
Chitinase-3-like Protein 1 Is Associated with Poor Virologic Control and Immune Activation in Children Living with HIV

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Clinical Definitions
2.3. Measurement of Biomarker Levels
2.4. Statistical Analysis
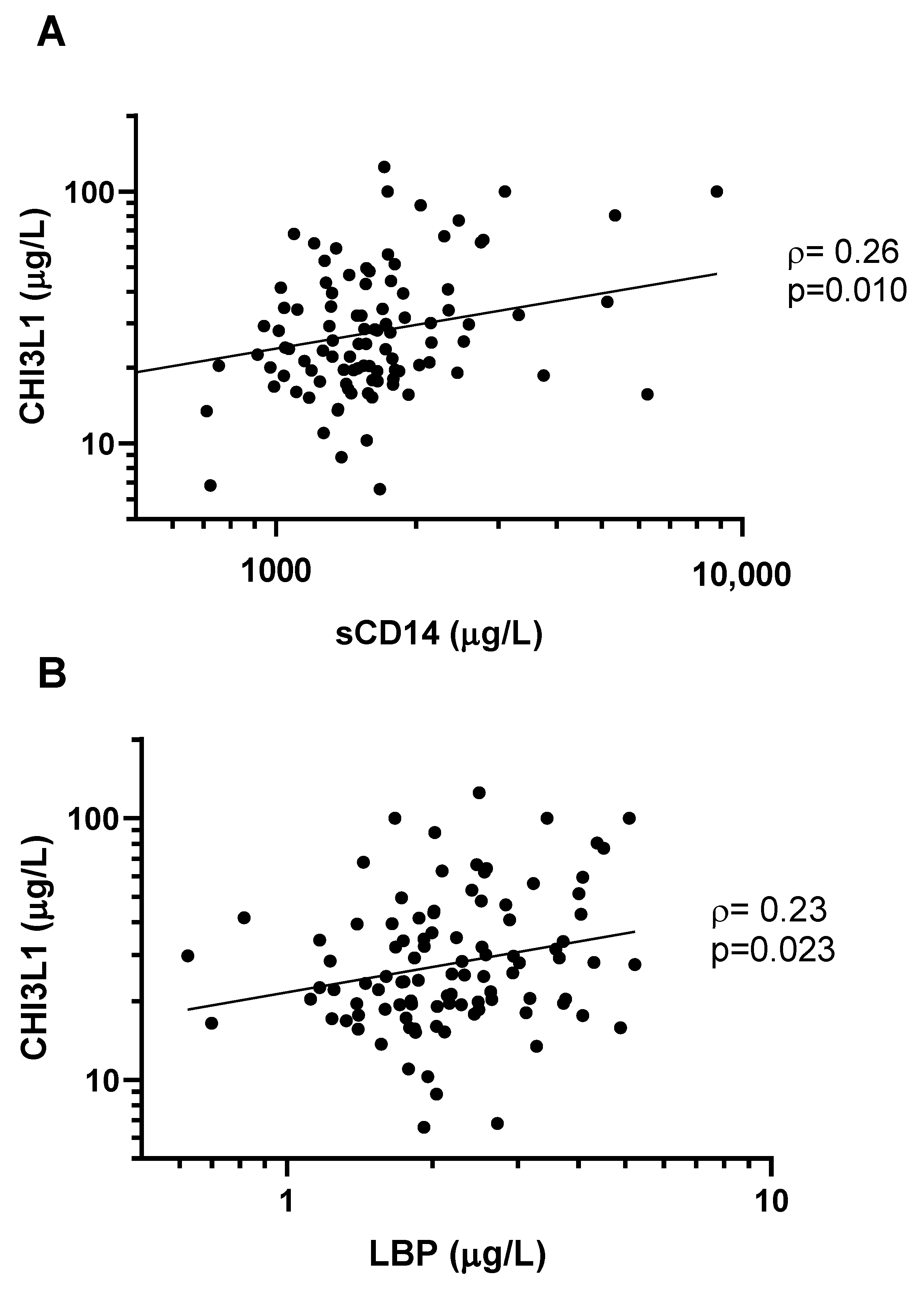
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Grinspoon, S.K.; Grunfeld, C.; Kotler, D.P.; Currier, J.S.; Lundgren, J.D.; Dubé, M.P.; Lipshultz, S.E.; Hsue, P.Y.; Squires, K.; Schambelan, M.; et al. State of the Science Conference. Circulation 2008, 118, 198–210. [Google Scholar] [CrossRef]
- Soehnlein, O.; Steffens, S.; Hidalgo, A.; Weber, C. Neutrophils as protagonists and targets in chronic inflammation. Nat. Rev. Immunol. 2017, 17, 248–261. [Google Scholar] [CrossRef]
- Campillo-Gimenez, L.; Casulli, S.; Dudoit, Y.; Seang, S.; Carcelain, G.; Lambert-Niclot, S.; Appay, V.; Autran, B.; Tubiana, R.; Elbim, C. Neutrophils in antiretroviral therapy-controlled HIV demonstrate hyperactivation associated with a specific IL-17/IL-22 environment. J. Allergy Clin. Immunol. 2014, 134, 1142–1152.e5. [Google Scholar] [CrossRef] [PubMed]
- Madzime, M.; Rossouw, T.M.; Theron, A.J.; Anderson, R.; Steel, H.C. Interactions of HIV and Antiretroviral Therapy With Neutrophils and Platelets. Front. Immunol. 2021, 12, 634386. [Google Scholar] [CrossRef] [PubMed]
- Gabali, A.M.; Anzinger, J.J.; Spear, G.T.; Thomas, L.L. Activation by inflammatory stimuli increases neutrophil binding of human immunodeficiency virus type 1 and subsequent infection of lymphocytes. J. Virol. 2004, 78, 10833–10836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hensley-McBain, T.; Wu, M.C.; Manuzak, J.A.; Cheu, R.K.; Gustin, A.; Driscoll, C.B.; Zevin, A.S.; Miller, C.J.; Coronado, E.; Smith, E.; et al. Increased mucosal neutrophil survival is associated with altered microbiota in HIV infection. PLoS Pathog. 2019, 15, e1007672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp, F.; Kupsch, S.; Schromm, A.B. Lipopolysaccharide-binding protein is bound and internalized by host cells and colocalizes with LPS in the cytoplasm: Implications for a role of LBP in intracellular LPS-signaling. Biochim. Biophys. Acta 2016, 1863, 660–672. [Google Scholar] [CrossRef]
- Pahwa, R.; Devaraj, S.; Jialal, I. The effect of the accessory proteins, soluble CD14 and lipopolysaccharide-binding protein on Toll-like receptor 4 activity in human monocytes and adipocytes. Int. J. Obes. 2016, 40, 907–911. [Google Scholar] [CrossRef]
- Sandler, N.G.; Douek, D.C. Microbial translocation in HIV infection: Causes, consequences and treatment opportunities. Nat. Rev. Microbiol. 2012, 10, 655–666. [Google Scholar] [CrossRef]
- Sandler, N.G.; Wand, H.; Roque, A.; Law, M.; Nason, M.C.; Nixon, D.E.; Pedersen, C.; Ruxrungtham, K.; Lewin, S.R.; Emery, S.; et al. Plasma levels of soluble CD14 independently predict mortality in HIV infection. J. Infect. Dis. 2011, 203, 780–790. [Google Scholar] [CrossRef]
- Zhao, T.; Su, Z.; Li, Y.; Zhang, X.; You, Q. Chitinase-3 like-protein-1 function and its role in diseases. Signal Transduct. Target. Ther. 2020, 5, 201. [Google Scholar] [CrossRef] [PubMed]
- Coffman, F.D. Chitinase 3-Like-1 (CHI3L1): A putative disease marker at the interface of proteomics and glycomics. Crit. Rev. Clin. Lab. Sci. 2008, 45, 531–562. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.; Kim, J.H.; Ha, E.K.; Jee, H.M.; Baek, H.S.; Han, M.Y.; Jeong, S.J. Serum YKL-40 Levels Are Associated with the Atherogenic Index of Plasma in Children. Mediat. Inflamm. 2020, 2020, 8713908. [Google Scholar] [CrossRef] [PubMed]
- Deutschmann, C.; Sowa, M.; Murugaiyan, J.; Roesler, U.; Röber, N.; Conrad, K.; Laass, M.W.; Bogdanos, D.; Sipeki, N.; Papp, M.; et al. Identification of Chitinase-3-Like Protein 1 as a Novel Neutrophil Antigenic Target in Crohn’s Disease. J. Crohn’s Colitis 2019, 13, 894–904. [Google Scholar] [CrossRef] [Green Version]
- He, C.H.; Lee, C.G.; Ma, B.; Kamle, S.; Choi, A.M.K.; Elias, J.A. N-Glycosylation Regulates Chitinase 3-like-1 and IL-13 Ligand Binding to IL-13 Receptor α2. Am. J. Respir. Cell Mol. Biol. 2020, 63, 386–395. [Google Scholar] [CrossRef]
- He, C.H.; Lee, C.G.; Dela Cruz, C.S.; Lee, C.M.; Zhou, Y.; Ahangari, F.; Ma, B.; Herzog, E.L.; Rosenberg, S.A.; Li, Y.; et al. Chitinase 3-like 1 regulates cellular and tissue responses via IL-13 receptor α2. Cell Rep. 2013, 4, 830–841. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.J.Z.M.; Schacker, T.W.; Reilly, C.S.; Haase, A.T. 131 The Role of Chitinase 3-Like-1 in the Pathological Deposition of Collagen in Lymphatic Tissue During HIV-1 Infection. JAIDS J. Acquir. Immune Defic. Syndr. 2011, 56, 53. [Google Scholar] [CrossRef]
- Bitnun, A.; Ransy, D.G.; Brophy, J.; Kakkar, F.; Hawkes, M.; Samson, L.; Annabi, B.; Pagliuzza, A.; Morand, J.A.; Sauve, L.; et al. Clinical Correlates of Human Immunodeficiency Virus-1 (HIV-1) DNA and Inducible HIV-1 RNA Reservoirs in Peripheral Blood in Children With Perinatally Acquired HIV-1 Infection With Sustained Virologic Suppression for at Least 5 Years. Clin. Infect. Dis. 2020, 70, 859–866. [Google Scholar] [CrossRef]
- Kastrup, J.; Johansen, J.S.; Winkel, P.; Hansen, J.F.; Hildebrandt, P.; Jensen, G.B.; Jespersen, C.M.; Kjøller, E.; Kolmos, H.J.; Lind, I.; et al. High serum YKL-40 concentration is associated with cardiovascular and all-cause mortality in patients with stable coronary artery disease. Eur. Heart J. 2009, 30, 1066–1072. [Google Scholar] [CrossRef] [Green Version]
- Rathcke, C.N.; Raymond, I.; Kistorp, C.; Hildebrandt, P.; Faber, J.; Vestergaard, H. Low grade inflammation as measured by levels of YKL-40: Association with an increased overall and cardiovascular mortality rate in an elderly population. Int. J. Cardiol. 2010, 143, 35–42. [Google Scholar] [CrossRef]
- McBride, J.A.; Striker, R. Imbalance in the game of T cells: What can the CD4/CD8 T-cell ratio tell us about HIV and health? PLoS Pathog. 2017, 13, e1006624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankowska-Konsur, A.; Łyko, M.; Rubas, K.; Nowicka-Suszko, D.; Maj, J.; Szepietowski, J.C. Chitinase-3-like Protein 1 (YKL-40): A New Biomarker of Inflammation in Pyoderma Gangrenosum. Acta Derm. Venereol. 2022, 102, adv00646. [Google Scholar] [CrossRef]
- Hattori, N.; Oda, S.; Sadahiro, T.; Nakamura, M.; Abe, R.; Shinozaki, K.; Nomura, F.; Tomonaga, T.; Matsushita, K.; Kodera, Y.; et al. YKL-40 identified by proteomic analysis as a biomarker of sepsis. Shock 2009, 32, 393–400. [Google Scholar] [CrossRef]
- De Lorenzo, R.; Sciorati, C.; Lorè, N.I.; Capobianco, A.; Tresoldi, C.; Cirillo, D.M.; Ciceri, F.; Rovere-Querini, P.; Manfredi, A.A. Chitinase-3-like protein-1 at hospital admission predicts COVID-19 outcome: A prospective cohort study. Sci. Rep. 2022, 12, 7606. [Google Scholar] [CrossRef] [PubMed]
- Conroy, A.L.; Hawkes, M.T.; Elphinstone, R.; Opoka, R.O.; Namasopo, S.; Miller, C.; John, C.C.; Kain, K.C. Chitinase-3-like 1 is a biomarker of acute kidney injury and mortality in paediatric severe malaria. Malar. J. 2018, 17, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A.; Kamrul-Hasan, A.; Kabir, M.R.; Das, S.; Zaki, K.; Al Mahtab, M. Evaluation of Chitinase 3-like 1 (CHI3L1) as a noninvasive biomarker of hepatic fibrosis in patients with Hepatitis B virus-related compensated chronic liver disease. J. Fam. Med. Prim. Care 2021, 10, 1694–1698. [Google Scholar]
- Seers, T.; Vassallo, P.; Pollock, K.; Thornhill, J.P.; Fidler, S.; Foster, C. CD4:CD8 ratio in children with perinatally acquired HIV-1 infection. HIV Med. 2018, 19, 668–672. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Villar, S.; Sainz, T.; Lee, S.A.; Hunt, P.W.; Sinclair, E.; Shacklett, B.L.; Ferre, A.L.; Hayes, T.L.; Somsouk, M.; Hsue, P.Y.; et al. HIV-infected individuals with low CD4/CD8 ratio despite effective antiretroviral therapy exhibit altered T cell subsets, heightened CD8+ T cell activation, and increased risk of non-AIDS morbidity and mortality. PLoS Pathog. 2014, 10, e1004078. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Villar, S.; Moreno, S.; Fuentes-Ferrer, M.; Sánchez-Marcos, C.; Avila, M.; Sainz, T.; de Villar, N.G.; Fernández-Cruz, A.; Estrada, V. The CD4:CD8 ratio is associated with markers of age-associated disease in virally suppressed HIV-infected patients with immunological recovery. HIV Med. 2014, 15, 40–49. [Google Scholar] [CrossRef]
- Sims, A.; Hadigan, C. Cardiovascular complications in children with HIV infection. Curr. HIV/AIDS Rep. 2011, 8, 209–214. [Google Scholar] [CrossRef]
- Merz, L.E.; Achebe, M. When non-Whiteness becomes a condition. Blood 2021, 137, 13–15. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, B.; Brown, J.; Ransy, D.G.; Brophy, J.; Kakkar, F.; Bitnun, A.; Samson, L.; Read, S.; Soudeyns, H.; Vaudry, W.; et al. Endothelial activation is associated with intestinal epithelial injury, systemic inflammation and treatment regimen in children living with vertically acquired HIV-1 infection. HIV Med. 2021, 22, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Saumoy, M.; Sánchez-Quesada, J.L.; Martínez, E.; Llibre, J.M.; Ribera, E.; Knobel, H.; Gatell, J.M.; Clotet, B.; Curran, A.; Curto, J.; et al. LDL subclasses and lipoprotein-phospholipase A2 activity in suppressed HIV-infected patients switching to raltegravir: Spiral substudy. Atherosclerosis 2012, 225, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Kjaergaard, A.D.; Helby, J.; Johansen, J.S.; Nordestgaard, B.G.; Bojesen, S.E. Elevated plasma YKL-40 and risk of infectious disease: A prospective study of 94665 individuals from the general population. Clin. Microbiol. Infect. 2020, 26, e1–e1411. [Google Scholar] [CrossRef]
- Glesby, M. Cardiovascular Complications of HIV Infection. Top. Antivir. Med. 2016, 24, 127–131. [Google Scholar]
- Kamle, S.; Ma, B.; He, C.H.; Akosman, B.; Zhou, Y.; Lee, C.M.; El-Deiry, W.S.; Huntington, K.; Liang, O.; Machan, J.T.; et al. Chitinase 3-like-1 is a therapeutic target that mediates the effects of aging in COVID-19. JCI Insight 2021, 6, e148749. [Google Scholar] [CrossRef] [PubMed]
- Park, K.R.; Park, J.I.; Lee, S.; Yoo, K.; Kweon, G.R.; Kwon, I.K.; Yun, H.M.; Hong, J.T. Chi3L1 is a therapeutic target in bone metabolism and a potential clinical marker in patients with osteoporosis. Pharmacol. Res. 2022, 184, 106423. [Google Scholar] [CrossRef] [PubMed]
- Libreros, S.; Garcia-Areas, R.; Shibata, Y.; Carrio, R.; Torroella-Kouri, M.; Iragavarapu-Charyulu, V. Induction of proinflammatory mediators by CHI3L1 is reduced by chitin treatment: Decreased tumor metastasis in a breast cancer model. Int. J. Cancer 2012, 131, 377–386. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Overall Cohort (N = 105) | Low CHI3L1 1 (N = 52) | High CHI3L1 1 (N = 53) | p-Value |
|---|---|---|---|---|
| Demographics | ||||
| Age (yr), median (IQR) | 13 (8.6–17) | 13 (8.5–16) | 14 (8.6–18) | 0.28 |
| <8 years, n (%) | 22 (21) | 10 (19) | 12 (23) | 0.70 |
| 8 to <12 years, n (%) | 27 (26) | 13 (25) | 14 (26) | |
| 12 to <16 years, n (%) | 20 (19) | 9 (17) | 11 (21) | |
| ≥16 years, n (%) | 36 (34) | 21 (40) | 15 (28) | |
| Female sex, n (%) | 62 (59) | 33 (63) | 29 (55) | 0.48 |
| Ethnicity (mother), n (%) | 0.16 | |||
| African, Caribbean, or Black | 67 (64) | 38 (73) | 29 (55) | |
| White | 12 (11) | 8 (15) | 4 (7.5) | |
| First Nations | 7 (6.7) | 2 (3.8) | 5 (9.4) | |
| Other | 16 (15) | 4 (7.7) | 12 (23) | |
| Unknown | 3 (2.9) | 0 (0) | 3 (5.7) | |
| HIV Clade, n (%) | 0.21 | |||
| Clade A | 4 (3.8) | 0 (0) | 4 (7.5) | |
| Clade B | 32 (30) | 18 (35) | 14 (26) | |
| Clade C | 21 (20) | 10 (19) | 11 (21) | |
| Other | 22 (21) | 11 (21) | 11 (21) | |
| Unknown | 26 (25) | 13 (25) | 13 (25) | |
| Historical HIV control, median (IQR) | ||||
| Age at initiation of any ARVs (yr) | 2.1 (0.38–5.5) | 3.4 (0.84–5.5) | 1.7 (0.36–5.3) | 0.32 |
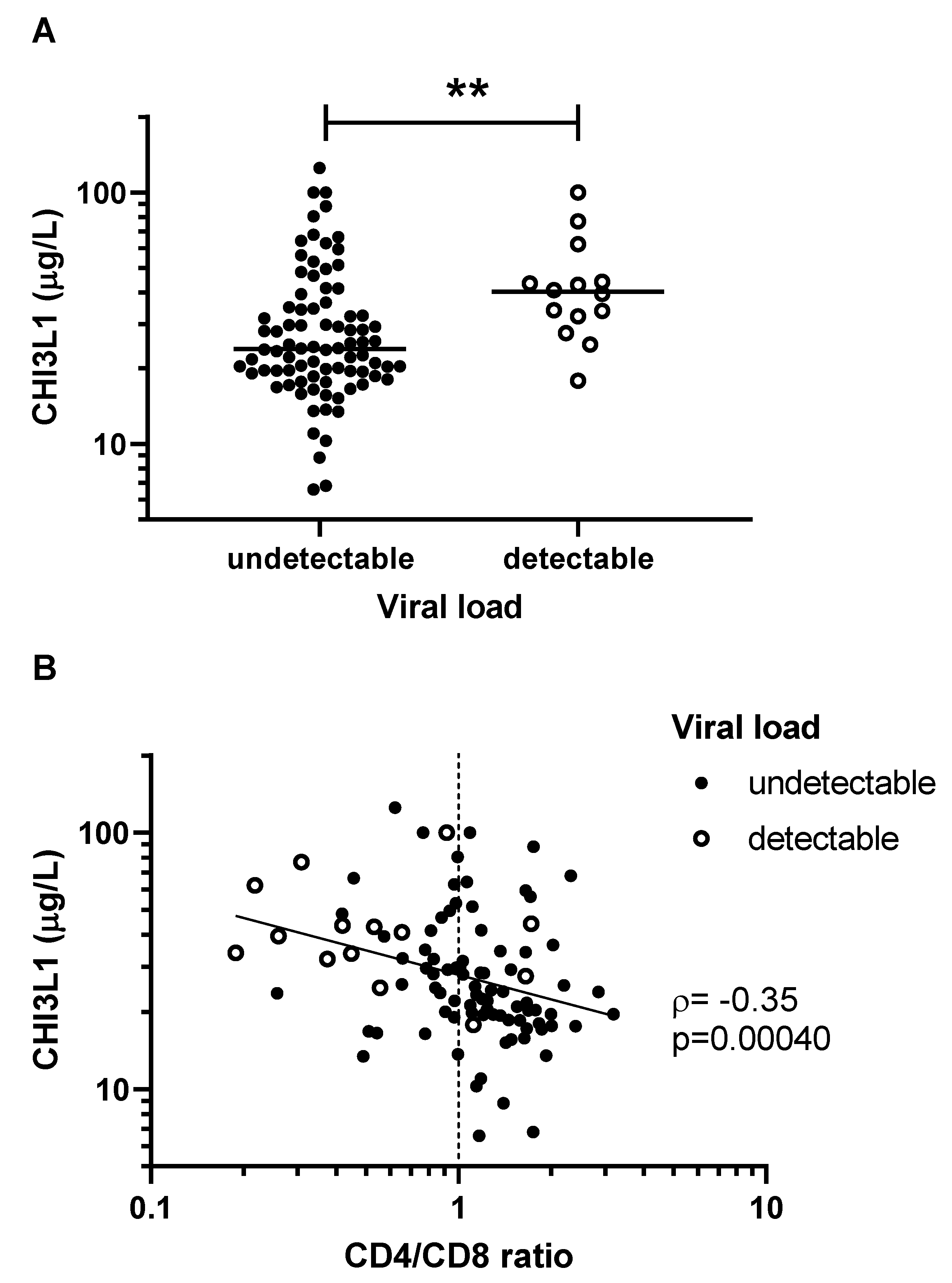
| Undetectable viral load, n (%) | 84 (86) | 46 (96) | 38 (76) | 0.0026 |
| Proportion SVS, n (%) | 81 (77) | 45 (87) | 36 (68) | 0.041 |
| Age at SVS (yr) 2 | 6.6 (3–11) | 6.6 (3.8–12) | 6.6 (2.4–8.9) | 0.27 |
| Duration of SVS (yr) 2 | 6.0 (3–8.4) | 4.9 (2.4–7.3) | 6.1 (3.7–9.8) | 0.11 |
| Current cART regimen, n (%) | 0.096 | |||
| NNRTI-based | 41 (39) | 22 (42) | 19 (36) | |
| Protease-inhibitor-based | 23 (22) | 11 (21) | 12 (23) | |
| Integrase inhibitor-based | 17 (16) | 12 (23) | 5 (9.4) | |
| No ARVs | 10 (9.5) | 2 (3.8) | 8 (15) | |
| Other 3 | 14 (13) | 5 (9.6) | 9 (17) | |
| Immunologic variables, median (IQR) | ||||
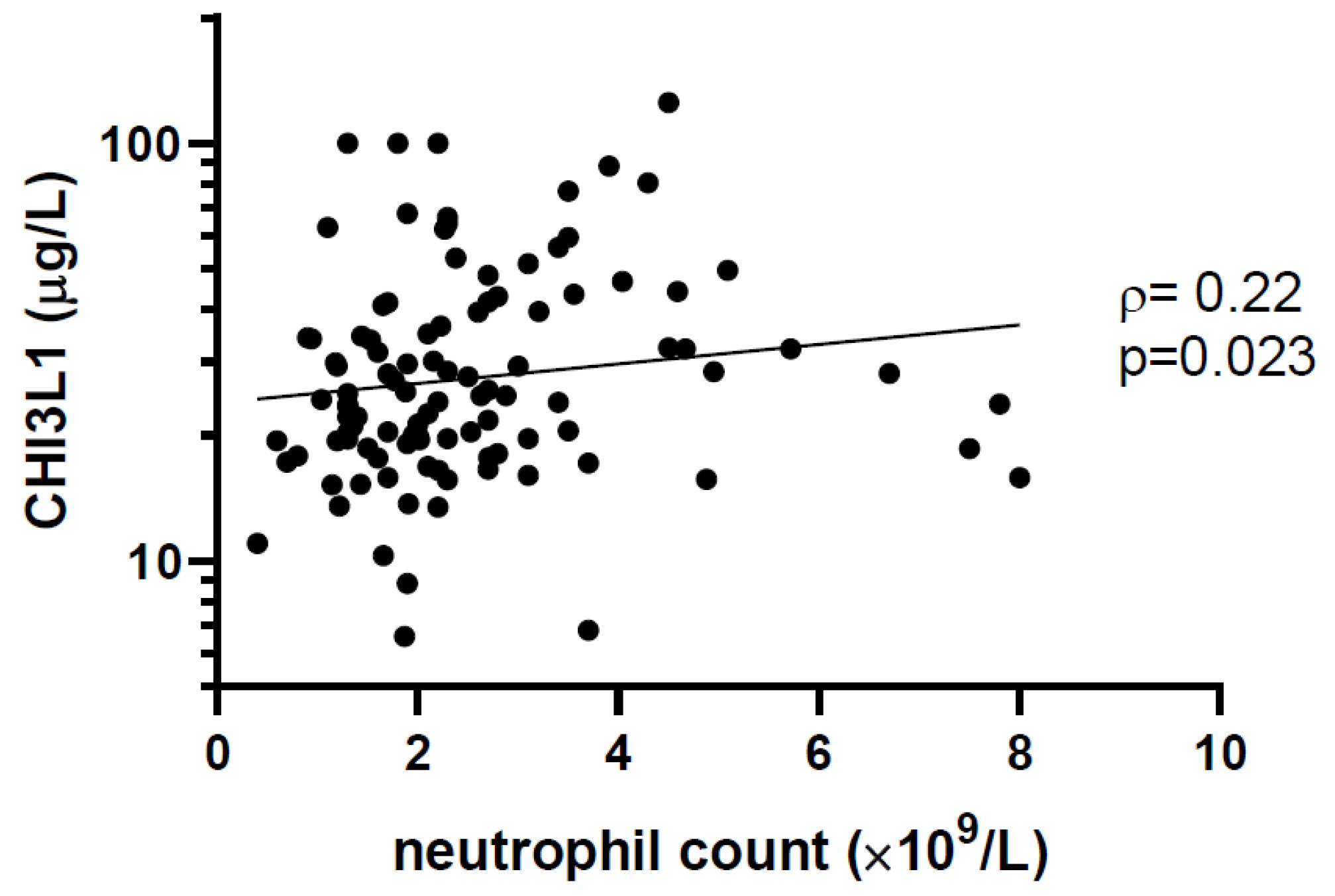
| Neutrophil count (×109/L) | 2.2 (1.6–3.1) | 2.0 (1.3–2.7) | 2.3 (1.8–3.5) | 0.041 |
| Neutrophil percent | 41 (33–52) | 38 (32–50) | 43 (36–57) | 0.11 |
| CD4+ T-cell count (×109/L) | 770 (550–1100) | 830 (590–1100) | 690 (510–990) | 0.13 |
| CD4+ T-cell percent | 37 (30–42) | 40 (35–43) | 33 (26–40) | 0.0011 |
| CD4 lifetime nadir (×109/L) | 440 (280–570) | 440 (290–570) | 440 (250–610) | 0.79 |
| CD4 lifetime nadir percent | 20 (14–29) | 23 (15–29) | 19 (13–26) | 0.12 |
| CD8+ T-cell count (×109/L) | 740 (520–1000) | 620 (460–860) | 810 (600–1100) | 0.0071 |
| CD8+ T-cell percent | 33 (27–38) | 30 (25–35) | 36 (29–44) | 0.00039 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernard, I.; Ransy, D.G.; Brophy, J.; Kakkar, F.; Bitnun, A.; Samson, L.; Read, S.; Soudeyns, H.; Hawkes, M.T.; EPIC4 Study Group. Chitinase-3-like Protein 1 Is Associated with Poor Virologic Control and Immune Activation in Children Living with HIV. Viruses 2022, 14, 2602. https://doi.org/10.3390/v14122602
Bernard I, Ransy DG, Brophy J, Kakkar F, Bitnun A, Samson L, Read S, Soudeyns H, Hawkes MT, EPIC4 Study Group. Chitinase-3-like Protein 1 Is Associated with Poor Virologic Control and Immune Activation in Children Living with HIV. Viruses. 2022; 14(12):2602. https://doi.org/10.3390/v14122602
Chicago/Turabian StyleBernard, Isabelle, Doris G. Ransy, Jason Brophy, Fatima Kakkar, Ari Bitnun, Lindy Samson, Stanley Read, Hugo Soudeyns, Michael T. Hawkes, and EPIC4 Study Group. 2022. "Chitinase-3-like Protein 1 Is Associated with Poor Virologic Control and Immune Activation in Children Living with HIV" Viruses 14, no. 12: 2602. https://doi.org/10.3390/v14122602
APA StyleBernard, I., Ransy, D. G., Brophy, J., Kakkar, F., Bitnun, A., Samson, L., Read, S., Soudeyns, H., Hawkes, M. T., & EPIC4 Study Group. (2022). Chitinase-3-like Protein 1 Is Associated with Poor Virologic Control and Immune Activation in Children Living with HIV. Viruses, 14(12), 2602. https://doi.org/10.3390/v14122602