
In Vitro Evaluation of Anti-Rotaviral Activity and Intestinal Toxicity of a Phytotherapeutic Prototype of Achyrocline bogotensis (Kunth) DC.
 , , and
, , and 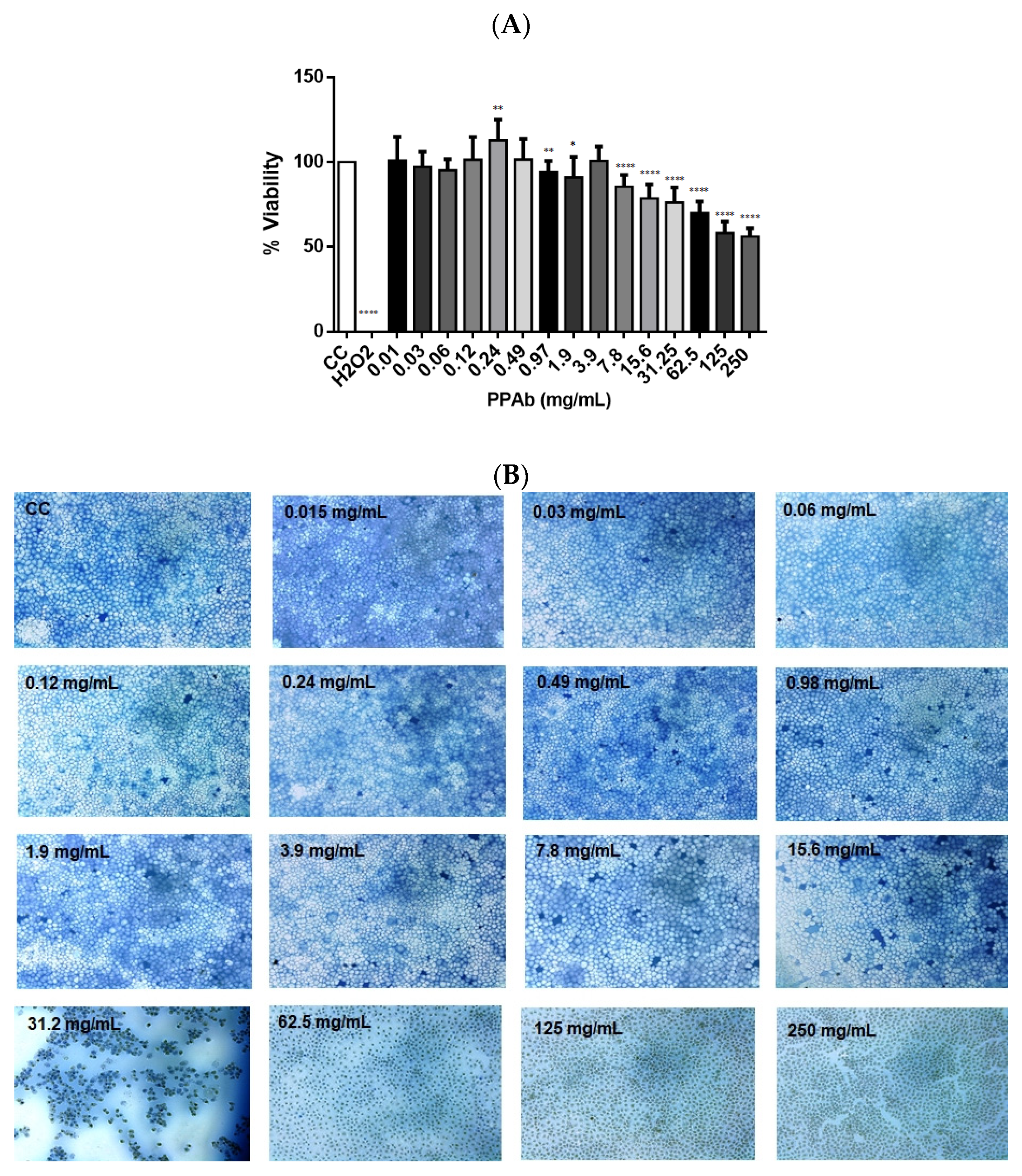{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cytotoxicity of PPAb for MA104 Cells, Susceptible to RV Infection
2.2. Anti-Rotavirus Activity of the PPAb: Virucidal Effect and Viral Yield Reduction
2.2.1. Immunocytochemistry (ICC)
2.2.2. Flow Cytometry
2.3. PPAB Toxicity for Human Intestinal Cells: C2BBe1 Cells
2.3.1. Cytotoxicity Assay
2.3.2. Evaluation of Death Markers Exerted by the PPAb on C2BBe1 Cells: Cytoplasmic Membrane Asymmetry and DNA Fragmentation
3. Results
3.1. Cytotoxic Effect of the PPAb on MA104 Cells
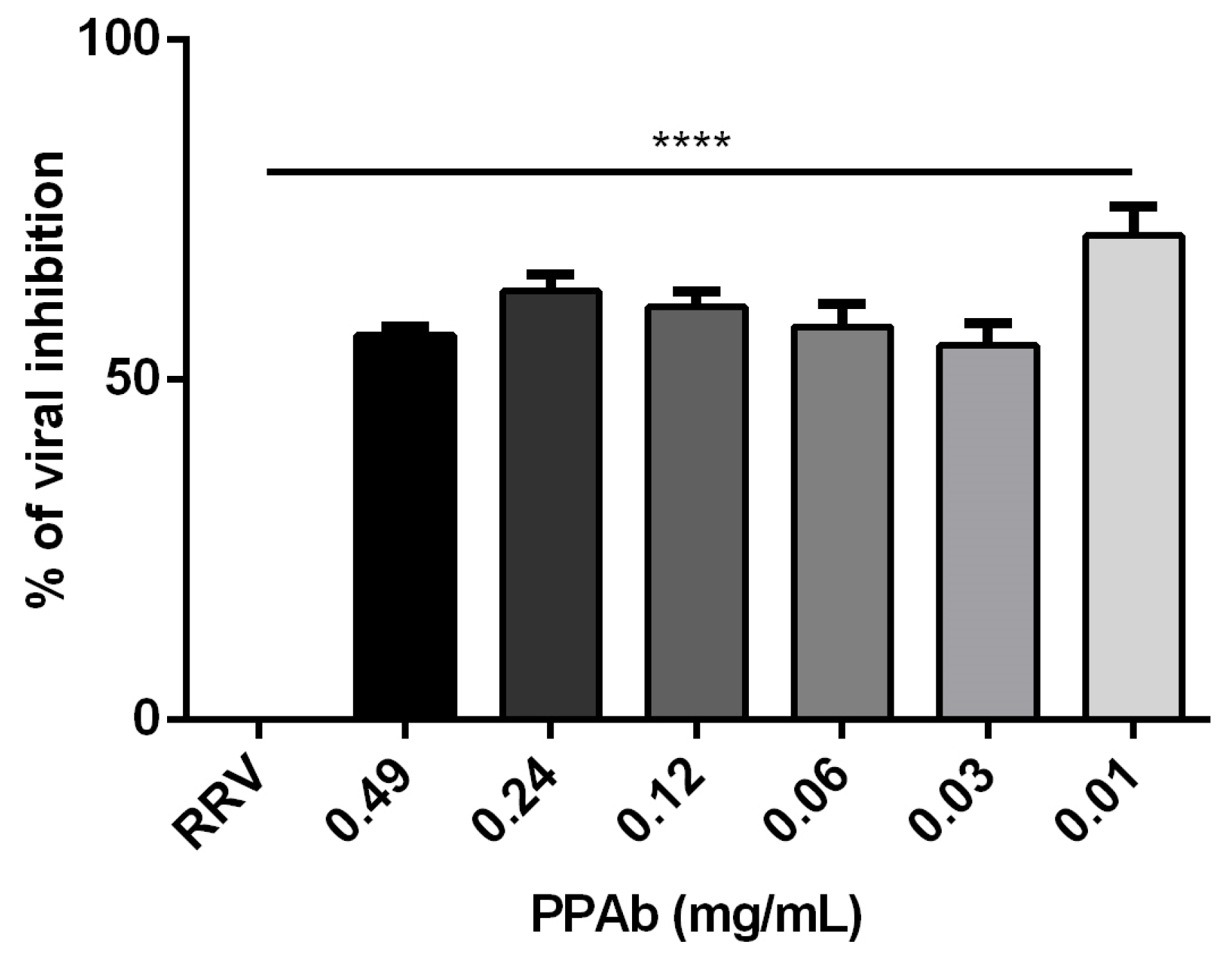
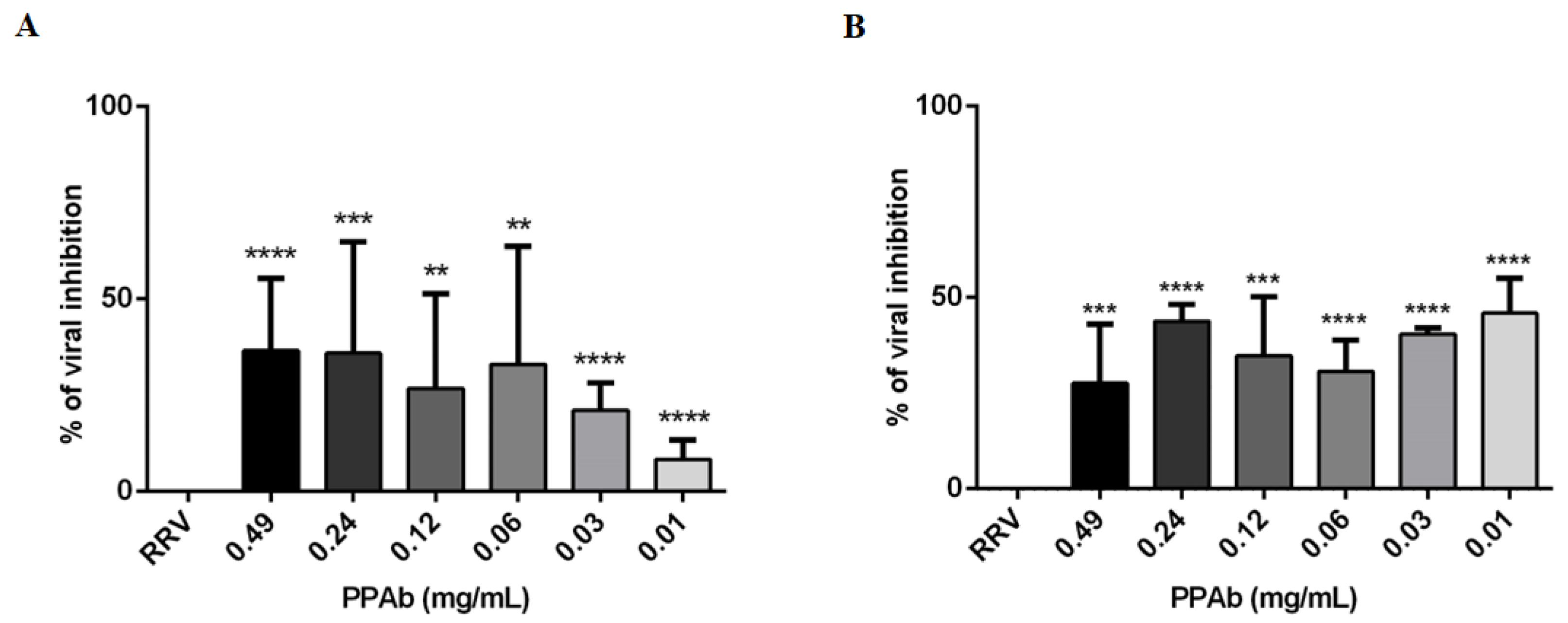
3.2. Anti-Rotavirus Activity of the PPAb
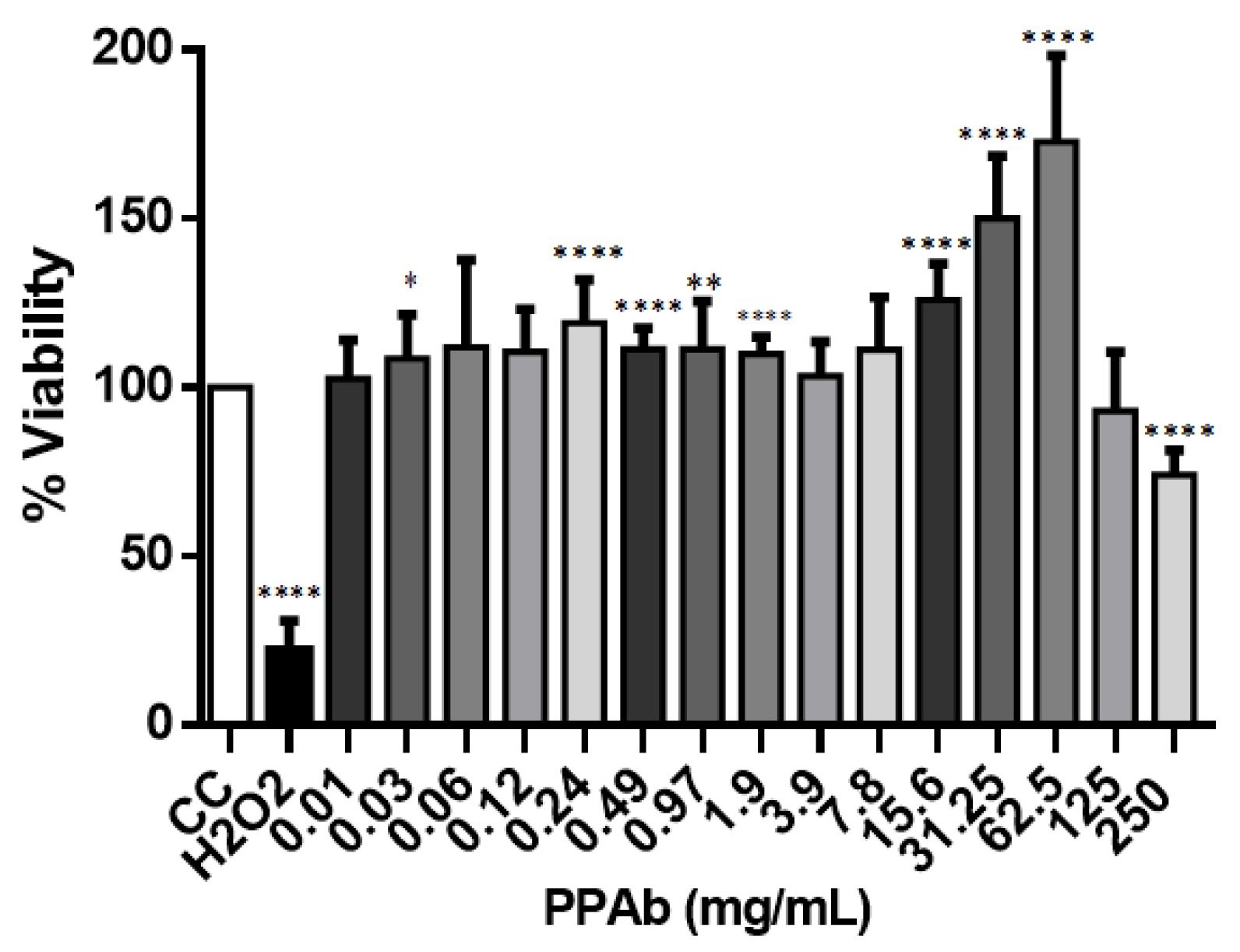
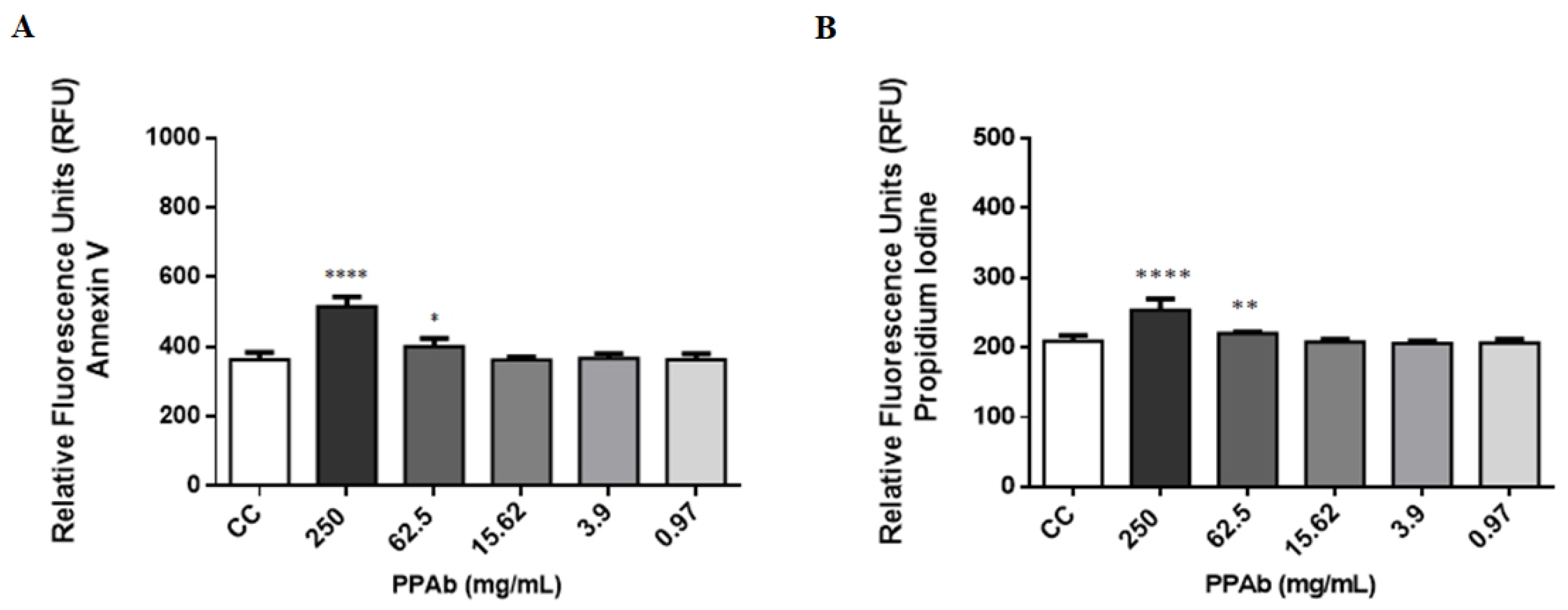
3.3. Intestinal Toxicity of the PPAb on C2BBe1 Cells: Cytotoxicity, Cytoplasmic Membrane Asymmetry and DNA Fragmentation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WGO. WGO Practice Guideline—Acute Diarrhea. Available online: https://www.worldgastroenterology.org/guidelines/global-guidelines/acute-diarrhea#:~:text=Definition%3A%20Acute%20diarrhea%20is%20defined,lasting%20less%20than%2014%20days (accessed on 15 July 2020).
- WHO. Diarrhoeal Disease. Available online: https://www.who.int/news-room/fact-sheets/detail/diarrhoeal-disease (accessed on 21 August 2019).
- Long, C.P.; McDonald, S.M. Rotavirus genome replication: Some assembly required. PLoS Pathog. 2017, 13, e1006242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saurabh, S.; Sircar, S.; Kattoor, J.J.; Ghosh, S.; Kobayashi, N.; Banyai, K.; VinodhKumar, O.R.; De, U.K.; Sahoo, N.R.; Dhama, K.; et al. Analysis of structure-function relationship in porcine rotavirus A enterotoxin gene. J. Vet. Sci. 2018, 19, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Desselberger, U. Rotaviruses. Virus Res. 2014, 190, 75–96. [Google Scholar] [CrossRef] [Green Version]
- WHO Immunization, Vaccines and Biologicals Rotavirus. Available online: https://www.who.int/teams/immunization-vaccines-and-biologicals/diseases/rotavirus (accessed on 15 June 2022).
- Gruber, J.F.; Hille, D.A.; Liu, G.F.; Kaplan, S.S.; Nelson, M.; Goveia, M.G.; Mast, T.C. Heterogeneity of Rotavirus Vaccine Efficacy Among Infants in Developing Countries. Pediatr. Infect. Dis. J. 2017, 36, 72–78. [Google Scholar] [CrossRef]
- Bucardo, F.; Nordgren, J. Impact of vaccination on the molecular epidemiology and evolution of group A rotaviruses in Latin America and factors affecting vaccine efficacy. Infect. Genet. Evol. 2015, 34, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Rose, T.L.; Da Silva, M.F.M.; Goméz, M.M.; Resque, H.R.; Ichihara, M.Y.T.; Volotão, E.D.M.; Leite, J.P.G. Evidence of vaccine-related reassortment of rotavirus, Brazil, 2008–2010. Emerg. Infect. Dis. 2013, 19, 1843–1846. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.S.; Sousa-Júnior, E.C.; Guerra, S.F.S.; Lobo, P.S.; Penha, E.T., Jr.; Lima, A.B.F.; Vinente, C.B.G.; Chagas, E.N.H.; Justino, M.C.A.; Linhares, A.C.; et al. G1P[8] Rotavirus in children with severe diarrhea in the post-vaccine introduction era in Brazil: Evidence of reassortments and structural modifications of the antigenic VP7 and VP4 regions. Infect. Genet. Evol. 2019, 69, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Koch, J.; Harder, T.; von Kries, R.; Wichmann, O. Risk of Intussusception after Rotavirus Vaccination. Dtsch. Arztebl. Int. 2017, 114, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Katherine, Y.W.; Lieu, T.A.; Kulldorff, M.; Martin, D.; McMahill-Walraven, C.N.; Platt, R.; Selvam, N.; Selvan, M.; Lee, G.M.; Nguyen, M. Intussusception risk after rotavirus vaccination in U.S. infants. N. Engl. J. Med. 2014, 370, 503–512. [Google Scholar]
- Bravo, L.; Chitraka, A.; Liu, A.; Choudhury, J.; Kumar, K.; Berezo, L.; Cimafranca, L.; Chatterjee, P.; Garg, P.; Siriwardene, P.; et al. Reactogenicity and safety of the human rotavirus vaccine, Rotarix™ in The Philippines, Sri Lanka, and India: A post-marketing surveillance study. Hum. Vaccines Immunother. 2014, 10, 2276–2283. [Google Scholar] [CrossRef] [Green Version]
- Haber, P.; Amin, M.; Ng, C.; Weintraub, E.; McNeil, M.M. Reports of lower respiratory tract infection following dose 1 of RotaTeq and Rotarix vaccines to the Vaccine Adverse Event Reporting System (VAERS), 2008–2016. Hum. Vaccines Immunother. 2018, 14, 2755–2759. [Google Scholar] [CrossRef] [PubMed]
- Batista, A.; Behr, D.; De Carvalho, P.; Caldas, S.; Alves, D.; Guerra, M.E.; Resende, M.G.; De Souza, C.P.; Gontijo, C.; De Almeidaa, V. Screening of Brazilian medicinal plants for antiviral activity against rotavirus. J. Ethnophamacol. 2012, 141, 975–981. [Google Scholar]
- Takahashi, K.; Matsuda, M.; Ohashi, K.; Taniguchi, K.; Nakagomi, O.; Abe, Y.; Mori, S.; Sato, N.; Okutani, K.; Shigeta, S. Analysis of anti-rotavirus activity of extract from Stevia rebaudiana. Antivir. Res. 2001, 49, 15–24. [Google Scholar] [CrossRef]
- Tam, K.I.; Roner, M.R. Characterization of in vivo anti-rotavirus activities of saponin extracts from Quillaja saponaria Molina. Antivir. Res. 2011, 90, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calixto, J. Efficacy, safety, quality control, marketing and regulatory guidelines for herbal medicines (phytotherapeutic agents). Braz. J. Med. Biol. Res. 2000, 33, 179–189. [Google Scholar] [CrossRef]
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; Tsigalou, C.; Bezirtzoglou, E. Towards Advances in Medicinal Plant Antimicrobial Activity: A Review Study on Challenges and Future Perspectives. Microorganisms 2021, 9, 2041. [Google Scholar] [CrossRef]
- WHO. General Guidelines for Methodologies on Research and Evaluation of Traditional Medicine; World Health Organization: Geneva, Switzerland, 2000. [Google Scholar]
- Téllez, M.A.; Téllez, A.N.; Velez, F.; Ulloa, J.C. In vitro antiviral activity against rotavirus and astrovirus infection exerted by substances obtained from Achyrocline bogotensis (Kunth) DC. (Compositae). BMC Complement. Altern. Med. 2015, 15, 428. [Google Scholar] [CrossRef] [Green Version]
- Valle, C. Evaluation of the Anti-Rotavirus Effect Exerted by a Pilot Phytotherapeutic Product; Academic Undergraduate Research Work; Pontificia Universidad Javeriana: Bogotá, Colombia, 2016. [Google Scholar]
- Ulloa, J.C.; Téllez, M.A.; Tellez, A.N. Fracción bioactiva de Achyrocline bogotensis (Kunth) DC. (Compositae) con actividad antiviral. 16-017515-0000-0000, 3 August 2017. [Google Scholar]
- Carreño-Torres, J.J.; Gutiérrez, M.; Arias, C.F.; López, S.; Isa, P. Characterization of viroplasm formation during the early stages of rotavirus infection. Virol. J. 2010, 7, 350. [Google Scholar] [CrossRef] [Green Version]
- WHO. WHO Guidelines for Assesing Quality of Herbal Medicines with Reference to Contaminants and Residues; WHO Library Cataloguing-in-Publication Data; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Retta, D.; Dellacassa, E.; Villamil, J.; Suárez, S.A.; Bandoni, A.L. Marcela, a promising medicinal and aromatic plant from Latin America: A review. Ind. Crops Prod. 2012, 38, 27–38. [Google Scholar] [CrossRef]
- Chattopadhyay, D.; Naik, T.N. Antivirals of ethnomedicinal origin: Structure-activity relationship and scope. Mini Rev. Med. Chem. 2007, 7, 275–301. [Google Scholar] [CrossRef]
- Seo, D.; Choi, C. Inhibitory mechanism of five natural flavonoids against murine norovirus. Phytomedicine 2017, 1, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Evers, D.L.; Chao, C.-F.; Wang, X.; Zhang, Z.; Huong, S.-M.; Huang, E.-S. Human cytomegalovirus-inhibitory flavonoids: Studies on antiviral activity and mechanism of action. Antivir. Res. 2005, 68, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Alfajaro, M.M.; Rho, M.C.; Kim, H.-J.; Park, J.-G.; Kim, D.-S.; Hosmillo, M.; Son, K.-Y.; Lee, J.-H.; Park, S.-I.; Kang, M.-I.; et al. Anti-rotavirus effects by combination therapy of stevioside and Sophora flavescens extract. Res. Vet. Sci. 2014, 96, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.J.; Kim, H.H.; Ryu, Y.B.; Kim, J.H.; Jeong, H.J.; Lee, S.W.; Chang, J.S.; Cho, K.O.; Rho, M.C.; Park, S.J.; et al. In vitro anti-rotavirus activity of polyphenol compounds isolated from the roots of Glycyrrhiza uralensis. Bioorg. Med. Chem. 2010, 18, 7668–7674. [Google Scholar] [CrossRef]
- Savi, L.A.; Caon, T.; de Oliveira, A.P.; Sobottka, A.M.; Werner, W.; Reginatto, F.H.; Schenkel, E.P.; Barardi, C.R.M.; Simões, C.M.O. Evaluation of antirotavirus activity of flavonoids. Fitoterapia 2010, 81, 1142–1146. [Google Scholar] [CrossRef]
- Bae, E.-A.; Han, M.J.; Lee, M.; Kim, D.-H. In vitro inhibitory effect of some flavonoids on rotavirus infectivity. Biol. Pharm. Bull. 2000, 23, 1122–1124. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.M.; Wood, R.C.; Wyatt, J.E.; Pendleton, M.H.; Torrenegra, R.D.; Rodriguez, O.E.; Harirforoosh, S.; Ballester, M.; Lightner, J.; Krishnan, K.; et al. Anti-Neoplastic Activity of Two Flavone Isomers Derived from Gnaphalium elegans and Achyrocline bogotensis. PLoS ONE 2012, 7, e39806. [Google Scholar] [CrossRef] [Green Version]
- Torrenegra, R.; Escarria, S.; Tenorio, E.; Achembach, H. Estudio Fitoquímico del Achyrocline bogotensis. Rev. Latinoam. Química 1982, 13, 75–76. [Google Scholar]
- Buccigrossi, V.; Laudiero, G.; Russo, C.; Miele, E.; Sofia, M.; Monini, M.; Ruggeri, F.M.; Guarino, A. Chloride secretion induced by rotavirus is oxidative stress-dependent and inhibited by Saccharomyces boulardii in human enterocytes. PLoS ONE 2014, 11, e99830. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, C.A.; Acosta, O. Inflammatory and oxidative stress in rotavirus infection. World J. Virol. 2016, 5, 38–62. [Google Scholar] [CrossRef]
- FDA. Guidance for Industry Antiviral Product Development; Food and Drug Administration: Rockville, MD, USA, 2006; Volume 20857, p. 17. [Google Scholar]
- Kim, H.-H.; Kwon, H.J.; Ryu, Y.B.; Chang, J.S.; Cho, K.-O.; Hosmillo, M.; Rho, M.-C.; Park, S.-J.; Lee, W.S. Antiviral activity of Alpinia katsumadai extracts against rotaviruses. Res. Vet. Sci. 2012, 92, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Ministerio-de-Protección-Social. Vademécum Colombiano de Plantas Medicinales, 1st ed.; Ministerio de Salud y Protección Social: Bogotá, Colombia, 2008; p. 241.
- Ahmet, A.; Aktay, G.; Yesilada, E. A Guidance Manual for the Toxicity Assessment of Traditional Herbal Medicines. Nat. Prod. Commun. 2016, 11, 1763–1773. [Google Scholar]
- Naha, P.C.; Chhour, P.; Cormode, D.P. Systematic in vitro toxicological screening of gold nanoparticles designed for nanomedicine applications. Toxicol. Vitr. 2015, 29, 1445–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohal, I.S.; O’Fallon, K.S.; Gaines, P.; Demokritou, P.; Bello, D. Ingested engineered nanomaterials: State of science in nanotoxicity testing and future research needs. Part. Fibre Toxicol. 2018, 15, 29. [Google Scholar] [CrossRef]
- Cieniak, C.; Liu, R.; Fottinger, A.; Smiley, S.A.; Guerrero-Analco, J.A.; Bennett, S.A.; Haddad, P.S.; Cuerrier, A.; Saleem, A.; Arnason, J.T.; et al. In vitro inhibition of metabolism but not transport of gliclazide and repaglinide by Cree medicinal plant extracts. J. Ethnopharmacol. 2013, 150, 1087–1095. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez, M.-C.; Méndez, K.; Castelblanco-Mora, A.; Quijano, S.; Ulloa, J. In Vitro Evaluation of Anti-Rotaviral Activity and Intestinal Toxicity of a Phytotherapeutic Prototype of Achyrocline bogotensis (Kunth) DC. Viruses 2022, 14, 2394. https://doi.org/10.3390/v14112394
Ramírez M-C, Méndez K, Castelblanco-Mora A, Quijano S, Ulloa J. In Vitro Evaluation of Anti-Rotaviral Activity and Intestinal Toxicity of a Phytotherapeutic Prototype of Achyrocline bogotensis (Kunth) DC. Viruses. 2022; 14(11):2394. https://doi.org/10.3390/v14112394
Chicago/Turabian StyleRamírez, María-Camila, Kelly Méndez, Alicia Castelblanco-Mora, Sandra Quijano, and Juan Ulloa. 2022. "In Vitro Evaluation of Anti-Rotaviral Activity and Intestinal Toxicity of a Phytotherapeutic Prototype of Achyrocline bogotensis (Kunth) DC." Viruses 14, no. 11: 2394. https://doi.org/10.3390/v14112394
APA StyleRamírez, M. -C., Méndez, K., Castelblanco-Mora, A., Quijano, S., & Ulloa, J. (2022). In Vitro Evaluation of Anti-Rotaviral Activity and Intestinal Toxicity of a Phytotherapeutic Prototype of Achyrocline bogotensis (Kunth) DC. Viruses, 14(11), 2394. https://doi.org/10.3390/v14112394