

Detection and Molecular Characterization of Enteric Viruses in Bivalve Mollusks Collected in Arraial do Cabo, Rio de Janeiro, Brazil

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Shellfish Sampling
2.2. Viral Recovery Method
2.3. Nucleic Acid Extraction
2.4. Viral Detection
2.5. Molecular Characterization and Phylogenetic Analysis
2.6. Data Analysis
3. Results
3.1. Enteric Viruses Single and Co-Detections in Bivalve Shellfish
3.2. Enteric Viruses Quantification
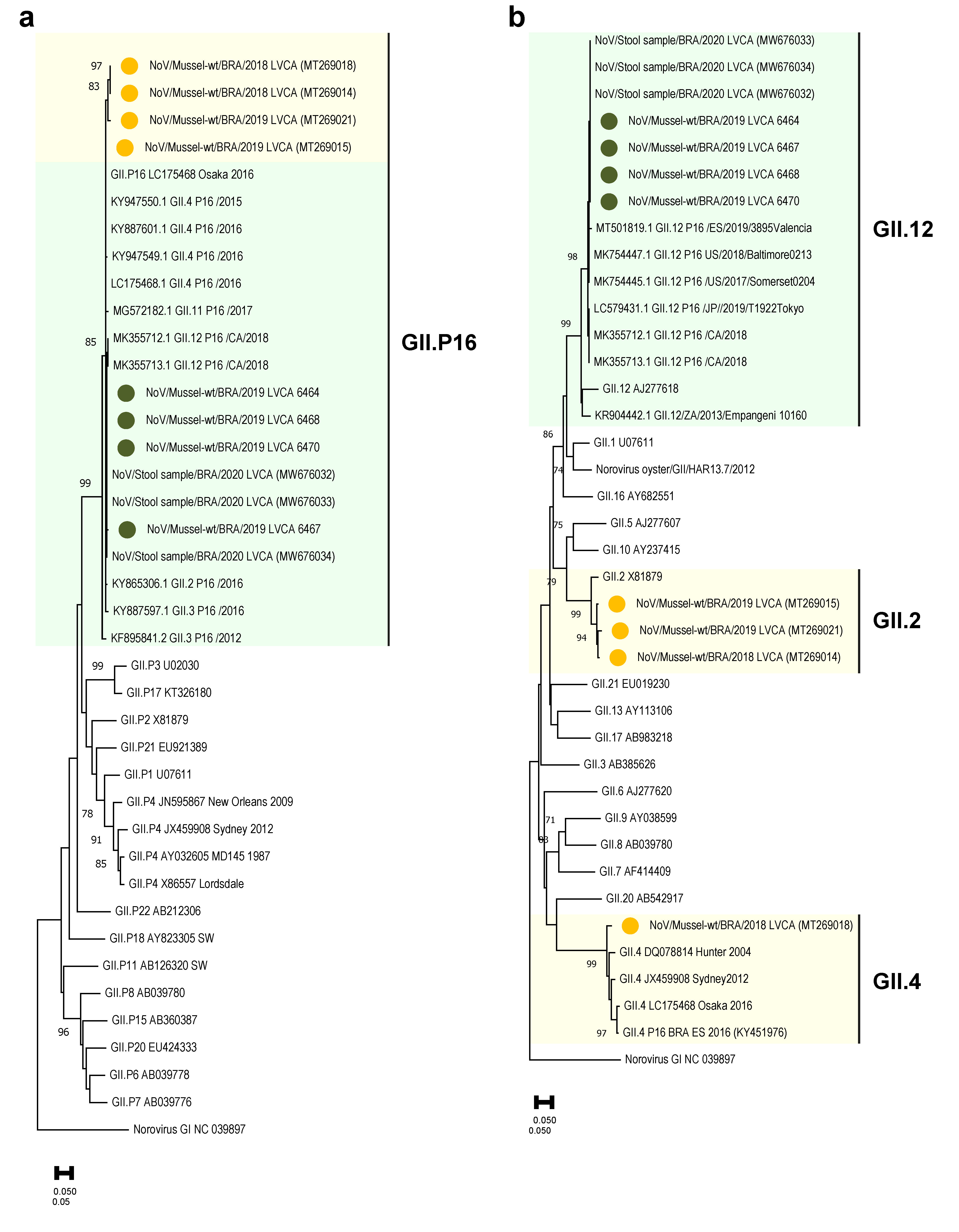
3.3. Enteric Viruses Characterization
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). WHO Estimates of the Global Burden of Foodborne Diseases. 2015. Available online: https://apps.who.int/iris/bitstream/handle/10665/199350/9789241565165_eng.pdf?sequence=1 (accessed on 15 September 2022).
- Fong, T.-T.; Lipp, E.K. Enteric Viruses of Humans and Animals in Aquatic Environments: Health Risks, Detection, and Potential Water Quality Assessment Tools. Microbiol. Mol. Biol. Rev. 2005, 69, 357–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, K.E. Viral Pathogens in Water: Occurrence, Public Health Impact, and Available Control Strategies. Curr. Opin. Virol. 2014, 4, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Pommepuy, M.; Dumas, F.; Caprais, M.P.; Camus, P.; Le Mennec, C.; Parnaudeau, S.; Haugarreau, L.; Sarrette, B.; Vilagines, P.; Pothier, P.; et al. Sewage Impact on Shellfish Microbial Contamination. Water Sci. Technol. 2004, 50, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Gerba, C.P.; Betancourt, W.Q.; Kitajima, M.; Rock, C.M. Reducing Uncertainty in Estimating Virus Reduction by Advanced Water Treatment Processes. Water Res. 2018, 133, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, M.; Iker, B.C.; Pepper, I.L.; Gerba, C.P. Relative Abundance and Treatment Reduction of Viruses during Wastewater Treatment Processes—Identification of Potential Viral Indicators. Sci. Total Environ. 2014, 488–489, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Lanrewaju, A.A.; Enitan-Folami, A.M.; Sabiu, S.; Edokpayi, J.N.; Swalaha, F.M. Global Public Health Implications of Human Exposure to Viral Contaminated Water. Front. Microbiol. 2022, 13, 981896. [Google Scholar] [CrossRef] [PubMed]
- Fouillet, A.; Fournet, N.; Forgeot, C.; Jones, G.; Septfons, A.; Franconeri, L.; Ambert-Balay, K.; Schmidt, J.; Guérin, P.; de Valk, H.; et al. Large Concomitant Outbreaks of Acute Gastroenteritis Emergency Visits in Adults and Food-Borne Events Suspected to Be Linked to Raw Shellfish, France, December 2019 to January 2020. Eurosurveillance 2020, 25, 2000060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fumian, T.M.; Ferreira, F.C.; de Andrade, J.d.S.R.; Canal, N.; Silva Gomes, G.; Teixeira, L.B.; Miagostovich, M.P. Norovirus Foodborne Outbreak Associated With the Consumption of Ice Pop, Southern Brazil, 2020. Food Environ. Virol. 2021, 13, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Meghnath, K.; Hasselback, P.; McCormick, R.; Prystajecky, N.; Taylor, M.; McIntyre, L.; Man, S.; Whitfield, Y.; Warshawsky, B.; McKinley, M.; et al. Outbreaks of Norovirus and Acute Gastroenteritis Associated with British Columbia Oysters, 2016–2017. Food Environ. Virol. 2019, 11, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Le Guyader, F.S.; Bon, F.; DeMedici, D.; Parnaudeau, S.; Bertone, A.; Crudeli, S.; Doyle, A.; Zidane, M.; Suffredini, E.; Kohli, E.; et al. Detection of Multiple Noroviruses Associated with an International Gastroenteritis Outbreak Linked to Oyster Consumption. J. Clin. Microbiol. 2006, 44, 3878–3882. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Gkogka, E.; Le Guyader, F.S.; Loisy-Hamon, F.; Lee, A.; van Lieshout, L.; Marthi, B.; Myrmel, M.; Sansom, A.; Schultz, A.C.; et al. Foodborne Viruses: Detection, Risk Assessment, and Control Options in Food Processing. Int. J. Food Microbiol. 2018, 285, 110–128. [Google Scholar] [CrossRef] [PubMed]
- Bellou, M.; Kokkinos, P.; Vantarakis, A. Shellfish-Borne Viral Outbreaks: A Systematic Review. Food Environ. Virol. 2013, 5, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.K.; Lekshmi, M.; Das, O.; Kumar, S.; Nayak, B.B. Occurrence of Human Enteric Adenoviruses in Fresh Tropical Seafood from Retail Markets and Landing Centers. J. Food Sci. 2019, 84, 2256–2260. [Google Scholar] [CrossRef] [PubMed]
- Rigotto, C.; Victoria, M.; Moresco, V.; Kolesnikovas, C.K.; Corrêa, A.A.; Souza, D.S.M.; Miagostovich, M.P.; Simões, C.M.O.; Barardi, C.R.M. Assessment of Adenovirus, Hepatitis A Virus and Rotavirus Presence in Environmental Samples in Florianopolis, South Brazil. J. Appl. Microbiol. 2010, 109, 1979–1987. [Google Scholar] [CrossRef] [PubMed]
- Keller, R.; Pratte-Santos, R.; Scarpati, K.; Martins, S.A.; Loss, S.M.; Fumian, T.M.; Miagostovich, M.P.; Cassini, S.T. Surveillance of Enteric Viruses and Thermotolerant Coliforms in Surface Water and Bivalves from a Mangrove Estuary in Southeastern Brazil. Food Environ. Virol. 2019, 11, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Sarmento, S.K.; Guerra, C.R.; Malta, F.C.; Coutinho, R.; Miagostovich, M.P.; Fumian, T.M. Human Norovirus Detection in Bivalve Shellfish in Brazil and Evaluation of Viral Infectivity Using PMA Treatment. Mar. Pollut. Bull. 2020, 157, 111315. [Google Scholar] [CrossRef] [PubMed]
- Fusco, G.; Anastasio, A.; Kingsley, D.H.; Amoroso, M.G.; Pepe, T.; Fratamico, P.M.; Cioffi, B.; Rossi, R.; La Rosa, G.; Boccia, F. Detection of Hepatitis A Virus and Other Enteric Viruses in Shellfish Collected in the Gulf of Naples, Italy. Int. J. Environ. Res. Public Health 2019, 16, 2588. [Google Scholar] [CrossRef] [Green Version]
- Vasquez-García, A.; Mejia-Ballesteros, J.E.; de Godoy, S.H.S.; Barbieri, E.; de Sousa, R.L.M.; Fernandes, A.M. Norovirus GII and Astrovirus in Shellfish from a Mangrove Region in Cananéia, Brazil: Molecular Detection and Characterization. Braz. J. Microbiol. 2022, 53, 317–326. [Google Scholar] [CrossRef]
- Dirks, R.A.M.; Jansen, C.C.C.; Hägele, G.; Zwartkruis-Nahuis, A.J.T.; Tijsma, A.S.L.; Boxman, I.L.A. Quantitative Levels of Norovirus and Hepatitis A Virus in Bivalve Molluscs Collected along the Food Chain in the Netherlands, 2013–2017. Int. J. Food Microbiol. 2021, 344, 109089. [Google Scholar] [CrossRef] [PubMed]
- Purpari, G.; Macaluso, G.; Di Bella, S.; Gucciardi, F.; Mira, F.; Di Marco, P.; Lastra, A.; Petersen, E.; La Rosa, G.; Guercio, A. Molecular Characterization of Human Enteric Viruses in Food, Water Samples, and Surface Swabs in Sicily. Int. J. Infect. Dis. 2019, 80, 66–72. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, 324. [Google Scholar] [CrossRef]
- Burkhardt, W.; Calci, K.R. Selective Accumulation May Account for Shellfish-Associated Viral Illness. Appl. Environ. Microbiol. 2000, 66, 1375–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, D.C.; Lovelace, G.L.; Sobsey, M.D. Removal of Escherichia Coli, Enterococcus Fecalis, Coliphage MS2, Poliovirus, and Hepatitis A Virus from Oysters (Crassostrea Virginica) and Hard Shell Clams (Mercinaria Mercinaria) by Depuration. Int. J. Food Microbiol. 2010, 143, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Van Cauwenberghe, L.; Janssen, C.R. Microplastics in Bivalves Cultured for Human Consumption. Environ. Pollut. 2014, 193, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Desdouits, M.; Piquet, J.-C.; Wacrenier, C.; Le Mennec, C.; Parnaudeau, S.; Jousse, S.; Rocq, S.; Bigault, L.; Contrant, M.; Garry, P.; et al. Can Shellfish Be Used to Monitor SARS-CoV-2 in the Coastal Environment? Sci. Total Environ. 2021, 778, 146270. [Google Scholar] [CrossRef] [PubMed]
- Polo, D.; Lois, M.; Fernández-Núñez, M.T.; Romalde, J.L. Detection of SARS-CoV-2 RNA in Bivalve Mollusks and Marine Sediments. Sci. Total Environ. 2021, 786, 147534. [Google Scholar] [CrossRef] [PubMed]
- Mancusi, A.; Capuano, F.; Girardi, S.; Di Maro, O.; Suffredini, E.; Di Concilio, D.; Vassallo, L.; Cuomo, M.C.; Tafuro, M.; Signorelli, D.; et al. Detection of SARS-CoV-2 RNA in Bivalve Mollusks by Droplet Digital RT-PCR (Dd RT-PCR). Int. J. Environ. Res. Public Health 2022, 19, 943. [Google Scholar] [CrossRef] [PubMed]
- GBD 2015 Mortality and Causes of Death Collaborators. Global, Regional, and National Life Expectancy, All-Cause Mortality, and Cause-Specific Mortality for 249 Causes of Death, 1980–2015: A Systematic Analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1459–1544. [Google Scholar] [CrossRef] [Green Version]
- Cohen, A.L.; Platts-Mills, J.A.; Nakamura, T.; Operario, D.J.; Antoni, S.; Mwenda, J.M.; Weldegebriel, G.; Rey-Benito, G.; Oliveira, L.H.d.; Ortiz, C.; et al. Aetiology and Incidence of Diarrhoea Requiring Hospitalisation in Children under 5 Years of Age in 28 Low-Income and Middle-Income Countries: Findings from the Global Pediatric Diarrhea Surveillance Network. BMJ Global Health 2022, 7, e009548. [Google Scholar] [CrossRef]
- Chhabra, P.; de Graaf, M.; Parra, G.I.; Chan, M.C.-W.; Green, K.; Martella, V.; Wang, Q.; White, P.A.; Katayama, K.; Vennema, H.; et al. Updated Classification of Norovirus Genogroups and Genotypes. J. Gen. Virol. 2019, 100, 1393–1406. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, H.B.; Estes, M.K. Rotaviruses: From Pathogenesis to Vaccination. Gastroenterology 2009, 136, 1939–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthijnssens, J.; Attoui, H.; Bányai, K.; Brussaard, C.P.D.; Danthi, P.; del Vas, M.; Dermody, T.S.; Duncan, R.; Fāng, Q.; Johne, R.; et al. ICTV Virus Taxonomy Profile: Sedoreoviridae 2022. J. Gen. Virol. 2022, 103, 001782. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, G.; Goncalves Rossi, L.M.; Forbi, J.C.; de Paula, V.S.; Purdy, M.A.; Xia, G.; Khudyakov, Y.E. Hepatitis A Virus: Host Interactions, Molecular Epidemiology and Evolution. Infect. Genet. Evol. 2014, 21, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Di Cola, G.; Fantilli, A.C.; Pisano, M.B.; Ré, V.E. Foodborne Transmission of Hepatitis A and Hepatitis E Viruses: A Literature Review. Int. J. Food Microbiol. 2021, 338, 108986. [Google Scholar] [CrossRef]
- Benkő, M.; Aoki, K.; Arnberg, N.; Davison, A.J.; Echavarría, M.; Hess, M.; Jones, M.S.; Kaján, G.L.; Kajon, A.E.; Mittal, S.K.; et al. ICTV Virus Taxonomy Profile: Adenoviridae 2022. J. Gen. Virol. 2022, 103, 001721. [Google Scholar] [CrossRef]
- Khanal, S.; Ghimire, P.; Dhamoon, A.S. The Repertoire of Adenovirus in Human Disease: The Innocuous to the Deadly. Biomedicines 2018, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Rames, E.; Roiko, A.; Stratton, H.; Macdonald, J. Technical Aspects of Using Human Adenovirus as a Viral Water Quality Indicator. Water Res. 2016, 96, 308–326. [Google Scholar] [CrossRef]
- Hundesa, A.; Maluquer de Motes, C.; Bofill-Mas, S.; Albinana-Gimenez, N.; Girones, R. Identification of Human and Animal Adenoviruses and Polyomaviruses for Determination of Sources of Fecal Contamination in the Environment. Appl. Environ. Microbiol. 2006, 72, 7886–7893. [Google Scholar] [CrossRef] [Green Version]
- Allander, T.; Tammi, M.T.; Eriksson, M.; Bjerkner, A.; Tiveljung-Lindell, A.; Andersson, B. Cloning of a Human Parvovirus by Molecular Screening of Respiratory Tract Samples. Proc. Natl. Acad. Sci. USA 2005, 102, 12891–12896. [Google Scholar] [CrossRef] [Green Version]
- Malta, F.C.; Varella, R.B.; Guimarães, M.A.A.M.; Miagostovich, M.P.; Fumian, T.M. Human Bocavirus in Brazil: Molecular Epidemiology, Viral Load and Co-Infections. Pathogens 2020, 9, 645. [Google Scholar] [CrossRef]
- Kapoor, A.; Simmonds, P.; Slikas, E.; Li, L.; Bodhidatta, L.; Sethabutr, O.; Triki, H.; Bahri, O.; Oderinde, B.S.; Baba, M.M.; et al. Human Bocaviruses Are Highly Diverse, Dispersed, Recombination Prone, and Prevalent in Enteric Infections. J. Infect. Dis. 2010, 201, 1633–1643. [Google Scholar] [CrossRef]
- Kapoor, A.; Slikas, E.; Simmonds, P.; Chieochansin, T.; Naeem, A.; Shaukat, S.; Masroor Alam, M.; Sharif, S.; Angez, M.; Zaidi, S.; et al. A Newly Identified Bocavirus Species in Human Stool. J. Infect. Dis. 2009, 199, 196–200. [Google Scholar] [CrossRef]
- Arthur, J.L.; Higgins, G.D.; Davidson, G.P.; Givney, R.C.; Ratcliff, R.M. A Novel Bocavirus Associated with Acute Gastroenteritis in Australian Children. PLOS Pathog. 2009, 5, e1000391. [Google Scholar] [CrossRef]
- Polo, D.; Lema, A.; Gándara, E.; Romalde, J.L. Prevalence of Human Bocavirus Infections in Europe. A Systematic Review and Meta-Analysis. Transbound. Emerg. Dis. 2022, 69, 2451–2461. [Google Scholar] [CrossRef]
- Nantachit, N.; Kochjan, P.; Khamrin, P.; Kumthip, K.; Maneekarn, N. Human Bocavirus Genotypes 1, 2, and 3 Circulating in Pediatric Patients with Acute Gastroenteritis in Chiang Mai, Thailand, 2012–2018. J. Infect. Public Health 2021, 14, 179–186. [Google Scholar] [CrossRef]
- Leitão, G.A.A.; Olivares, A.I.O.; Pimenta, Y.C.; Delgado, I.F.; Miagostovich, M.P.; Leite, J.P.G.; de Moraes, M.T.B. Human Bocavirus Genotypes 1 and 2 Detected in Younger Amazonian Children with Acute Gastroenteritis or Respiratory Infections, Respectively. Int. J. Infect. Dis. 2020, 95, 32–37. [Google Scholar] [CrossRef]
- ISO 15216-1:2017; Microbiology of Food a Chain—Horizontal Method for Determination of Hepatitis A Virus and Norovirus in Food Using Real-Time RT-PCR—Part 1: Method for Quantification. International Organization for Standardization: Geneva, Switzerland, 2017; Volume 2017, pp. 1–48.
- Rajal, V.B.; McSwain, B.S.; Thompson, D.E.; Leutenegger, C.M.; Kildare, B.J.; Wuertz, S. Validation of Hollow Fiber Ultrafiltration and Real-Time PCR Using Bacteriophage PP7 as Surrogate for the Quantification of Viruses from Water Samples. Water Res. 2007, 41, 1411–1422. [Google Scholar] [CrossRef]
- Kageyama, T.; Kojima, S.; Shinohara, M.; Uchida, K.; Fukushi, S.; Hoshino, F.B.; Takeda, N.; Katayama, K. Broadly Reactive and Highly Sensitive Assay for Norwalk-like Viruses Based on Real-Time Quantitative Reverse Transcription-PCR. J. Clin. Microbiol. 2003, 41, 1548–1557. [Google Scholar] [CrossRef] [Green Version]
- Kojima, S.; Kageyama, T.; Fukushi, S.; Hoshino, F.B.; Shinohara, M.; Uchida, K.; Natori, K.; Takeda, N.; Katayama, K. Genogroup-Specific PCR Primers for Detection of Norwalk-like Viruses. J. Virol. Methods 2002, 100, 107–114. [Google Scholar] [CrossRef]
- Zeng, S.-Q.; Halkosalo, A.; Salminen, M.; Szakal, E.D.; Puustinen, L.; Vesikari, T. One-Step Quantitative RT-PCR for the Detection of Rotavirus in Acute Gastroenteritis. J. Virol. Methods 2008, 153, 238–240. [Google Scholar] [CrossRef]
- Iturriza Gómara, M.; Wong, C.; Blome, S.; Desselberger, U.; Gray, J. Molecular Characterization of VP6 Genes of Human Rotavirus Isolates: Correlation of Genogroups with Subgroups and Evidence of Independent Segregation. J. Virol. 2002, 76, 6596–6601. [Google Scholar] [CrossRef] [Green Version]
- Costafreda, M.I.; Bosch, A.; Pintó, R.M. Development, Evaluation, and Standardization of a Real-Time TaqMan Reverse Transcription-PCR Assay for Quantification of Hepatitis A Virus in Clinical and Shellfish Samples. Appl. Environ. Microbiol. 2006, 72, 3846–3855. [Google Scholar] [CrossRef] [Green Version]
- Hernroth, B.E.; Conden-Hansson, A.-C.; Rehnstam-Holm, A.-S.; Girones, R.; Allard, A.K. Environmental Factors Influencing Human Viral Pathogens and Their Potential Indicator Organisms in the Blue Mussel, Mytilus Edulis: The First Scandinavian Report. Appl. Environ. Microbiol. 2002, 68, 4523–4533. [Google Scholar] [CrossRef] [Green Version]
- Allard, A.; Albinsson, B.; Wadell, G. Rapid Typing of Human Adenoviruses by a General PCR Combined with Restriction Endonuclease Analysis. J. Clin. Microbiol. 2001, 39, 498–505. [Google Scholar] [CrossRef] [Green Version]
- Kantola, K.; Sadeghi, M.; Antikainen, J.; Kirveskari, J.; Delwart, E.; Hedman, K.; Söderlund-Venermo, M. Real-Time Quantitative PCR Detection of Four Human Bocaviruses. J. Clin. Microbiol. 2010, 48, 4044–4050. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Gutierrez, M.B.; Fialho, A.M.; Maranhão, A.G.; Malta, F.C.; Andrade, J.d.S.R.d.; Assis, R.M.S.d.; Miagostovich, M.P.; Leite, J.P.G.; Machado Fumian, T. Rotavirus A in Brazil: Molecular Epidemiology and Surveillance during 2018–2019. Pathogens 2020, 9, 515. [Google Scholar] [CrossRef]
- Nascimento, L.G.; Fialho, A.M.; Andrade, J.d.S.R.; Assis, R.M.S.; Fumian, T.M. Human Enteric Adenovirus F40/41 as a Major Cause of Acute Gastroenteritis in Children in Brazil, 2018 to 2020. Sci. Rep. 2022, 12, 11220. [Google Scholar] [CrossRef]
- Sarmento, S.K.; de Andrade, J.d.S.R.; Miagostovich, M.P.; Fumian, T.M. Virological and Epidemiological Features of Norovirus Infections in Brazil, 2017–2018. Viruses 2021, 13, 1724. [Google Scholar] [CrossRef]
- Staggemeier, R.; Heck, T.M.S.; Demoliner, M.; Ritzel, R.G.F.; Röhnelt, N.M.S.; Girardi, V.; Venker, C.A.; Spilki, F.R. Enteric Viruses and Adenovirus Diversity in Waters from 2016 Olympic Venues. Sci. Total Environ. 2017, 586, 304–312. [Google Scholar] [CrossRef]
- Assis, A.S.F.; Fumian, T.M.; Miagostovich, M.P.; Drumond, B.P.; da Rosa, E.; Silva, M.L. Adenovirus and Rotavirus Recovery from a Treated Effluent through an Optimized Skimmed-Milk Flocculation Method. Environ. Sci. Pollut. Res. Int. 2018, 25, 17025–17032. [Google Scholar] [CrossRef]
- Fumian, T.M.; Fioretti, J.M.; Lun, J.H.; Dos Santos, I.A.L.; White, P.A.; Miagostovich, M.P. Detection of Norovirus Epidemic Genotypes in Raw Sewage Using next Generation Sequencing. Environ. Int. 2019, 123, 282–291. [Google Scholar] [CrossRef]
- Suffredini, E.; Le, Q.H.; Di Pasquale, S.; Pham, T.D.; Vicenza, T.; Losardo, M.; To, K.A.; De Medici, D. Occurrence and Molecular Characterization of Enteric Viruses in Bivalve Shellfish Marketed in Vietnam. Food Control 2020, 108, 106828. [Google Scholar] [CrossRef]
- Tan, M.T.H.; Ho, S.X.; Chu, J.J.H.; Li, D. Application of Virome Capture Sequencing in Shellfish Sold at Retail Level in Singapore. Lett. Appl. Microbiol. 2021, 73, 486–494. [Google Scholar] [CrossRef]
- Mesquita, J.R.; Vaz, L.; Cerqueira, S.; Castilho, F.; Santos, R.; Monteiro, S.; Manso, C.F.; Romalde, J.L.; Nascimento, M.S.J. Norovirus, Hepatitis A Virus and Enterovirus Presence in Shellfish from High Quality Harvesting Areas in Portugal. Food Microbiol. 2011, 28, 936–941. [Google Scholar] [CrossRef]
- Lowther, J.A.; Gustar, N.E.; Powell, A.L.; Hartnell, R.E.; Lees, D.N. Two-Year Systematic Study to Assess Norovirus Contamination in Oysters from Commercial Harvesting Areas in the United Kingdom. Appl. Environ. Microbiol. 2012, 78, 5812–5817. [Google Scholar] [CrossRef] [Green Version]
- Lowther, J.A.; Gustar, N.E.; Powell, A.L.; O’Brien, S.; Lees, D.N. A One-Year Survey of Norovirus in UK Oysters Collected at the Point of Sale. Food Environ. Virol. 2018, 10, 278–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, O.; Lekshmi, M.; Kumar, S.; Nayak, B.B. Incidence of Norovirus in Tropical Seafood Harbouring Fecal Indicator Bacteria. Mar. Pollut. Bull. 2020, 150, 110777. [Google Scholar] [CrossRef]
- Benabbes, L.; Ollivier, J.; Schaeffer, J.; Parnaudeau, S.; Rhaissi, H.; Nourlil, J.; Le Guyader, F.S. Norovirus and Other Human Enteric Viruses in Moroccan Shellfish. Food Environ. Virol. 2013, 5, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DePaola, A.; Jones, J.L.; Woods, J.; Burkhardt, W.; Calci, K.R.; Krantz, J.A.; Bowers, J.C.; Kasturi, K.; Byars, R.H.; Jacobs, E.; et al. Bacterial and Viral Pathogens in Live Oysters: 2007 United States Market Survey. Appl. Environ. Microbiol. 2010, 76, 2754–2768. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, J.; Le Saux, J.-C.; Lora, M.; Atmar, R.L.; Le Guyader, F.S. Norovirus Contamination on French Marketed Oysters. Int. J. Food Microbiol. 2013, 166, 244–248. [Google Scholar] [CrossRef] [Green Version]
- Seo, D.J.; Lee, M.H.; Son, N.R.; Seo, S.; Lee, K.B.; Wang, X.; Choi, C. Seasonal and Regional Prevalence of Norovirus, Hepatitis A Virus, Hepatitis E Virus, and Rotavirus in Shellfish Harvested from South Korea. Food Control 2014, 41, 178–184. [Google Scholar] [CrossRef]
- Tao, J.; Chunhui, H.; Fanning, S.; Nan, L.; Jiahui, W.; Hongyuan, Z.; Jing, Z.; Fengqin, L. Norovirus Contamination in Retail Oysters from Beijing and Qingdao, China. Food Control 2018, 86, 415–419. [Google Scholar] [CrossRef]
- Torok, V.; Hodgson, K.; McLeod, C.; Tan, J.; Malhi, N.; Turnbull, A. National Survey of Foodborne Viruses in Australian Oysters at Production. Food Microbiol. 2018, 69, 196–203. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, D.; Zhang, Z.; Hewitt, J.; Li, X.; Hou, P.; Wang, D.; Wu, Q. Surveillance of Human Norovirus in Oysters Collected from Production Area in Shandong Province, China during 2017–2018. Food Control 2021, 121, 107649. [Google Scholar] [CrossRef]
- Fusco, G.; Di Bartolo, I.; Cioffi, B.; Ianiro, G.; Palermo, P.; Monini, M.; Amoroso, M.G. Prevalence of Foodborne Viruses in Mussels in Southern Italy. Food Environ. Virol. 2017, 9, 187–194. [Google Scholar] [CrossRef]
- Vilariño, M.L.; Le Guyader, F.S.; Polo, D.; Schaeffer, J.; Kröl, J.; Romalde, J.L. Assessment of Human Enteric Viruses in Cultured and Wild Bivalve Molluscs. Int. Microbiol. 2009, 12, 145–151. [Google Scholar]
- Le Guyader, F.S.; Atmar, R.L.; Le Pendu, J. Transmission of Viruses through Shellfish: When Specific Ligands Come into Play. Curr. Opin. Virol. 2012, 2, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Maalouf, H.; Zakhour, M.; Le Pendu, J.; Le Saux, J.-C.; Atmar, R.L.; Le Guyader, F.S. Distribution in Tissue and Seasonal Variation of Norovirus Genogroup I and II Ligands in Oysters. Appl. Environ. Microbiol. 2010, 76, 5621–5630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansman, G.S.; Oka, T.; Li, T.-C.; Nishio, O.; Noda, M.; Takeda, N. Detection of Human Enteric Viruses in Japanese Clams. J. Food Prot. 2008, 71, 1689–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, J.; Pinto, R.N.; Vieira, C.B.; de Abreu Corrêa, A. Detection and Quantification of Human Adenovirus (HAdV), JC Polyomavirus (JCPyV) and Hepatitis A Virus (HAV) in Recreational Waters of Niterói, Rio de Janeiro, Brazil. Mar. Pollut. Bull. 2018, 133, 240–245. [Google Scholar] [CrossRef]
- Miagostovich, M.P.; Ferreira, F.F.M.; Guimarães, F.R.; Fumian, T.M.; Diniz-Mendes, L.; Luz, S.L.B.; Silva, L.A.; Leite, J.P.G. Molecular Detection and Characterization of Gastroenteritis Viruses Occurring Naturally in the Stream Waters of Manaus, Central Amazonia, Brazil. Appl. Environ. Microbiol. 2008, 74, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Pedrosa de Macena, L.D.G.; Castiglia Feitosa, R.; Vieira, C.B.; Araújo, I.T.; Taniuchi, M.; Miagostovich, M.P. Microbiological Assessment of an Urban Lagoon System in the Coastal Zone of Rio de Janeiro, Brazil. Environ Sci. Pollut. Res. Int. 2021, 28, 1170–1180. [Google Scholar] [CrossRef]
- Victoria, M.; Fumian, T.M.; Rocha, M.S.; Dalmao, F.; Leite, J.P.G.; Girones, R.; Miagostovich, M.P. Gastroenteric Virus Dissemination and Influence of Rainfall Events in Urban Beaches in Brazil. J. Appl. Microbiol. 2014, 117, 1210–1218. [Google Scholar] [CrossRef]
- Gularte, J.S.; Girardi, V.; Demoliner, M.; de Souza, F.G.; Filippi, M.; Eisen, A.K.A.; Mena, K.D.; de Quevedo, D.M.; Rigotto, C.; de Barros, M.P.; et al. Human Mastadenovirus in Water, Sediment, Sea Surface Microlayer, and Bivalve Mollusk from Southern Brazilian Beaches. Mar. Pollut. Bull. 2019, 142, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Hatib, A.; Hassou, N.; Benani, A.; Hafid, J.E.; Ennaji, M.M. Molecular Detection of Rotavirus in Mollusks from the Oued El Maleh Estuary of Mohammedia, Morocco. J. Pure Appl. Microbiol. 2021, 15, 2358–2367. [Google Scholar] [CrossRef]
- Ito, E.; Pu, J.; Miura, T.; Kazama, S.; Nishiyama, M.; Ito, H.; Konta, Y.; Nguyen, G.T.; Omura, T.; Watanabe, T. Weekly Variation of Rotavirus A Concentrations in Sewage and Oysters in Japan, 2014–2016. Pathogens 2019, 8, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mozgovoj, M.; Miño, S.; Barbieri, E.S.; Tort, F.L.; Victoria-Montero, M.; Frydman, C.; Cap, M.; Baron, P.J.; Colina, R.; Matthijnssens, J.; et al. GII.4 Human Norovirus and G8P[1] Bovine-like Rotavirus in Oysters (Crassostrea Gigas) from Argentina. Int. J. Food Microbiol. 2022, 365, 109553. [Google Scholar] [CrossRef]
- Kittigul, L.; Panjangampatthana, A.; Rupprom, K.; Pombubpa, K. Genetic Diversity of Rotavirus Strains Circulating in Environmental Water and Bivalve Shellfish in Thailand. Int. J. Environ. Res. Public Health 2014, 11, 1299–1311. [Google Scholar] [CrossRef] [Green Version]
- Kumthip, K.; Khamrin, P.; Ushijima, H.; Maneekarn, N. Predominance of Human Bocavirus Genotypes 1 and 2 in Oysters in Thailand. Appl. Environ. Microbiol. 2021, 87, e0045621. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, G.; Purpari, G.; Guercio, A.; Di Bella, S.; Gucciardi, F.; Proroga, Y.T.R.; Pisanu, M.; Della Libera, S.; Iaconelli, M.; Suffredini, E. Detection of Human Bocavirus Species 2 and 3 in Bivalve Shellfish in Italy. Appl. Environ. Microbiol. 2018, 84, e0275417. [Google Scholar] [CrossRef] [PubMed]
- Onosi, O.; Upfold, N.S.; Jukes, M.D.; Luke, G.A.; Knox, C. The First Detection of Human Bocavirus Species 2 and 3 in Raw Sewage and Mussels in South Africa. Food Environ. Virol. 2020, 12, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Salvo, M.; Lizasoain, A.; Castells, M.; Bortagaray, V.; Castro, S.; Colina, R.; Tort, F.L.; Victoria, M. Human Bocavirus: Detection, Quantification and Molecular Characterization in Sewage and Surface Waters in Uruguay. Food Environ. Virol. 2018, 10, 193–200. [Google Scholar] [CrossRef]
- Blinkova, O.; Rosario, K.; Li, L.; Kapoor, A.; Slikas, B.; Bernardin, F.; Breitbart, M.; Delwart, E. Frequent Detection of Highly Diverse Variants of Cardiovirus, Cosavirus, Bocavirus, and Circovirus in Sewage Samples Collected in the United States. J. Clin. Microbiol. 2009, 47, 3507–3513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamza, H.; Leifels, M.; Wilhelm, M.; Hamza, I.A. Relative Abundance of Human Bocaviruses in Urban Sewage in Greater Cairo, Egypt. Food Environ. Virol. 2017, 9, 304–313. [Google Scholar] [CrossRef]
- Hamza, I.A.; Jurzik, L.; Wilhelm, M.; Überla, K. Detection and Quantification of Human Bocavirus in River Water. J. Gen. Virol. 2009, 90, 2634–2637. [Google Scholar] [CrossRef] [PubMed]
- Iaconelli, M.; Divizia, M.; Della Libera, S.; Di Bonito, P.; La Rosa, G. Frequent Detection and Genetic Diversity of Human Bocavirus in Urban Sewage Samples. Food Environ. Virol. 2016, 8, 289–295. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, G.; Sanseverino, I.; Della Libera, S.; Iaconelli, M.; Ferrero, V.E.V.; Barra Caracciolo, A.; Lettieri, T. The Impact of Anthropogenic Pressure on the Virological Quality of Water from the Tiber River, Italy. Lett. Appl. Microbiol. 2017, 65, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Soares, L.S.; Lima, A.B.F.; Pantoja, K.C.; Lobo, P.S.; Cruz, J.F.; Guerra, S.F.S.; Bezerra, D.A.M.; Bandeira, R.S.; Mascarenhas, J.D.P. Molecular Epidemiology of Human Bocavirus in Children with Acute Gastroenteritis from North Region of Brazil. J. Med. Microbiol. 2019, 68, 1233–1239. [Google Scholar] [CrossRef]
- Kumthip, K.; Khamrin, P.; Yodmeeklin, A.; Ushijima, H.; Maneekarn, N. Contamination of Human Bocavirus Genotypes 1, 2, 3, and 4 in Environmental Waters in Thailand. Microbiol. Spectr. 2021, 9, e0217821. [Google Scholar] [CrossRef]
- Barclay, L.; Cannon, J.L.; Wikswo, M.E.; Phillips, A.R.; Browne, H.; Montmayeur, A.M.; Tatusov, R.L.; Burke, R.M.; Hall, A.J.; Vinjé, J. Emerging Novel GII.P16 Noroviruses Associated with Multiple Capsid Genotypes. Viruses 2019, 11, 535. [Google Scholar] [CrossRef]
- Pabbaraju, K.; Wong, A.A.; Tipples, G.A.; Pang, X.-L. Emergence of a Novel Recombinant Norovirus GII.P16-GII.12 Strain Causing Gastroenteritis, Alberta, Canada. Emerg. Infect. Dis. 2019, 25, 1556–1559. [Google Scholar] [CrossRef]
- Xiong, Q.; Jiang, H.; Liu, Z.; Peng, J.; Sun, J.; Fang, L.; Li, C.; Qiu, M.; Zhang, X.; Lu, J. Untangling an AGS Outbreak Caused by the Recombinant GII.12[P16] Norovirus With Nanopore Sequencing. Front. Cell. Infect. Microbiol. 2022, 12, 911563. [Google Scholar] [CrossRef]
- Marinho, A.N.R.; Rocha, D.C.C.; Kanai, Y.K.; Alves, C.M.; Costa, D.C.; Sousa, A.H.; Barros, B.C.V.; Bonfim, M.C.M.S.; Mascarenhas, J.D.P. Rotavirus Analyses by SYBR Green Real-Time PCR and Microbiological Contamination in Bivalves Cultivated in Coastal Water of Amazonian Brazil. J. Water Health 2018, 16, 970–979. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, M.B.; de Figueiredo, M.R.; Fialho, A.M.; Cantelli, C.P.; Miagostovich, M.P.; Fumian, T.M. Nosocomial Acute Gastroenteritis Outbreak Caused by an Equine-like G3P[8] DS-1-like Rotavirus and GII.4 Sydney[P16] Norovirus at a Pediatric Hospital in Rio de Janeiro, Brazil, 2019. Hum. Vaccin. Immunother. 2021, 17, 4654–4660. [Google Scholar] [CrossRef]
- Miagostovich, M.P.; Guimarães, F.R.; Vieira, C.B.; Fumian, T.M.; da Gama, N.P.; Victoria, M.; de Oliveira, J.M.; de Oliveira Mendes, A.C.; Gaspar, A.M.C.; Leite, J.P.G. Assessment of Water Quality in a Border Region between the Atlantic Forest and an Urbanised Area in Rio de Janeiro, Brazil. Food Environ. Virol. 2014, 6, 110–115. [Google Scholar] [CrossRef] [Green Version]
- Polo, D.; Varela, M.F.; Romalde, J.L. Detection and Quantification of Hepatitis A Virus and Norovirus in Spanish Authorized Shellfish Harvesting Areas. Int. J. Food Microbiol. 2015, 193, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Macaluso, G.; Guercio, A.; Gucciardi, F.; Di Bella, S.; La Rosa, G.; Suffredini, E.; Randazzo, W.; Purpari, G. Occurrence of Human Enteric Viruses in Shellfish along the Production and Distribution Chain in Sicily, Italy. Foods 2021, 10, 1384. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, G.; Mancini, P.; Bonanno Ferraro, G.; Iaconelli, M.; Veneri, C.; Paradiso, R.; De Medici, D.; Vicenza, T.; Proroga, Y.T.R.; Di Maro, O.; et al. Hepatitis A Virus Strains Circulating in the Campania Region (2015–2018) Assessed through Bivalve Biomonitoring and Environmental Surveillance. Viruses 2020, 13, 16. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, T.M.; Vieira, N.S.G.; Sepp, T.D.S.; Souto, F.J.D. Recent Trends in Hepatitis A Incidence in Brazil. J. Med. Virol. 2020, 92, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Souto, F.J.D.; de Brito, W.I.; Fontes, C.J.F. Impact of the Single-Dose Universal Mass Vaccination Strategy against Hepatitis A in Brazil. Vaccine 2019, 37, 771–775. [Google Scholar] [CrossRef] [PubMed]
- Desdouits, M.; de Graaf, M.; Strubbia, S.; Oude Munnink, B.B.; Kroneman, A.; Le Guyader, F.S.; Koopmans, M.P.G. Novel Opportunities for NGS-Based One Health Surveillance of Foodborne Viruses. One Health Outlook 2020, 2, 14. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Molecular Method | Primers and Probes | Sequence (5′–3′) | Target Region | Reference |
|---|---|---|---|---|---|
| NoV | real-time RT-PCR | COG1F | CGYTGGATGCGNTTYCATGA | ORF1/2 junction | [50] |
| COG1R | CTTAGACGCCATCATCATTYAC | ||||
| RING1 (P) | FAM-AGATYGCGATCYCCTGTCCA-TAMRA | ||||
| COG2F | CARGARBCNATGTTYAGRTGGATGAG | ||||
| COG2R | TCGACGCCATCTTCATTCACA | ||||
| RING2 (P) | HEX-TGGGAGGGCGATCGCAATCT-TAMRA | ||||
| RT-PCR | MON431 (F) | TGGACIAGRGGICCYAAYCA | 3′ORF1 and 5′ORF2 | [51] | |
| G2SKR (R) | CCRCCNGCATRHCCRTTRTACAT | ||||
| RVA | real-time RT-PCR | NSP3-F | ACCATCTWCACRTRACCCTCTATGAG | NSP3 | [52] |
| NSP3-R1 | GGTCACATAACGCCCCTATAGC | ||||
| NSP3 (P) | FAM-AGTTAAAAGCTAACACTGTCAAA-MGB | ||||
| RT-PCR | VP6-F | GACGGVGCRACTACATGGT | VP6 gene | [53] | |
| VP6-R | GTCCAATTCATNCCTGGTGG | ||||
| HVA | real-time RT-PCR | HAV68 (F) | TCACCGCCGTTTGCCTAG | 5′-NCR | [54] |
| HAV240 (R) | GGAGAGCCCTGGAAGAAAG | ||||
| HAV150p (P) | FAM-TTAATTCCTGCAGGTTCAGG-MGB | ||||
| HAdV | real-time PCR | AdF | CATTACATGCACATCGTCCGGG | Hexon gene | [55] |
| AdR | CAGCGGGCGAAACTTGCACCAG | ||||
| AdP1 | FAM-CCGGGCTCAGGTACTCCGAGGCGTCCT-TAMRA | ||||
| PCR | Hex1deg (F) | GCCSCARTGGKCWTACATGCACATC | Hexon gene | [56] | |
| Hex2deg (R) | CAGCACSCCICGRATGTCAAA | ||||
| HBoV | real-time PCR | HBoV1 (F) | CCTATATAAGCTGCTGCACTTCCTG | NS1 | [57] |
| HBoV1 (R) | AAGCCATAGTAGACTCACCACAAG | ||||
| HBoV234 (F) | GCACTTCCGCATYTCGTCAG | ||||
| HBoV3 (R) | GTGGATTGAAAGCCATAATTTGA | ||||
| HBoV24 (R) | AGCAGAAAAGGCCATAGTGTCA | ||||
| (Probe) | FAM-CCAGAGATGTTCACTCGCCG-MGB | ||||
| nested PCR | AK-VP-F1 | CGCCGTGGCTCCTGCTCT | VP1/2 | [42] | |
| AK-VP-R1 | TGTTCGCCATCACAAAAGATGTG | ||||
| AK-VP-F2 | GGCTCCTGCTCTAGGAAATAAAGAG | ||||
| AK-VP-R2 | CCTGCTGTTAGGTCGTTGTTGTATGT |
| Bivalve Shellfish | Collecting Site | Samples (n) | Number of Positive Samples | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Single Detection | Double Detection | Triple Detection | Quadruple Detection | ||||||||||
| NoV * | RVA | HAdV | NoV + RVA | NoV + HAdV | NoV + HBoV | HBoV + HAdV | NoV + RVA + HAdV | NoV + RVA + HBoV | NoV + HAdV + HBoV | NoV + RVA + HAdV + HBoV | |||
| Mussels (Perna perna) | Anjos beach | 22 | 3 | 1 | 2 | 0 | 2 | 0 | 0 | 1 | 0 | 2 | 0 |
| Farol beach | 25 | 4 | 1 | 3 | 2 | 4 | 0 | 1 | 0 | 0 | 3 | 0 | |
| Forno beach | 26 | 6 | 4 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 3 | 0 | |
| Oysters (Pseudochama cristella) | Forno beach | 24 | 5 | 4 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 |
| Total | 97 | 18 | 10 | 5 | 2 | 7 | 2 | 1 | 2 | 1 | 8 | 1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
do Nascimento, L.G.; Sarmento, S.K.; Leonardo, R.; Gutierrez, M.B.; Malta, F.C.; de Oliveira, J.M.; Guerra, C.R.; Coutinho, R.; Miagostovich, M.P.; Fumian, T.M. Detection and Molecular Characterization of Enteric Viruses in Bivalve Mollusks Collected in Arraial do Cabo, Rio de Janeiro, Brazil. Viruses 2022, 14, 2359. https://doi.org/10.3390/v14112359
do Nascimento LG, Sarmento SK, Leonardo R, Gutierrez MB, Malta FC, de Oliveira JM, Guerra CR, Coutinho R, Miagostovich MP, Fumian TM. Detection and Molecular Characterization of Enteric Viruses in Bivalve Mollusks Collected in Arraial do Cabo, Rio de Janeiro, Brazil. Viruses. 2022; 14(11):2359. https://doi.org/10.3390/v14112359
Chicago/Turabian Styledo Nascimento, Lilian Gonçalves, Sylvia Kahwage Sarmento, Raphael Leonardo, Meylin Bautista Gutierrez, Fábio Correia Malta, Jaqueline Mendes de Oliveira, Caroline Rezende Guerra, Ricardo Coutinho, Marize Pereira Miagostovich, and Tulio Machado Fumian. 2022. "Detection and Molecular Characterization of Enteric Viruses in Bivalve Mollusks Collected in Arraial do Cabo, Rio de Janeiro, Brazil" Viruses 14, no. 11: 2359. https://doi.org/10.3390/v14112359
APA Styledo Nascimento, L. G., Sarmento, S. K., Leonardo, R., Gutierrez, M. B., Malta, F. C., de Oliveira, J. M., Guerra, C. R., Coutinho, R., Miagostovich, M. P., & Fumian, T. M. (2022). Detection and Molecular Characterization of Enteric Viruses in Bivalve Mollusks Collected in Arraial do Cabo, Rio de Janeiro, Brazil. Viruses, 14(11), 2359. https://doi.org/10.3390/v14112359