

The Optimized γ-Globin Lentiviral Vector GGHI-mB-3D Leads to Nearly Therapeutic HbF Levels In Vitro in CD34+ Cells from Sickle Cell Disease Patients

 ,
, 
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Production and Titration
2.2. Human CD34+ Stem Cell Isolation and Transduction: Sample Collection and Processing
2.3. Reversed-Phase High-Performance Liquid Chromatography (RP-HPLC) Analysis for Globin Chain Aγ/α Ratio Quantitation
2.4. Hemoglobin Electrophoresis and Cation Exchange HPLC (CE-HPLC) for HbF and HbS Quantitation
2.5. RNA Analysis and Measurement of γ-Globin Transcript Levels Using Quantitative Real-Time PCR
2.6. Flow Cytometry
2.7. Determination of Vector Copy Number and Transduction Efficiency
2.8. Statistical Analysis
3. Results
3.1. GGHI and GGHI-mB-3D LVs Exhibit High Titers
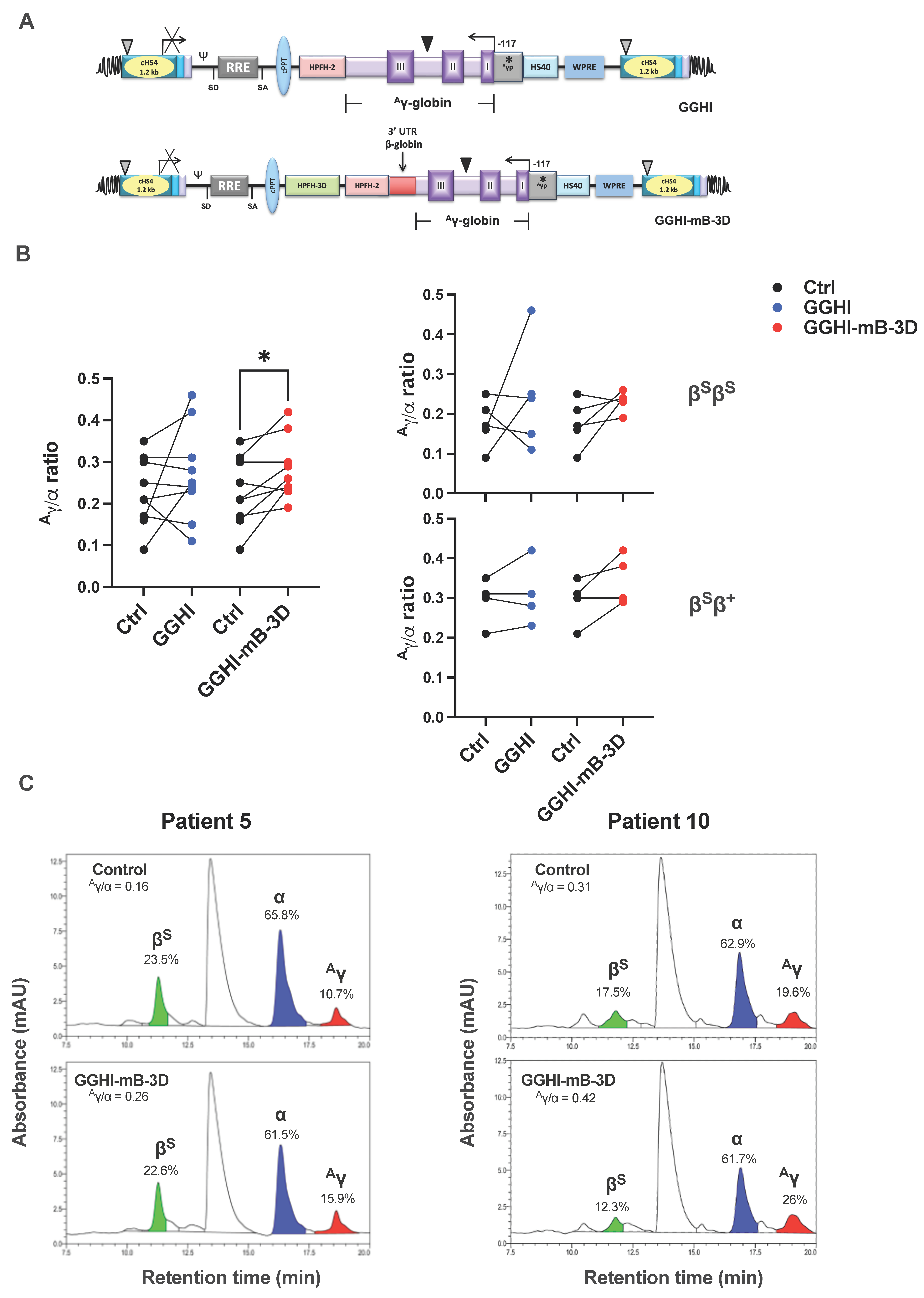
3.2. Increased Aγ/α Chain Ratio in SCD CD34+ Cells following Transduction with GGHI-mB-3D Lentiviral Vector
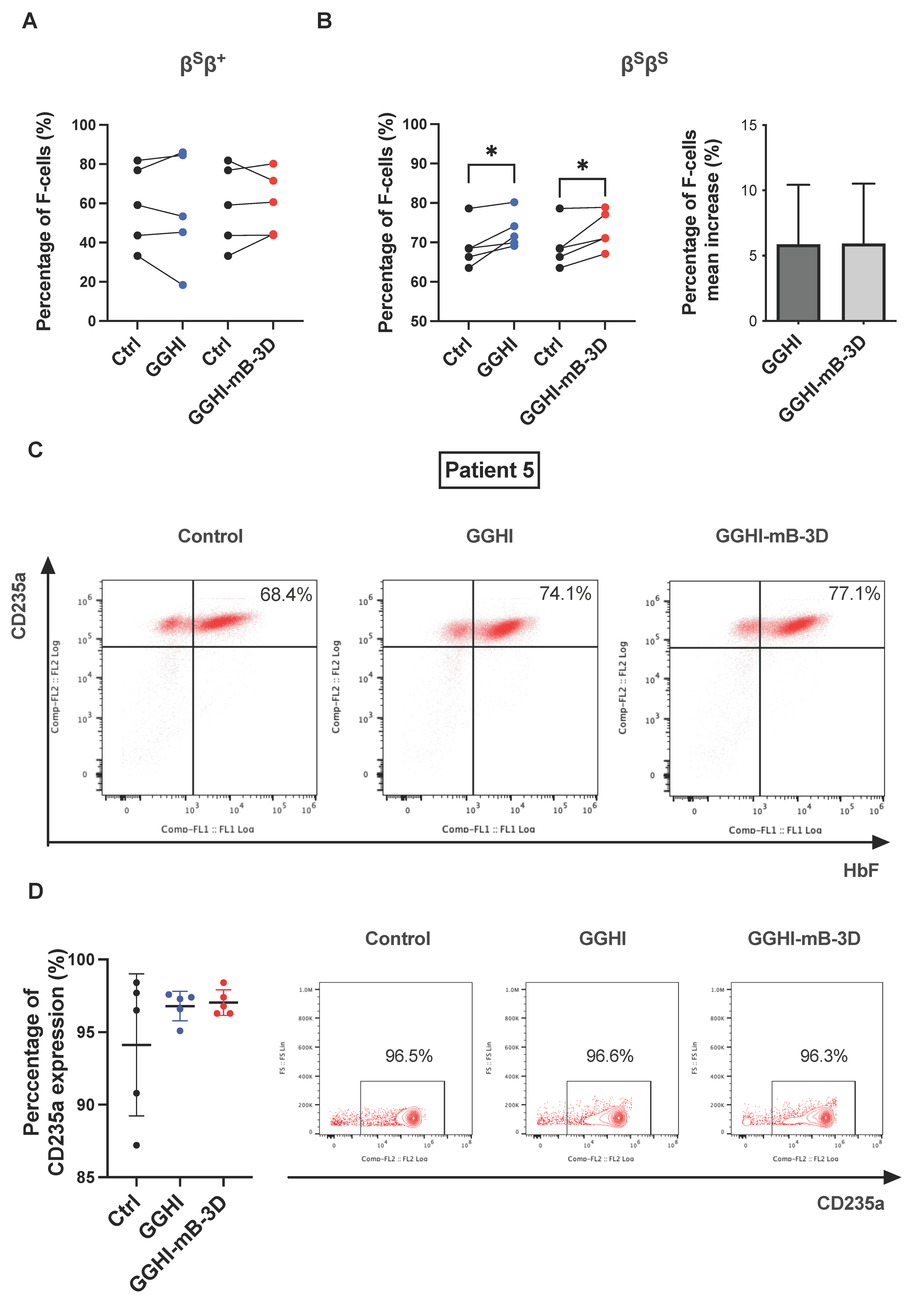
3.3. Increased Percentage of F-Cells following Transduction with GGHI-mB-3D Lentiviral Vector
3.4. Improvement in the SCD CD34+ Cell Phenotype In Vitro following Transduction with GGHI-mB-3D Lentiviral Vector
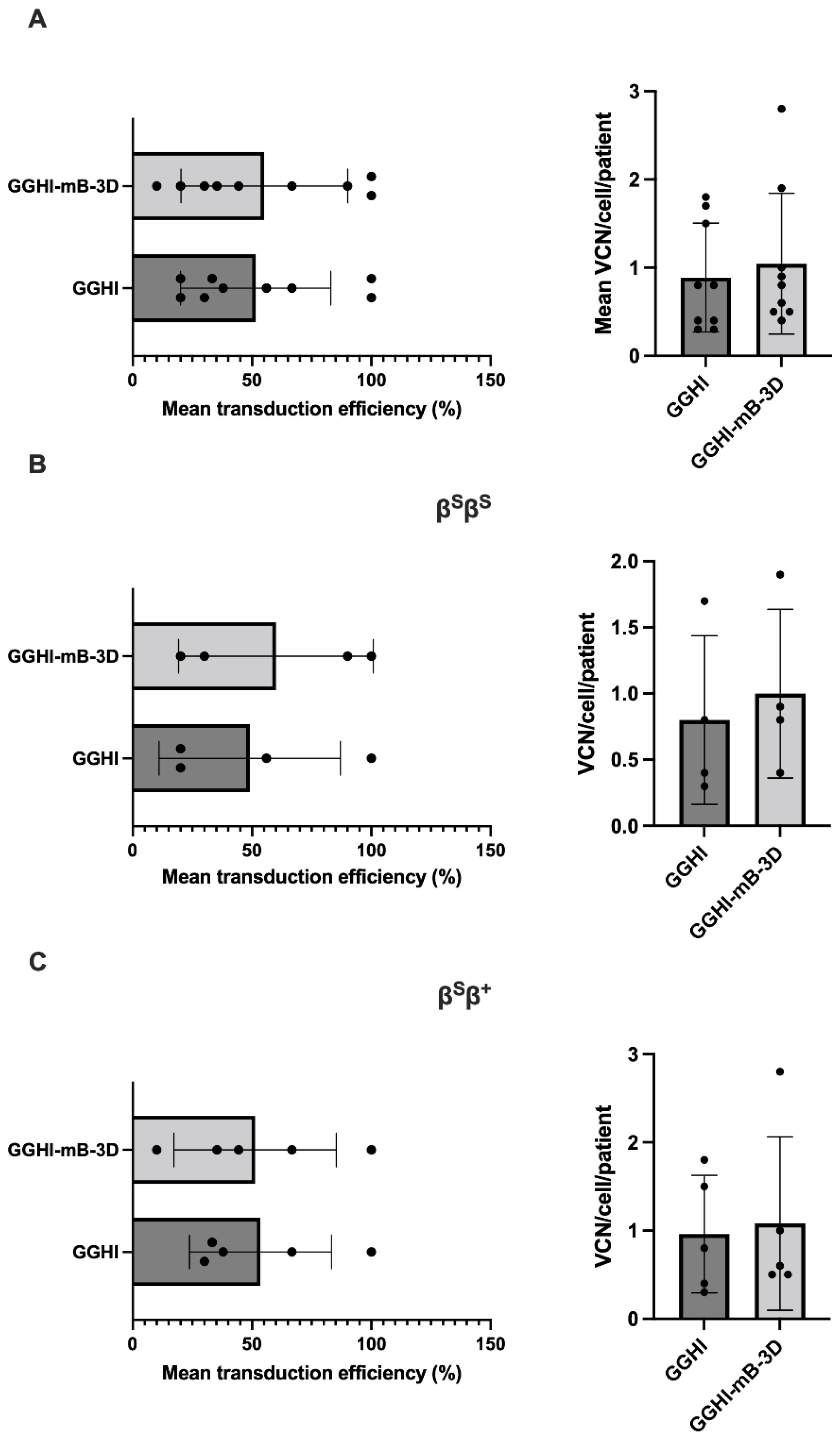
3.5. Gene Transfer Efficiency and Vector Copy Number in Transduced BFUe from SCD Patients
3.6. γ-Globin mRNA Analysis Using Quantitative Real-Time PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ingram, V.M. Gene Mutations in Human Hæmoglobin: The Chemical Difference Between Normal and Sickle Cell Hæmoglobin. Nature 1957, 180, 326–328. [Google Scholar] [CrossRef] [PubMed]
- Driss, A.; Asare, K.O.; Hibbert, J.M.; Gee, B.E.; Adamkiewicz, T.V.; Stiles, J.K. Sickle Cell Disease in the Post Genomic Era: A Monogenic Disease with a Polygenic Phenotype. Genom. Insights 2009, 2009, 23–48. [Google Scholar] [CrossRef] [Green Version]
- Milunsky, A. Sickle Cell Disease. N. Engl. J. Med. 2017, 377, 302–305. [Google Scholar] [CrossRef]
- Powars, D.R.; Weiss, J.N.; Chan, L.S.; Schroeder, W.A. Is there a threshold level of fetal hemoglobin that ameliorates morbidity in sickle cell anemia? Blood 1984, 63, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Platt, O.S.; Brambilla, D.J.; Rosse, W.F.; Milner, P.F.; Castro, O.; Steinberg, M.H.; Klug, P.P. Mortality in sickle cell disease—Life expectancy and risk factors for early death. N. Engl. J. Med. 1994, 330, 1639–1644. [Google Scholar] [CrossRef] [PubMed]
- Koshy, M.; Entsuah, R.; Koranda, A.; Kraus, A.P.; Johnson, R.; Bellvue, R.; Flournoy-Gill, Z.; Levy, P. Leg ulcers in patients with sickle cell disease. Blood 1989, 74, 1403–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolan, V.G.; Adewoye, A.; Baldwin, C.; Wang, L.; Ma, Q.; Wyszynski, D.F.; Farrell, J.J.; Sebastiani, P.; Farrer, L.A.; Steinberg, M.H. Sickle cell leg ulcers: Associations with haemolysis and SNPs in Klotho, TEK and genes of the TGF-beta/BMP pathway. Br. J. Haematol. 2006, 133, 570–578. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, M.H.; Forget, B.G.; Higgs, D.R.; Weatherall, D.J. Disorders of Hemoglobin: Genetics, Pathophysiology and Clinical Management; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Bunn, H.F.; Forget, B.G. Hemoglobin-Molecular, Genetic and Clinical Aspects; W. B. Saunders Co.,Ltd.: Philadelphia, PA, USA, 1986; Volume VII. [Google Scholar]
- Steinberg, M.H. Targeting fetal hemoglobin expression to treat β hemoglobinopathies. Expert Opin. Ther. Targets 2022, 26, 347–359. [Google Scholar] [CrossRef]
- Ley, T.J.; DeSimone, J.; Anagnou, N.P.; Keller, G.H.; Humphries, R.K.; Turner, P.H.; Young, N.S.; Heller, P.; Nienhuis, A.W. 5-Azacytidine Selectively Increases γ-Globin Synthesis in a Patient with β+Thalassemia. N. Engl. J. Med. 1982, 307, 1469–1475. [Google Scholar] [CrossRef]
- Charache, S.; Dover, G.; Smith, K.; Talbot, C.C., Jr.; Moyer, M.; Boyer, S. Treatment of sickle cell anemia with 5-azacytidine results in increased fetal hemoglobin production and is associated with nonrandom hypomethylation of DNA around the gamma-delta-beta-globin gene complex. Proc. Natl. Acad. Sci. USA 1983, 80, 4842–4846. [Google Scholar] [CrossRef]
- Charache, S.; Terrin, M.L.; Moore, R.D.; Dover, G.J.; Barton, F.B.; Eckert, S.V.; McMahon, R.P.; Bonds, D.R. Effect of Hydroxyurea on the Frequency of Painful Crises in Sickle Cell Anemia. N. Engl. J. Med. 1995, 332, 1317–1322. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, M.H.; Barton, F.; Castro, O.; Pegelow, C.H.; Ballas, S.K.; Kutlar, A.; Orringer, E.; Bellevue, R.; Olivieri, N.; Eckman, J.; et al. Effect of Hydroxyurea on Mortality and Morbidity in Adult Sickle Cell Anemia. JAMA 2003, 289, 1645–1651. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, M.H.; McCarthy, W.F.; Castro, O.; Ballas, S.K.; Armstrong, F.D.; Smith, W.; Ataga, K.; Swerdlow, P.; Kutlar, A.; DeCastro, L.; et al. The risks and benefits of long-term use of hydroxyurea in sickle cell anemia: A 17.5 year follow-up. Am. J. Hematol. 2010, 85, 403–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivieri, N.F.; Brittenham, G.M. Management of the Thalassemias. Cold Spring Harb. Perspect. Med. 2013, 3, a011767. [Google Scholar] [CrossRef]
- Ribeil, J.-A.; Hacein-Bey-Abina, S.; Payen, E.; Magnani, A.; Semeraro, M.; Magrin, E.; Caccavelli, L.; Neven, B.; Bourget, P.; El Nemer, W.; et al. Gene Therapy in a Patient with Sickle Cell Disease. N. Engl. J. Med. 2017, 376, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Pestina, T.I.; Hargrove, P.W.; Jay, D.; Gray, J.T.; Boyd, K.M.; Persons, D.A. Correction of murine sickle cell disease using gamma-globin lentiviral vectors to mediate high-level expression of fetal hemoglobin. Mol. Ther. 2009, 17, 245–252. [Google Scholar] [CrossRef]
- Levasseur, D.N.; Ryan, T.M.; Pawlik, K.M.; Townes, T.M. Correction of a mouse model of sickle cell disease: Lentiviral/antisickling beta-globin gene transduction of unmobilized, purified hematopoietic stem cells. Blood 2003, 102, 4312–4319. [Google Scholar] [CrossRef]
- Perumbeti, A.; Higashimoto, T.; Urbinati, F.; Franco, R.; Meiselman, H.J.; Witte, D.; Malik, P. A novel human gamma-globin gene vector for genetic correction of sickle cell anemia in a humanized sickle mouse model: Critical determinants for successful correction. Blood 2009, 114, 1174–1185. [Google Scholar] [CrossRef] [Green Version]
- Tisdale, J.F.; Thein, S.L.; Eaton, W.A. Treating sickle cell anemia. Science 2020, 367, 1198–1199. [Google Scholar] [CrossRef]
- Urbinati, F.; Hargrove, P.W.; Geiger, S.; Romero, Z.; Wherley, J.; Kaufman, M.L.; Hollis, R.P.; Chambers, C.B.; Persons, D.A.; Kohn, D.B.; et al. Potentially therapeutic levels of anti-sickling globin gene expression following lentivirus-mediated gene transfer in sickle cell disease bone marrow CD34+ cells. Exp. Hematol. 2015, 43, 346–351. [Google Scholar] [CrossRef]
- Weber, L.; Poletti, V.; Magrin, E.; Antoniani, C.; Martin, S.; Bayard, C.; Sadek, H.; Felix, T.; Meneghini, V.; Antoniou, M.N.; et al. An Optimized Lentiviral Vector Efficiently Corrects the Human Sickle Cell Disease Phenotype. Mol. Ther. Methods Clin. Dev. 2018, 10, 268–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breda, L.; Casu, C.; Gardenghi, S.; Bianchi, N.; Cartegni, L.; Narla, M.; Yazdanbakhsh, K.; Musso, M.; Manwani, D.; Little, J.; et al. Therapeutic hemoglobin levels after gene transfer in beta-thalassemia mice and in hematopoietic cells of beta-thalassemia and sickle cells disease patients. PLoS ONE 2012, 7, e32345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samakoglu, S.; Lisowski, L.; Budak-Alpdogan, T.; Usachenko, Y.; Acuto, S.; Di Marzo, R.; Maggio, A.; Zhu, P.; Tisdale, J.F.; Riviere, I.; et al. A genetic strategy to treat sickle cell anemia by coregulating globin transgene expression and RNA interference. Nat. Biotechnol. 2006, 24, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Peng, C.; Sankaran, V.G.; Shao, Z.; Esrick, E.B.; Chong, B.G.; Ippolito, G.C.; Fujiwara, Y.; Ebert, B.L.; Tucker, P.W.; et al. Correction of sickle cell disease in adult mice by interference with fetal hemoglobin silencing. Science 2011, 334, 993–996. [Google Scholar] [CrossRef] [Green Version]
- Tallack, M.R.; Perkins, A.C. Three fingers on the switch: Kruppel-like factor 1 regulation of gamma-globin to beta-globin gene switching. Curr. Opin. Hematol. 2013, 20, 193–200. [Google Scholar] [CrossRef]
- Sankaran, V.G.; Xu, J.; Ragoczy, T.; Ippolito, G.C.; Walkley, C.R.; Maika, S.D.; Fujiwara, Y.; Ito, M.; Groudine, M.; Bender, M.A.; et al. Developmental and species-divergent globin switching are driven by BCL11A. Nature 2009, 460, 1093–1097. [Google Scholar] [CrossRef] [Green Version]
- Sankaran, V.G.; Menne, T.F.; Scepanovic, D.; Vergilio, J.A.; Ji, P.; Kim, J.; Thiru, P.; Orkin, S.H.; Lander, E.S.; Lodish, H.F. MicroRNA-15a and -16-1 act via MYB to elevate fetal hemoglobin expression in human trisomy 13. Proc. Natl. Acad. Sci. USA 2011, 108, 1519–1524. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.A.; Esrick, E. Investigational curative gene therapy approaches to sickle cell disease. Blood Adv. 2021, 5, 5452. [Google Scholar] [CrossRef]
- Magis, W.; DeWitt, M.A.; Wyman, S.K.; Vu, J.T.; Heo, S.J.; Shao, S.J.; Hennig, F.; Romero, Z.G.; Campo-Fernandez, B.; Said, S.; et al. High-level correction of the sickle mutation is amplified in vivo during erythroid differentiation. iScience 2022, 25, 104374. [Google Scholar] [CrossRef]
- Papanikolaou, E.; Georgomanoli, M.; Stamateris, E.; Panetsos, F.; Karagiorga, M.; Tsaftaridis, P.; Graphakos, S.; Anagnou, N.P. The new self-inactivating lentiviral vector for thalassemia gene therapy combining two HPFH activating elements corrects human thalassemic hematopoietic stem cells. Hum. Gene Ther. 2012, 23, 15–31. [Google Scholar] [CrossRef]
- Katsantoni, E.Z.; Langeveld, A.; Wai, A.W.; Drabek, D.; Grosveld, F.; Anagnou, N.P.; Strouboulis, J. Persistent gamma-globin expression in adult transgenic mice is mediated by HPFH-2, HPFH-3, and HPFH-6 breakpoint sequences. Blood 2003, 102, 3412–3419. [Google Scholar] [CrossRef] [PubMed]
- Fragkos, M.; Anagnou, N.P.; Tubb, J.; Emery, D.W. Use of the hereditary persistence of fetal hemoglobin 2 enhancer to increase the expression of oncoretrovirus vectors for human gamma-globin. Gene Ther. 2005, 12, 1591–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Emery, D.W.; Fernandez, M.; Han, H.; Stamatoyannopoulos, G. Development of viral vectors for gene therapy of beta-chain hemoglobinopathies: Optimization of a gamma-globin gene expression cassette. Blood 1999, 93, 2208–2216. [Google Scholar] [CrossRef] [PubMed]
- Cavazzana-Calvo, M.; Payen, E.; Negre, O.; Wang, G.; Hehir, K.; Fusil, F.; Down, J.; Denaro, M.; Brady, T.; Westerman, K.; et al. Transfusion independence and HMGA2 activation after gene therapy of human beta-thalassaemia. Nature 2010, 467, 318–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulad, F.; Maggio, A.; Wang, X.; Moi, P.; Acuto, S.; Kogel, F.; Takpradit, C.; Prockop, S.; Mansilla-Soto, J.; Cabriolu, A.; et al. Lentiviral globin gene therapy with reduced-intensity conditioning in adults with beta-thalassemia: A phase 1 trial. Nat. Med. 2022, 28, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Drakopoulou, E.; Georgomanoli, M.; Lederer, C.W.; Kleanthous, M.; Costa, C.; Bernadin, O.; Cosset, F.L.; Voskaridou, E.; Verhoeyen, E.; Papanikolaou, E.; et al. A Novel BaEVRless-Pseudotyped gamma-Globin Lentiviral Vector Drives High and Stable Fetal Hemoglobin Expression and Improves Thalassemic Erythropoiesis In Vitro. Hum. Gene Ther. 2019, 30, 601–617. [Google Scholar] [CrossRef] [PubMed]
- Elder, J.T.; Forrester, W.C.; Thompson, C.; Mager, D.; Henthorn, P.; Peretz, M.; Papayannopoulou, T.; Groudine, M. Translocation of an erythroid-specific hypersensitive site in deletion-type hereditary persistence of fetal hemoglobin. Mol. Cell. Biol. 1990, 10, 1382–1389. [Google Scholar] [CrossRef]
- Tuan, D.; Feingold, E.; Newman, M.; Weissman, S.M.; Forget, B.G. Different 3’ end points of deletions causing delta beta-thalassemia and hereditary persistence of fetal hemoglobin: Implications for the control of gamma-globin gene expression in man. Proc. Natl. Acad. Sci. USA 1983, 80, 6937–6941. [Google Scholar] [CrossRef] [Green Version]
- Frecha, C.; Costa, C.; Negre, D.; Amirache, F.; Trono, D.; Rio, P.; Bueren, J.; Cosset, F.L.; Verhoeyen, E. A novel lentiviral vector targets gene transfer into human hematopoietic stem cells in marrow from patients with bone marrow failure syndrome and in vivo in humanized mice. Blood 2012, 119, 1139–1150. [Google Scholar] [CrossRef] [Green Version]
- Girard-Gagnepain, A.; Amirache, F.; Costa, C.; Levy, C.; Frecha, C.; Fusil, F.; Negre, D.; Lavillette, D.; Cosset, F.L.; Verhoeyen, E. Baboon envelope pseudotyped LVs outperform VSV-G-LVs for gene transfer into early-cytokine-stimulated and resting HSCs. Blood 2014, 124, 1221–1231. [Google Scholar] [CrossRef]
- Dull, T.; Zufferey, R.; Kelly, M.; Mandel, R.J.; Nguyen, M.; Trono, D.; Naldini, L. A third-generation lentivirus vector with a conditional packaging system. J. Virol. 1998, 72, 8463–8471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zufferey, R.; Dull, T.; Mandel, R.J.; Bukovsky, A.; Quiroz, D.; Naldini, L.; Trono, D. Self-inactivating lentivirus vector for safe and efficient in vivo gene delivery. J. Virol. 1998, 72, 9873–9880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanikolaou, E.; Kontostathi, G.; Drakopoulou, E.; Georgomanoli, M.; Stamateris, E.; Vougas, K.; Vlahou, A.; Maloy, A.; Ware, M.; Anagnou, N.P. Characterization and comparative performance of lentiviral vector preparations concentrated by either one-step ultrafiltration or ultracentrifugation. Virus Res. 2013, 175, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Loucari, C.C.; Patsali, P.; van Dijk, T.B.; Stephanou, C.; Papasavva, P.; Zanti, M.; Kurita, R.; Nakamura, Y.; Christou, S.; Sitarou, M.; et al. Rapid and Sensitive Assessment of Globin Chains for Gene and Cell Therapy of Hemoglobinopathies. Hum. Gene Ther. Methods 2018, 29, 60–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Lisowski, L.; Sadelain, M. Locus control region elements HS1 and HS4 enhance the therapeutic efficacy of globin gene transfer in beta-thalassemic mice. Blood 2007, 110, 4175–4178. [Google Scholar] [CrossRef] [PubMed]
- Fujita, A.; Uchida, N.; Haro-Mora, J.J.; Winkler, T.; Tisdale, J. beta-Globin-Expressing Definitive Erythroid Progenitor Cells Generated from Embryonic and Induced Pluripotent Stem Cell-Derived Sacs. Stem Cells 2016, 34, 1541–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahajan, M.C.; Karmakar, S.; Newburger, P.E.; Krause, D.S.; Weissman, S.M. Dynamics of alpha-globin locus chromatin structure and gene expression during erythroid differentiation of human CD34(+) cells in culture. Exp. Hematol. 2009, 37, 1143–1156.e3. [Google Scholar] [CrossRef] [Green Version]
- Kanter, J.; Walters, M.C.; Krishnamurti, L.; Mapara, M.Y.; Kwiatkowski, J.L.; Rifkin-Zenenberg, S.; Aygun, B.; Kasow, K.A.; Pierciey, F.J., Jr.; Bonner, M.; et al. Biologic and Clinical Efficacy of LentiGlobin for Sickle Cell Disease. N. Engl. J. Med. 2022, 386, 617–628. [Google Scholar] [CrossRef]
- Cabriolu, A.; Odak, A.; Zamparo, L.; Yuan, H.; Leslie, C.S.; Sadelain, M. Globin vector regulatory elements are active in early hematopoietic progenitor cells. Mol. Ther. 2022, 30, 2199–2209. [Google Scholar] [CrossRef]
- Goyal, S.; Tisdale, J.; Schmidt, M.; Kanter, J.; Jaroscak, J.; Whitney, D.; Bitter, H.; Gregory, P.D.; Parsons, G.; Foos, M.; et al. Acute Myeloid Leukemia Case after Gene Therapy for Sickle Cell Disease. N. Engl. J. Med. 2022, 386, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Lettre, G.; Sankaran, V.G.; Bezerra, M.A.; Araujo, A.S.; Uda, M.; Sanna, S.; Cao, A.; Schlessinger, D.; Costa, F.F.; Hirschhorn, J.N.; et al. DNA polymorphisms at the BCL11A, HBS1L-MYB, and beta-globin loci associate with fetal hemoglobin levels and pain crises in sickle cell disease. Proc. Natl. Acad. Sci. USA 2008, 105, 11869–11874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galarneau, G.; Palmer, C.D.; Sankaran, V.G.; Orkin, S.H.; Hirschhorn, J.N.; Lettre, G. Fine-mapping at three loci known to affect fetal hemoglobin levels explains additional genetic variation. Nat. Genet. 2010, 42, 1049–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzel, S.; Thein, S.L. Genetic Modifiers of Fetal Haemoglobin in Sickle Cell Disease. Mol. Diagn. Ther. 2019, 23, 235–244. [Google Scholar] [CrossRef]
- Sales, R.R.; Nogueira, B.L.; Tosatti, J.A.G.; Gomes, K.B.; Luizon, M.R. Do Genetic Polymorphisms Affect Fetal Hemoglobin (HbF) Levels in Patients With Sickle Cell Anemia Treated With Hydroxyurea? A Systematic Review and Pathway Analysis. Front. Pharmacol. 2021, 12, 779497. [Google Scholar] [CrossRef]
- Iolascon, A.; Rivella, S.; Anagnou, N.P.; Camaschella, C.; Swinkels, D.; Muckenthaler, M.U.; Porto, G.; Barcellini, W.; Andolfo, I.; Risitano, A.M.; et al. The EHA Research Roadmap: Anemias. Hemasphere 2021, 5, e607. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
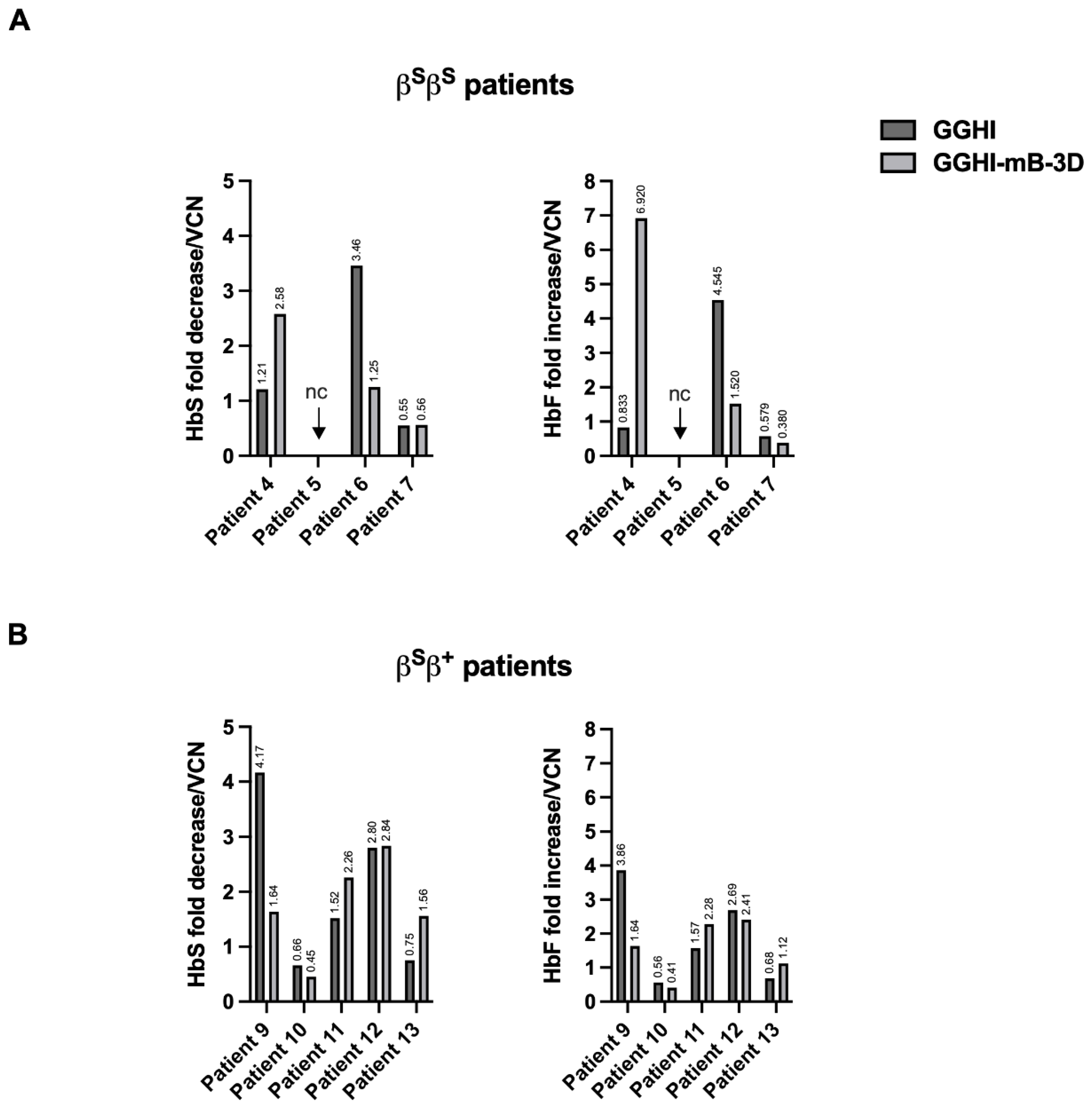{kind=link}
| Sample | Genotype | Mutation | Source | F-Cells (FACS) | Aγ/α Ratio (RP-HPLC) | HbS Decrease (%) | TSD Efficiency (%) | Mean VCN/Cell | Relative Fold Difference of γ-mRNA Transcripts | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ctrl | GGHI | GGHI-mB-3D | Ctrl | GGHI | GGHI-mB-3D | GGHI | GGHI- mB-3D | GGHI | GGHI- mB-3D | GGHI | GGHI- mB-3D | GGHI | GGHI- mB-3D | ||||
| #4 | βSβS | HbS/HbS | PB | 66.3 | 69.1 | 71.1 | 0.21 | 0.11 | 0.24 | 0 | 3.3 | 20 | 20 | 0.8 | 0.4 | 1.04 | 1 |
| #5 | βSβS | HbS/HbS | PB | 68.4 | 74.1 | 77.1 | 0.16 | 0.46 | 0.26 | 8 | 0 | - | - | - | - | 0.93 | 0.76 |
| #6 | βSβS | HbS/HbS | PB | 63.5 | 71.5 | 67.1 | 0.25 | 0.24 | 0.23 | 3.6 | 0.7 | 20 | 30 | 0.3 | 0.8 | 1.15 | 1.28 |
| #7 | βSβS | HbS/HbS | PB | 78.6 | 80.2 | 78.9 | 0.17 | 0.15 | 0.19 | 0 | 6 | 100 | 100 | 1.7 | 1.9 | - | - |
| #8 | βSβS | HbS/HbS | PB | 68.5 | 69.9 | 71 | 0.09 | 0.25 | 0.24 | - | - | 56 | 90 | 0.4 | 0.9 | 1.19 | 0.79 |
| #9 | βSβ+ | HbS/ IVS1-110 | PB | 81.9 | 84.5 | 71.5 | 0.21 | 0.23 | 0.29 | 20 | 0 | 30 | 10 | 0.3 | 0.6 | - | - |
| #10 | βSβ+ | HbS/ IVS1-110 | PB | 59.1 | 53.4 | 60.6 | 0.31 | 0.31 | 0.42 | 15.3 | 20.5 | 33.3 | 35.3 | 1.8 | 2.8 | 1.06 | 1.09 |
| #11 | βSβ+ | HbS/ IVS1-110 | PB | 43.6 | 45.3 | 43.8 | 0.30 | 0.28 | 0.30 | 17.8 | 11.7 | 100 | 100 | 0.8 | 0.5 | 1.29 | 1.34 |
| #12 | βSβ+ | HbS/ IVS1-1 | PB | 33.2 | 18.4 | 44.3 | 0.35 | 0.42 | 0.38 | 10.9 | 29.5 | 66.7 | 66.7 | 0.4 | 0.5 | 0.56 | 0.78 |
| #13 | βSβ+ | HbS/ IVS1-110 | PB | 76.9 | 86 | 80.2 | - | - | - | 11.1 | 36 | 37.9 | 44.4 | 1.5 | 1.0 | 1.06 | 0.95 |
| Average | 64 | 65 | 67 | 0.23 | 0.27 | 0.28 | 9.6 | 12 | 52 | 55 | 0.89 | 1.04 | 1.03 | 0.999 | |||
| p-value | 0.59 | 0.19 | 0.30 | 0.02 | 0.66 | 0.82 | 0.65 | 0.75 | |||||||||
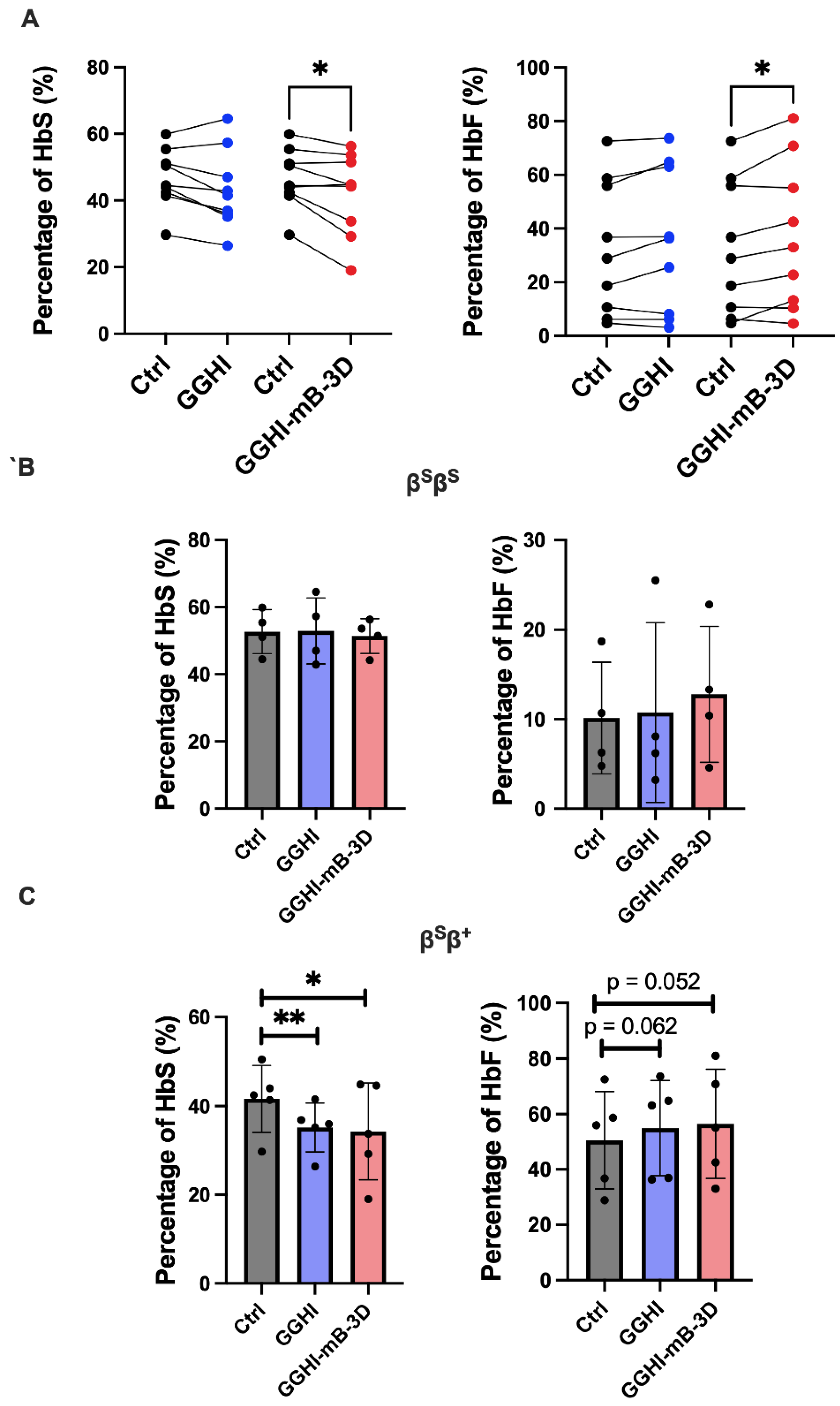
| Sample | Genotype | Mutation | F-Cells (FACS) | Aγ/α Ratio (RP-HPLC) | HbS (%) | HbF (%) | TSD Efficiency (%) | Mean VCN/cell | Relative Fold Difference of γ-mRNA Transcripts | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ctrl | GGHI | GGHI-mB-3D | Ctrl | GGHI | GGHI- mB-3D | Ctrl | GGHI | GGHI- mB-3D | Ctrl | GGHI | GGHI- mB-3D | GGHI | GGHI- mB-3D | GGHI | GGHI- mB-3D | GGHI | GGHI- mB-3D | |||
| #4 | βSβS | HbS/HbS | 66.3 | 69.1 | 71.1 | 0.21 | 0.11 | 0.24 | 55.4 | 57.3 | 53.6 | 4.8 | 3.2 | 13.3 | 20 | 20 | 0.8 | 0.4 | 1.04 | 1 |
| #5 | βSβS | HbS/HbS | 68.4 | 74.1 | 77.1 | 0.16 | 0.46 | 0.26 | 51.1 | 47 | 51.5 | 10.7 | 8.1 | 10.4 | - | - | - | - | 0.93 | 0.76 |
| #6 | βSβS | HbS/HbS | 63.5 | 71.5 | 67.1 | 0.25 | 0.24 | 0.23 | 44.5 | 42.9 | 44.2 | 18.7 | 25.5 | 22.8 | 20 | 30 | 0.3 | 0.8 | 1.15 | 1.28 |
| #7 | βSβS | HbS/HbS | 78.6 | 80.2 | 78.9 | 0.17 | 0.15 | 0.19 | 59.9 | 64.6 | 56.3 | 6.3 | 6.2 | 4.6 | 100 | 100 | 1.7 | 1.9 | - | - |
| #8 | βSβS | HbS/HbS | 68.5 | 69.9 | 71 | 0.09 | 0.25 | 0.24 | - | - | - | - | - | - | 56 | 90 | 0.4 | 0.9 | 1.19 | 0.79 |
| Average | 69 | 73 | 73 | 0.18 | 0.24 | 0.23 | 52.7 | 53 | 51.4 | 10.13 | 10.75 | 12.78 | 49 | 60 | 0.8 | 1 | 1.08 | 0.96 | ||
| p-value | 0.038 | 0.046 | 0.41 | 0.14 | 0.91 | 0.23 | 0.79 | 0.33 | 0.71 | 0.67 | 0.41 | |||||||||
| Sample | Genotype | Mutation | F-Cells (FACS) | Aγ/α Ratio (RP-HPLC) | HbS (%) | HbF (%) | TSD Efficiency (%) | Mean VCN/Cell | Relative Fold Difference of γ-mRNA Transcripts | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ctrl | GGHI | GGHI-mB-3D | Ctrl | GGHI | GGHI-mB-3D | Ctrl | GGHI | GGHI-mB-3D | Ctrl | GGHI | GGHI- mB-3D | GGHI | GGHI- mB-3D | GGHI | GGHI- mB-3D | GGHI | GGHI- mB-3D | |||
| #9 | βSβ+ | HbS/ IVS1-110 | 81.9 | 84.5 | 71.5 | 0.21 | 0.23 | 0.29 | 44.01 | 35.19 | 44.86 | 55.97 | 64.81 | 55.14 | 30 | 10 | 0.3 | 0.6 | - | - |
| #10 | βSβ+ | HbS/ IVS1-110 | 59.1 | 53.4 | 60.6 | 0.31 | 0.31 | 0.42 | 42.45 | 35.96 | 33.77 | 36.77 | 36.92 | 42.53 | 33.3 | 35.3 | 1.8 | 2.8 | 1.06 | 1.09 |
| #11 | βSβ+ | HbS/ IVS1-110 | 43.6 | 45.3 | 43.8 | 0.30 | 0.28 | 0.30 | 50.50 | 41.50 | 44.60 | 28.90 | 36.40 | 33.00 | 100 | 100 | 0.8 | 0.5 | 1.29 | 1.34 |
| #12 | βSβ+ | HbS/ IVS1-1 | 33.2 | 18.4 | 44.3 | 0.35 | 0.42 | 0.38 | 41.40 | 36.90 | 29.20 | 58.70 | 63.10 | 70.80 | 66.7 | 66.7 | 0.4 | 0.5 | 0.56 | 0.78 |
| #13 | βSβ+ | HbS/ IVS1-110 | 76.9 | 86 | 80.2 | - | - | - | 29.70 | 26.40 | 19.00 | 72.50 | 73.60 | 81.00 | 37.9 | 44.4 | 1.5 | 1.0 | 1.06 | 0.95 |
| Average | 59 | 58 | 60 | 0.29 | 0.31 | 0.35 | 41.61 | 35.19 | 34.29 | 50.57 | 54.97 | 56.49 | 54 | 51 | 0.96 | 1.1 | 0.99 | 1.04 | ||
| p-value | 0.75 | 0.76 | 0.45 | 0.12 | 0.005 | 0.03 | 0.062 | 0.052 | 0.91 | 0.83 | 0.82 | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drakopoulou, E.; Georgomanoli, M.; Lederer, C.W.; Panetsos, F.; Kleanthous, M.; Voskaridou, E.; Valakos, D.; Papanikolaou, E.; Anagnou, N.P. The Optimized γ-Globin Lentiviral Vector GGHI-mB-3D Leads to Nearly Therapeutic HbF Levels In Vitro in CD34+ Cells from Sickle Cell Disease Patients. Viruses 2022, 14, 2716. https://doi.org/10.3390/v14122716
Drakopoulou E, Georgomanoli M, Lederer CW, Panetsos F, Kleanthous M, Voskaridou E, Valakos D, Papanikolaou E, Anagnou NP. The Optimized γ-Globin Lentiviral Vector GGHI-mB-3D Leads to Nearly Therapeutic HbF Levels In Vitro in CD34+ Cells from Sickle Cell Disease Patients. Viruses. 2022; 14(12):2716. https://doi.org/10.3390/v14122716
Chicago/Turabian StyleDrakopoulou, Ekati, Maria Georgomanoli, Carsten W. Lederer, Fottes Panetsos, Marina Kleanthous, Ersi Voskaridou, Dimitrios Valakos, Eleni Papanikolaou, and Nicholas P. Anagnou. 2022. "The Optimized γ-Globin Lentiviral Vector GGHI-mB-3D Leads to Nearly Therapeutic HbF Levels In Vitro in CD34+ Cells from Sickle Cell Disease Patients" Viruses 14, no. 12: 2716. https://doi.org/10.3390/v14122716
APA StyleDrakopoulou, E., Georgomanoli, M., Lederer, C. W., Panetsos, F., Kleanthous, M., Voskaridou, E., Valakos, D., Papanikolaou, E., & Anagnou, N. P. (2022). The Optimized γ-Globin Lentiviral Vector GGHI-mB-3D Leads to Nearly Therapeutic HbF Levels In Vitro in CD34+ Cells from Sickle Cell Disease Patients. Viruses, 14(12), 2716. https://doi.org/10.3390/v14122716