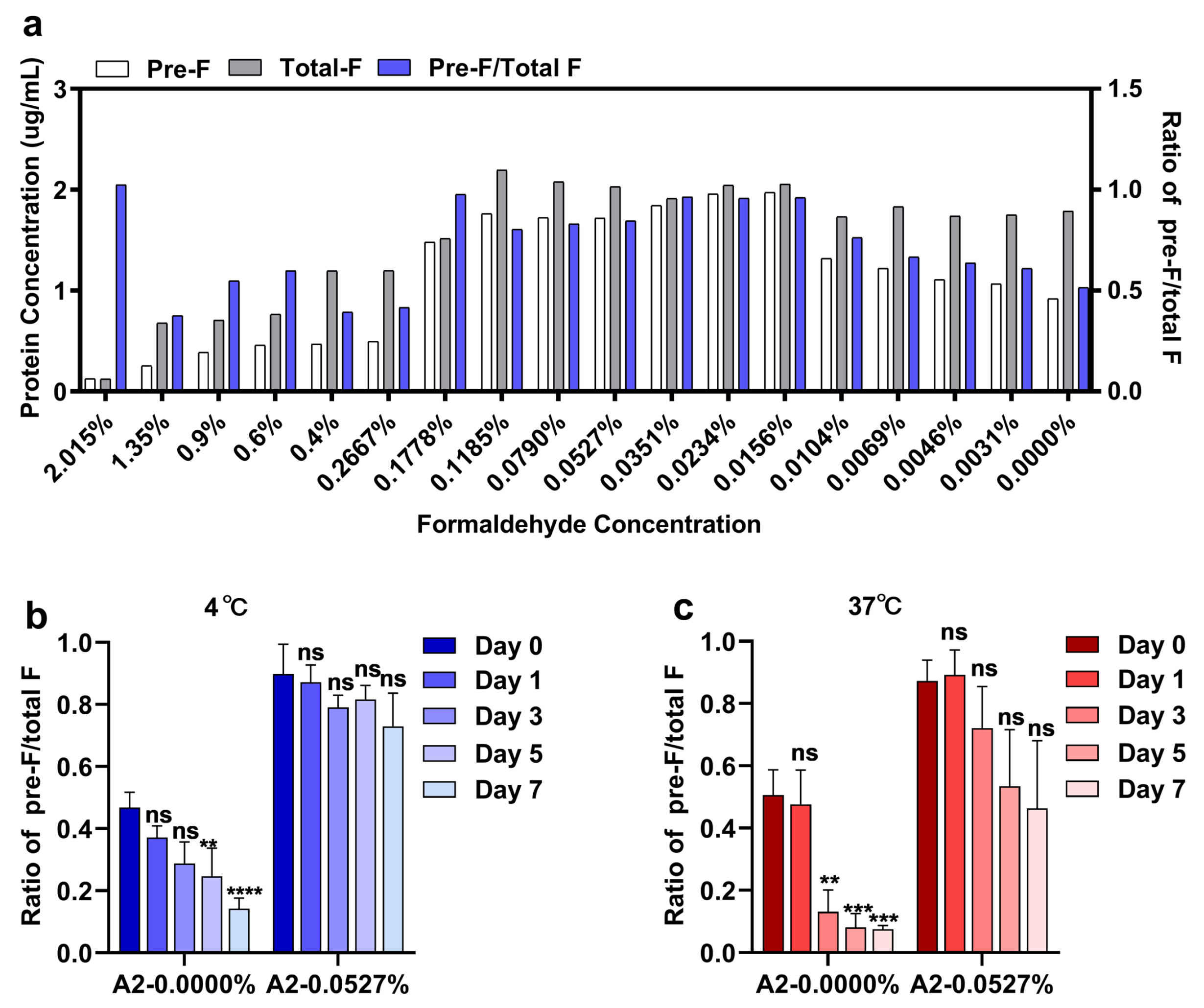
3.2. Pre-F-Specific Epitopes Are Preserved Well on RSV A2 Virions by an Optimized Formaldehyde Concentration with Improved Thermostability
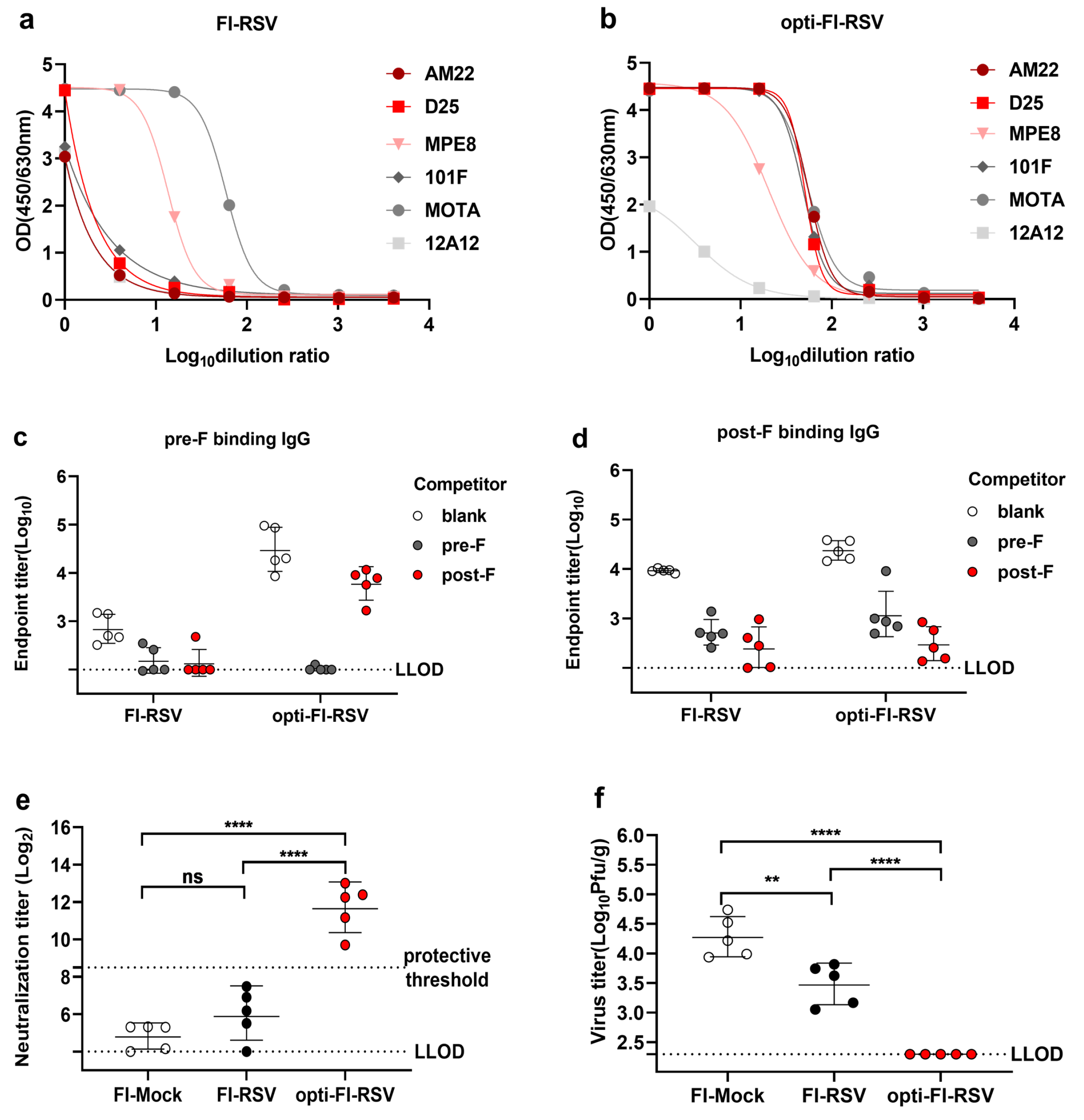
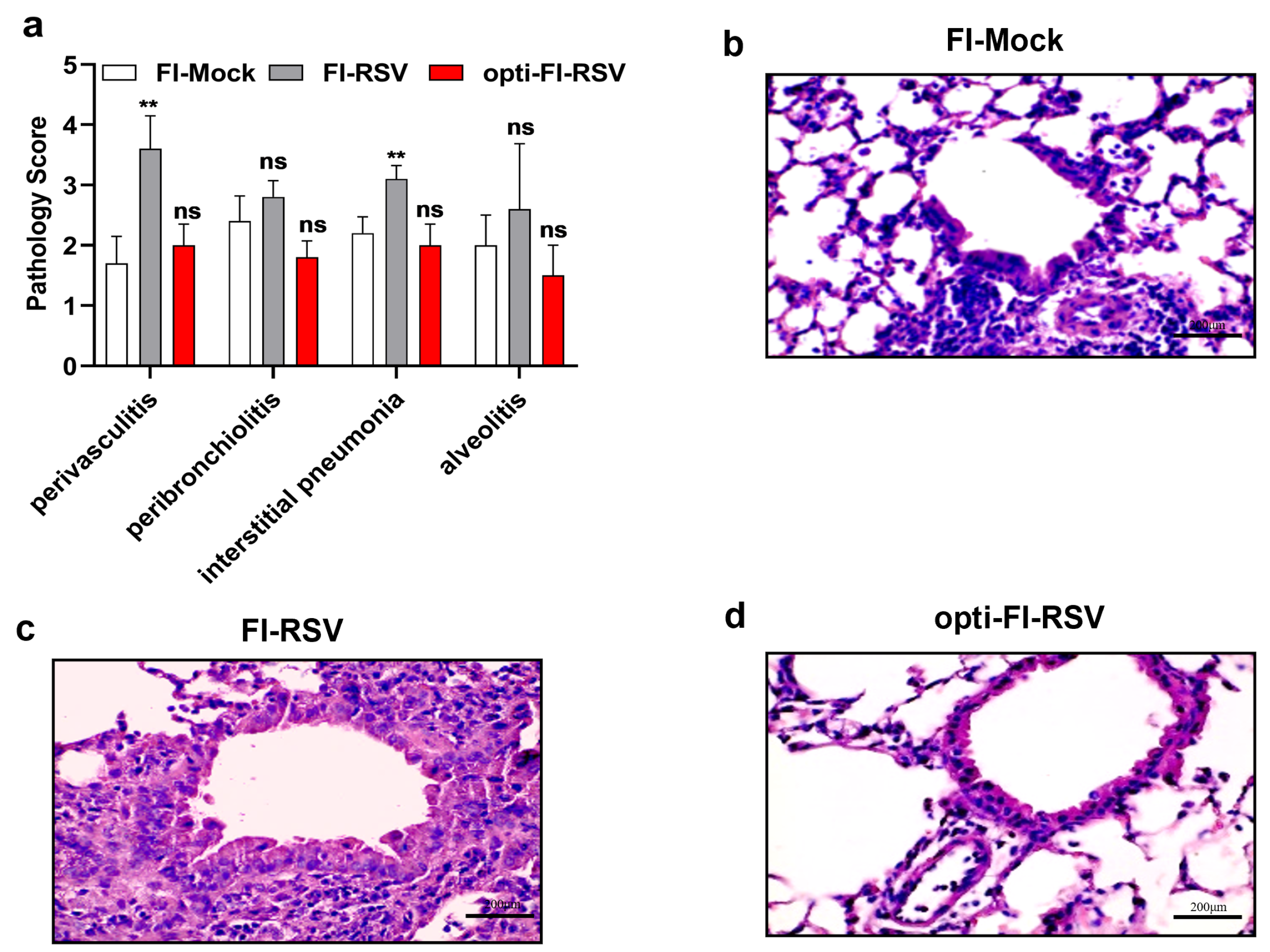
3.3. Opti-FI-RSV Exerts Immunogenic and Protective Effects without Causing ERD in Cotton Rats
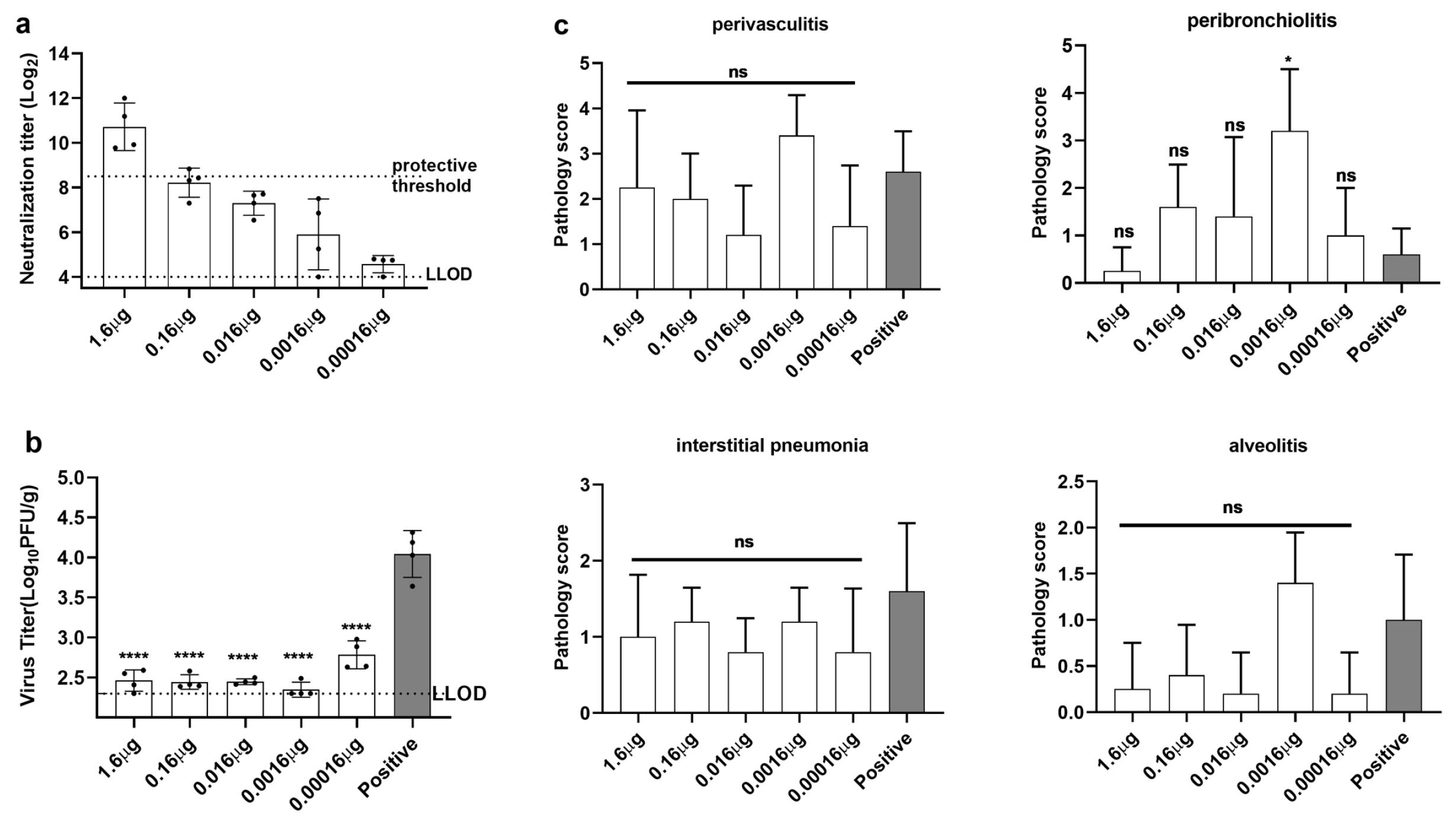
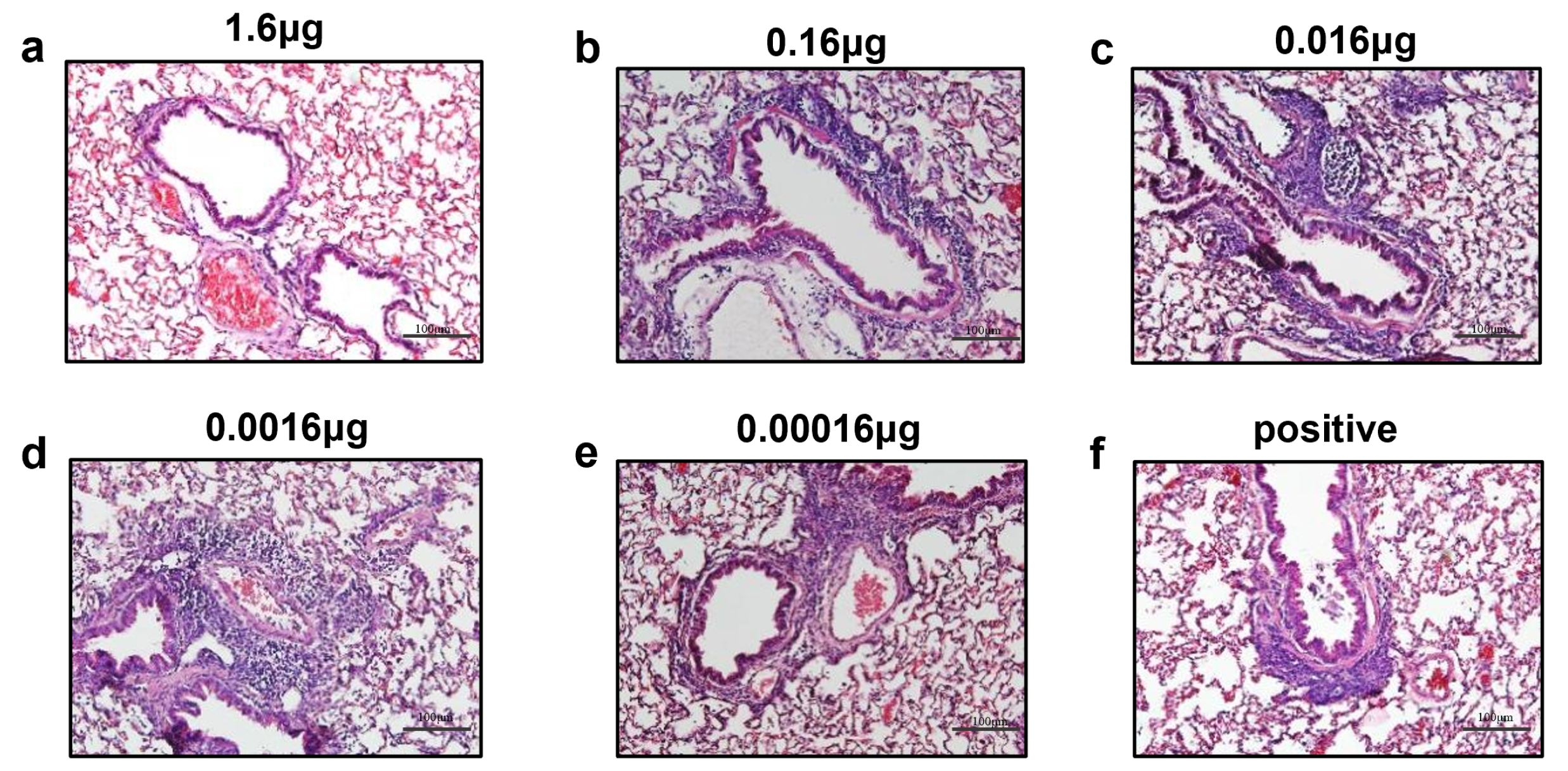
3.4. Immunization with opti-FI-RSV at Suboptimal Doses Primes ERD in Cotton Rats
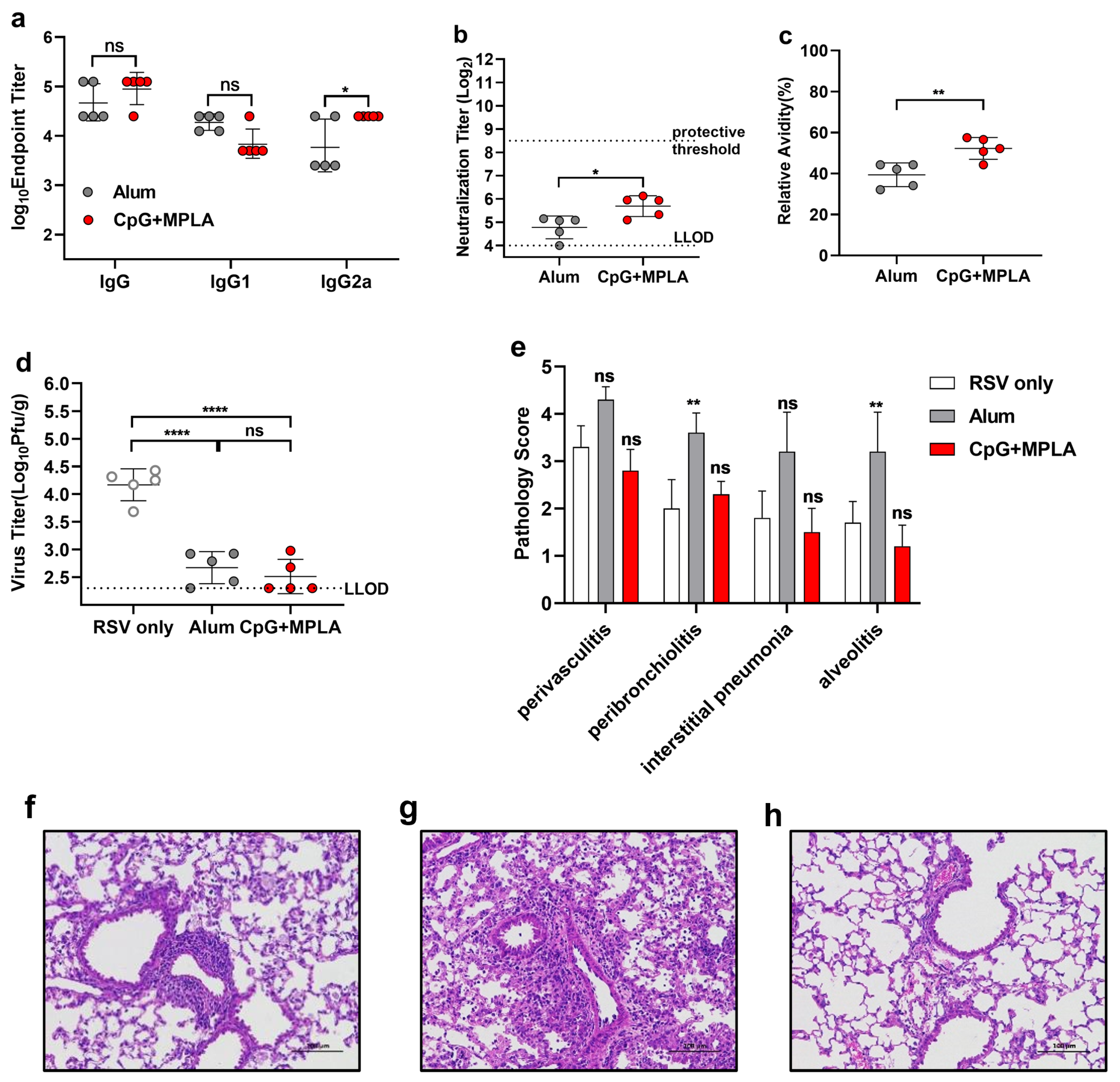
3.5. A Combination Adjuvant Modulates Immune Responses to Prevent ERD, Possibly by Promoting Antibody Affinity Maturation
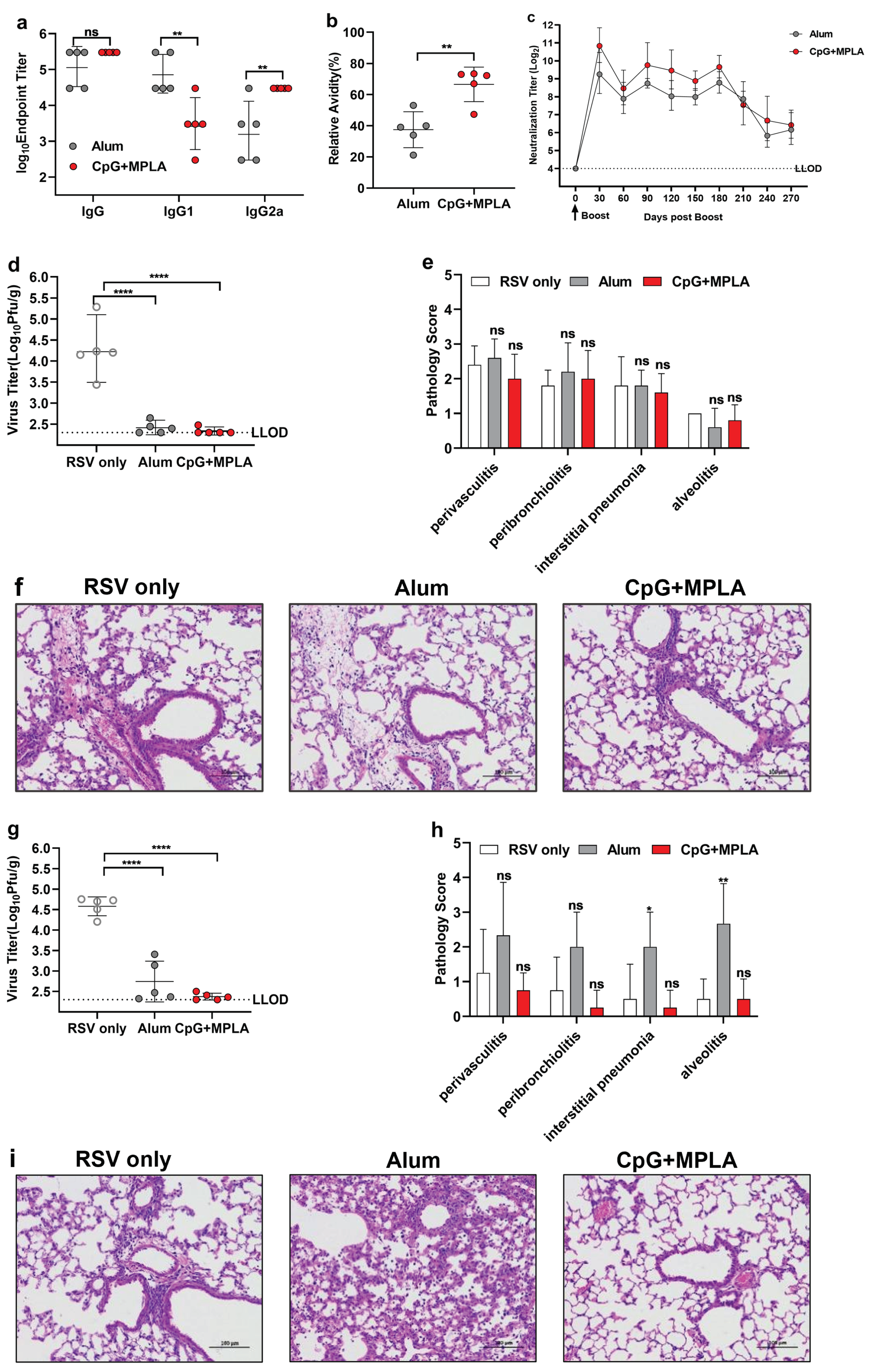
3.6. Opti-FI-RSV plus CpG + MPLA Adjuvant Still Provides Protection after the Neutralizing Antibody Levels Decline and Prevents ERD
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| RSV | respiratory syncytial virus |
| pre-F | prefusion protein |
| post-F | postfusion protein |
| ERD | enhanced respiratory disease |
| ALRIs | acute lower respiratory infections |
| FI-RSV | formaldehyde-inactivated RSV |
| DMEM | Dulbecco’s modified Eagle’s medium |
| PFU | plaque-forming units |
| HRP | horse radish peroxidase |
| TMB | tetramethylbenzidine |
| MOI | multiplicities of infection |
| H & E | hematoxylin and eosin |
| TLR-4 | Toll-like receptor 4 |
| TLR-9 | Toll-like receptor 9 |
| mAb | monoclonal antibody |
References
- McAllister, D.A.; Liu, L.; Shi, T.; Chu, Y.; Reed, C.; Burrows, J.; Adeloye, D.; Rudan, I.; Black, R.E.; Campbell, H.; et al. Global, regional, and national estimates of pneumonia morbidity and mortality in children younger than 5 years between 2000 and 2015: A systematic analysis. Lancet. Glob. Health 2019, 7, e47–e57. [Google Scholar] [CrossRef]
- Bénet, T.; Sánchez Picot, V.; Messaoudi, M.; Chou, M.; Eap, T.; Wang, J.; Shen, K.; Pape, J.W.; Rouzier, V.; Awasthi, S.; et al. Microorganisms Associated With Pneumonia in Children <5 Years of Age in Developing and Emerging Countries: The GABRIEL Pneumonia Multicenter, Prospective, Case-Control Study. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2017, 65, 604–612. [Google Scholar] [CrossRef]
- O’Brien, K.L.; Baggett, H.C.; Brooks, W.A.; Feikin, D.R.; Hammitt, L.L.; Higdon, M.M.; Howie, S.R.; Deloria Knoll, M.; Kotloff, K.L.; Levine, O.S.; et al. Causes of severe pneumonia requiring hospital admission in children without HIV infection from Africa and Asia: The PERCH multi-country case-control study. Lancet 2019, 394, 757–779. [Google Scholar] [CrossRef]
- Glezen, W.P.; Taber, L.H.; Frank, A.L.; Kasel, J.A. Risk of primary infection and reinfection with respiratory syncytial virus. Am. J. Dis. Child. (1960) 1986, 140, 543–546. [Google Scholar] [CrossRef]
- Hall, C.B.; Weinberg, G.A.; Iwane, M.K.; Blumkin, A.K.; Edwards, K.M.; Staat, M.A.; Auinger, P.; Griffin, M.R.; Poehling, K.A.; Erdman, D.; et al. The Burden of Respiratory Syncytial Virus Infection in Young Children. N. Engl. J. Med. 2009, 360, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Thompson, W.W.; Viboud, C.G.; Ringholz, C.M.; Cheng, P.Y.; Steiner, C.; Abedi, G.R.; Anderson, L.J.; Brammer, L.; Shay, D.K. Hospitalizations associated with influenza and respiratory syncytial virus in the United States, 1993–2008. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2012, 54, 1427–1436. [Google Scholar] [CrossRef]
- Mejias, A.; Rodríguez-Fernández, R.; Oliva, S.; Peeples, M.E.; Ramilo, O. The Journey to an RSV Vaccine. Ann. Allergy Asthma Immunol. Off. Publ. Am. Coll. Allergy Asthma, Immunol. 2020, 125, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Fulginiti, V.A.; Eller, J.J.; Sieber, O.F.; Joyner, J.W.; Minamitani, M.; Meiklejohn, G. Respiratory virus immunization. I. A field trial of two inactivated respiratory virus vaccines; an aqueous trivalent parainfluenza virus vaccine and an alum-precipitated respiratory syncytial virus vaccine. Am. J. Epidemiol. 1969, 89, 435–448. [Google Scholar] [CrossRef]
- Murphy, B.R.; Prince, G.A.; Walsh, E.E.; Kim, H.W.; Parrott, R.H.; Hemming, V.G.; Rodriguez, W.J.; Chanock, R.M. Dissociation between serum neutralizing and glycoprotein antibody responses of infants and children who received inactivated respiratory syncytial virus vaccine. J. Clin. Microbiol. 1986, 24, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Boelen, A.; Andeweg, A.; Kwakkel, J.; Lokhorst, W.; Bestebroer, T.; Dormans, J.; Kimman, T. Both immunisation with a formalin-inactivated respiratory syncytial virus (RSV) vaccine and a mock antigen vaccine induce severe lung pathology and a Th2 cytokine profile in RSV-challenged mice. Vaccine 2000, 19, 982–991. [Google Scholar] [CrossRef]
- McLellan, J.S.; Chen, M.; Joyce, M.G.; Sastry, M.; Stewart-Jones, G.B.E.; Yang, Y.; Zhang, B.; Chen, L.; Srivatsan, S.; Zheng, A.; et al. Structure-Based Design of a Fusion Glycoprotein Vaccine for Respiratory Syncytial Virus. Science 2013, 342, 592–598. [Google Scholar] [CrossRef]
- Krarup, A.; Truan, D.; Furmanova-Hollenstein, P.; Bogaert, L.; Bouchier, P.; Bisschop, I.J.M.; Widjojoatmodjo, M.N.; Zahn, R.; Schuitemaker, H.; McLellan, J.S.; et al. A highly stable prefusion RSV F vaccine derived from structural analysis of the fusion mechanism. Nat. Commun. 2015, 6, 8143. [Google Scholar] [CrossRef]
- Aliprantis, A.O.; Shaw, C.A.; Griffin, P.; Farinola, N.; Railkar, R.A.; Cao, X.; Liu, W.; Sachs, J.R.; Swenson, C.J.; Lee, H.; et al. A phase 1, randomized, placebo-controlled study to evaluate the safety and immunogenicity of an mRNA-based RSV prefusion F protein vaccine in healthy younger and older adults. Hum. Vaccines Immunother. 2021, 17, 1248–1261. [Google Scholar] [CrossRef]
- Espeseth, A.S.; Cejas, P.J.; Citron, M.P.; Wang, D.; DiStefano, D.J.; Callahan, C.; Donnell, G.O.; Galli, J.D.; Swoyer, R.; Touch, S.; et al. Modified mRNA/lipid nanoparticle-based vaccines expressing respiratory syncytial virus F protein variants are immunogenic and protective in rodent models of RSV infection. Npj Vaccines 2020, 5, 16. [Google Scholar] [CrossRef]
- Cullen, L.M.; Blanco, J.C.G.; Morrison, T.G. Cotton rat immune responses to virus-like particles containing the pre-fusion form of respiratory syncytial virus fusion protein. J. Transl. Med. 2015, 13, 350. [Google Scholar] [CrossRef]
- Cimica, V.; Boigard, H.; Bhatia, B.; Fallon, J.T.; Alimova, A.; Gottlieb, P.; Galarza, J.M. Novel Respiratory Syncytial Virus-Like Particle Vaccine Composed of the Postfusion and Prefusion Conformations of the F Glycoprotein. Clin. Vaccine Immunol. CVI 2016, 23, 451–459. [Google Scholar] [CrossRef]
- Williams, K.; Bastian, A.R.; Feldman, R.A.; Omoruyi, E.; de Paepe, E.; Hendriks, J.; van Zeeburg, H.; Godeaux, O.; Langedijk, J.P.M.; Schuitemaker, H.; et al. Phase 1 Safety and Immunogenicity Study of a Respiratory Syncytial Virus Vaccine With an Adenovirus 26 Vector Encoding Prefusion F (Ad26.RSV.preF) in Adults Aged ≥60 Years. J. Infect. Dis. 2020, 222, 979–988. [Google Scholar] [CrossRef]
- Killikelly, A.M.; Kanekiyo, M.; Graham, B.S. Pre-fusion F is absent on the surface of formalin-inactivated respiratory syncytial virus. Sci. Rep. 2016, 6, 34108. [Google Scholar] [CrossRef]
- Greco, D.; Salmaso, S.; Mastrantonio, P.; Giuliano, M.; Tozzi, A.E.; Anemona, A.; Ciofi degli Atti, M.L.; Giammanco, A.; Panei, P.; Blackwelder, W.C.; et al. A Controlled Trial of Two Acellular Vaccines and One Whole-Cell Vaccine against Pertussis. New Engl. J. Med. 1996, 334, 341–349. [Google Scholar] [CrossRef]
- Werzberger, A.; Mensch, B.; Kuter, B.; Brown, L.; Lewis, J.; Sitrin, R.; Miller, W.; Shouval, D.; Wiens, B.; Calandra, G.; et al. A controlled trial of a formalin-inactivated hepatitis A vaccine in healthy children. New Engl. J. Med. 1992, 327, 453–457. [Google Scholar] [CrossRef]
- Murdin, A.D.; Barreto, L.; Plotkin, S. Inactivated poliovirus vaccine: Past and present experience. Vaccine 1996, 14, 735–746. [Google Scholar] [CrossRef]
- Wester, K.; Wahlund, E.; Sundström, C.; Ranefall, P.; Bengtsson, E.; Russell, P.J.; Ow, K.T.; Malmström, P.U.; Busch, C. Paraffin section storage and immunohistochemistry. Effects of time, temperature, fixation, and retrieval protocol with emphasis on p53 protein and MIB1 antigen. Appl. Immunohistochem. Mol. Morphol. AIMM 2000, 8, 61–70. [Google Scholar] [CrossRef]
- van den Broek, L.J.; van de Vijver, M.J. Assessment of problems in diagnostic and research immunohistochemistry associated with epitope instability in stored paraffin sections. Appl. Immunohistochem. Mol. Morphol. AIMM 2000, 8, 316–321. [Google Scholar] [PubMed]
- Zhang, W.; Zhang, L.J.; Zhan, L.T.; Zhao, M.; Wu, G.H.; Si, J.Y.; Chen, L.; Lin, X.; Sun, Y.P.; Lin, M.; et al. The Optimal Concentration of Formaldehyde is Key to Stabilizing the Pre-Fusion Conformation of Respiratory Syncytial Virus Fusion Protein. Viruses 2019, 11, 628. [Google Scholar] [CrossRef] [PubMed]
- McLellan, J.S.; Chen, M.; Leung, S.; Graepel, K.W.; Du, X.; Yang, Y.; Zhou, T.; Baxa, U.; Yasuda, E.; Beaumont, T.; et al. Structure of RSV Fusion Glycoprotein Trimer Bound to a Prefusion-Specific Neutralizing Antibody. Science (N. Y.) 2013, 340, 1113–1117. [Google Scholar] [CrossRef]
- Chaiwatpongsakorn, S.; Epand, R.F.; Collins, P.L.; Epand, R.M.; Peeples, M.E. Soluble Respiratory Syncytial Virus Fusion Protein in the Fully Cleaved, Pretriggered State Is Triggered by Exposure to Low-Molarity Buffer. J. Virol. 2011, 85, 3968–3977. [Google Scholar] [CrossRef]
- Kim, H.W.; Canchola, J.G.; Brandt, C.D.; Pyles, G.; Chanock, R.M.; Jensen, K.; Parrott, R.H. Respiratory syncytial virus disease in infants despite prior administration of antigenic inactivated vaccine. Am. J. Epidemiol. 1969, 89, 422–434. [Google Scholar] [CrossRef]
- Fertey, J.; Thoma, M.; Beckmann, J.; Bayer, L.; Finkensieper, J.; Reißhauer, S.; Berneck, B.S.; Issmail, L.; Schönfelder, J.; Casado, J.P.; et al. Automated application of low energy electron irradiation enables inactivation of pathogen-and cell-containing liquids in biomedical research and production facilities. Sci. Rep. 2020, 10, 1–14. [Google Scholar]
- Varnamkhasti, F.A.; Kia, V.; Shokri, R.; Moghaddam, M.J.M.; Paryan, M. Design and development of a simple method for the detection and quantification of residual host cell DNA in recombinant rotavirus vaccine. Mol. Cell. Probes 2021, 55, 101674. [Google Scholar] [CrossRef]
- Blanco, J.C.; Pletneva, L.M.; Otoa, R.O.; Patel, M.C.; Vogel, S.N.; Boukhvalova, M.S. Preclinical assessment of safety of maternal vaccination against respiratory syncytial virus (RSV) in cotton rats. Vaccine 2017, 35, 3951–3958. [Google Scholar] [CrossRef]
- Delgado, M.F.; Coviello, S.; Monsalvo, A.C.; Melendi, G.A.; Hernandez, J.Z.; Batalle, J.P.; Diaz, L.; Trento, A.; Chang, H.Y.; Mitzner, W.; et al. Lack of antibody affinity maturation due to poor Toll stimulation led to enhanced RSV disease. Nat. Med. 2009, 15, 34–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polack, F.P.; Hoffman, S.J.; Crujeiras, G.; Griffin, D.E. A role for nonprotective complement-fixing antibodies with low avidity for measles virus in atypical measles. Nat. Med. 2003, 9, 1209–1213. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Tian, J.H.; Flores, R.; Jacobson, K.; Walker, M.; Portnoff, A.; Gueber-Xabier, M.; Massare, M.J.; Glenn, G.; Ellingsworth, L.; et al. Flexible RSV Prefusogenic Fusion Glycoprotein Exposes Multiple Neutralizing Epitopes that May Collectively Contribute to Protective Immunity. Vaccines 2020, 8, 607. [Google Scholar] [CrossRef]
- Hotard, A.L.; Shaikh, F.Y.; Lee, S.; Yan, D.; Teng, M.N.; Plemper, R.K.; Crowe, J.E.; Moore, M.L. A Stabilized Respiratory Syncytial Virus Reverse Genetics System Amenable to Recombination Mediated Mutagenesis. Virology 2012, 434, 129–136. [Google Scholar] [CrossRef]
- Huang, K.; Incognito, L.; Cheng, X.; Ulbrandt, N.D.; Wu, H. Respiratory Syncytial Virus-Neutralizing Monoclonal Antibodies Motavizumab and Palivizumab Inhibit Fusion. J. Virol. 2010, 84, 8132–8140. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Ohrum, K.; Cayatte, C.; Bennett, A.S.; Rajani, G.M.; McTamney, P.; Nacel, K.; Hostetler, L.; Cheng, L.; Ren, K.; O’Day, T.; et al. Immunization with Low Doses of Recombinant Postfusion or Prefusion Respiratory Syncytial Virus F Primes for Vaccine-Enhanced Disease in the Cotton Rat Model Independently of the Presence of a Th1-Biasing (GLA-SE) or Th2-Biasing (Alum) Adjuvant. J. Virol. 2017, 91, e02180-16. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Ohrum, K.; Snell Bennett, A.; Rajani, G.M.; Hostetler, L.; Maynard, S.K.; Lazzaro, M.; Cheng, L.I.; O’Day, T.; Cayatte, C. CD4 + T Cells Drive Lung Disease Enhancement Induced by Immunization with Suboptimal Doses of Respiratory Syncytial Virus Fusion Protein in the Mouse Model. J. Virol. 2019, 93, e00695-19. [Google Scholar] [CrossRef]
- Acosta, P.L.; Caballero, M.T.; Polack, F.P. Brief History and Characterization of Enhanced Respiratory Syncytial Virus Disease. Clin. Vaccine Immunol. CVI 2016, 23, 189–195. [Google Scholar] [CrossRef]
- Ngwuta, J.O.; Chen, M.; Modjarrad, K.; Joyce, M.G.; Kanekiyo, M.; Kumar, A.; Yassine, H.M.; Moin, S.M.; Killikelly, A.M.; Chuang, G.Y.; et al. Prefusion F–specific antibodies determine the magnitude of RSV neutralizing activity in human sera. Sci. Transl. Med. 2015, 7, 309ra162. [Google Scholar] [CrossRef]
- Hoffman, E.A.; Frey, B.L.; Smith, L.M.; Auble, D.T. Formaldehyde Crosslinking: A Tool for the Study of Chromatin Complexes. J. Biol. Chem. 2015, 290, 26404–26411. [Google Scholar] [CrossRef]
- Santana, H.; Espinosa, L.A.; Sánchez, A.; Bolaño Alvarez, A.; Besada, V.; González, L.J. Mass spectrometric and kinetics characterization of modified species of Growth Hormone Releasing Hexapeptide generated under thermal stress in different pH and buffers. J. Pharm. Biomed. Anal. 2021, 194, 113776. [Google Scholar] [CrossRef] [PubMed]
- Raie Jadidi, B.; Erfan-Niya, H.; Ameghi, A. Optimizing the process of inactivating influenza virus subtype H9N2 by formalin in the production of killed avian influenza vaccine. Arch. Razi Inst. 2017, 72, 43–49. [Google Scholar]
- De Gregorio, E.; Rappuoli, R. From empiricism to rational design: A personal perspective of the evolution of vaccine development. Nat. Rev. Immunol. 2014, 14, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Chua, B.Y.; Sekiya, T.; Jackson, D.C. Opinion: Making Inactivated and Subunit-Based Vaccines Work. Viral Immunol. 2018, 31, 150–158. [Google Scholar] [CrossRef]
- Browne, S.K.; Beeler, J.A.; Roberts, J.N. Summary of the Vaccines and Related Biological Products Advisory Committee meeting held to consider evaluation of vaccine candidates for the prevention of respiratory syncytial virus disease in RSV-naïve infants. Vaccine 2020, 38, 101–106. [Google Scholar] [CrossRef]
- Kurt-Jones, E.A.; Popova, L.; Kwinn, L.; Haynes, L.M.; Jones, L.P.; Tripp, R.A.; Walsh, E.E.; Freeman, M.W.; Golenbock, D.T.; Anderson, L.J.; et al. Pattern recognition receptors TLR4 and CD14 mediate response to respiratory syncytial virus. Nat. Immunol. 2000, 1, 398–401. [Google Scholar] [CrossRef]
- Siegrist, C.A.; Pihlgren, M.; Tougne, C.; Efler, S.M.; Morris, M.L.; AlAdhami, M.J.; Cameron, D.W.; Cooper, C.L.; Heathcote, J.; Davis, H.L.; et al. Co-administration of CpG oligonucleotides enhances the late affinity maturation process of human anti-hepatitis B vaccine response. Vaccine 2004, 23, 615–622. [Google Scholar] [CrossRef]
- Singh, S.M.; Alkie, T.N.; Nagy, É.; Kulkarni, R.R.; Hodgins, D.C.; Sharif, S. Delivery of an inactivated avian influenza virus vaccine adjuvanted with poly(D,L-lactic-co-glycolic acid) encapsulated CpG ODN induces protective immune responses in chickens. Vaccine 2016, 34, 4807–4813. [Google Scholar] [CrossRef]
- Klinman, D.M.; Xie, H.; Little, S.F.; Currie, D.; Ivins, B.E. CpG oligonucleotides improve the protective immune response induced by the anthrax vaccination of rhesus macaques. Vaccine 2004, 22, 2881–2886. [Google Scholar] [CrossRef]
- Troy, N.M.; Bosco, A. Respiratory viral infections and host responses insights from genomics. Respir. Res. 2016, 17, 1–12. [Google Scholar] [CrossRef]
- Malinczak, C.A.; Fonseca, W.; Rasky, A.J.; Ptaschinski, C.; Morris, S.; Ziegler, S.F.; Lukacs, N.W. Sex-associated TSLP-induced immune alterations following early-life RSV infection leads to enhanced allergic disease. Mucosal Immunol. 2019, 12, 969–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, M.; Zhang, W.; Yin, Y.-F.; Si, J.-Y.; Zhang, L.-J.; Chen, L.; Lin, X.; Wang, Y.-B.; Zhang, J.; Zheng, Z.-Z.; et al. An Optimized FI-RSV Vaccine Effectively Protects Cotton Rats and BALB/c Mice without Causing Enhanced Respiratory Disease. Viruses 2022, 14, 2085. https://doi.org/10.3390/v14102085
Lin M, Zhang W, Yin Y-F, Si J-Y, Zhang L-J, Chen L, Lin X, Wang Y-B, Zhang J, Zheng Z-Z, et al. An Optimized FI-RSV Vaccine Effectively Protects Cotton Rats and BALB/c Mice without Causing Enhanced Respiratory Disease. Viruses. 2022; 14(10):2085. https://doi.org/10.3390/v14102085
Chicago/Turabian StyleLin, Min, Wei Zhang, Yi-Fan Yin, Jun-Yu Si, Lu-Jing Zhang, Li Chen, Xue Lin, Ying-Bin Wang, Jun Zhang, Zi-Zheng Zheng, and et al. 2022. "An Optimized FI-RSV Vaccine Effectively Protects Cotton Rats and BALB/c Mice without Causing Enhanced Respiratory Disease" Viruses 14, no. 10: 2085. https://doi.org/10.3390/v14102085
APA StyleLin, M., Zhang, W., Yin, Y.-F., Si, J.-Y., Zhang, L.-J., Chen, L., Lin, X., Wang, Y.-B., Zhang, J., Zheng, Z.-Z., & Xia, N.-S. (2022). An Optimized FI-RSV Vaccine Effectively Protects Cotton Rats and BALB/c Mice without Causing Enhanced Respiratory Disease. Viruses, 14(10), 2085. https://doi.org/10.3390/v14102085