
In Vitro and Pre-Clinical Evaluation of Locally Isolated Phages, vB_Pae_SMP1 and vB_Pae_SMP5, Formulated as Hydrogels against Carbapenem-Resistant Pseudomonas aeruginosa

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Bacterial Isolates: Collection, Identification, and Antimicrobial Susceptibility Testing
2.2. Phenotypic and Genotypic Detection of Carbapenemase Producers (CPs)
2.3. Bacteriophage Recovery from Sewage Samples
2.3.1. Isolation of P. aeruginosa-Specific Bacteriophages
2.3.2. Screening for Lytic Activity against CRPA in the Acquired Lysates
2.3.3. Phage Propagation
2.4. Characterization of the Isolated Bacteriophages Showing Lytic Activities against CRPA
2.4.1. Host Range
2.4.2. Morphology of the Isolated Bacteriophages Showing Lytic Activities against CRPA
2.4.3. Molecular Analysis of the Bacteriophage
2.5. Formulation of Bacteriophage-Carboxymethyl Cellulose (CMC) Hydrogel
2.6. In Vitro Anti-CRPA Activity of the Tested Hydrogels
2.7. Preclinical Evaluation of the Formulated Tested Hydrogels
2.7.1. Laboratory Animals
2.7.2. Thermal Injury Model
- Group I: Control, Burned, non-infected, untreated.
- Group II: Control, burned, infected, untreated.
- Group III: Control, Burned, infected, treated with vehicle (control hydrogel).
- Group IV: Burned, infected, treated with tested hydrogel-1 (phage vB_Pae_SMP1)
- Group V: Burned, infected, treated with tested hydrogel-2 (phage P5).
- Group VI: Burned, infected, treated with phage cocktail hydrogel (Phage vB_Pae_SMP1 + phage P5).
- Group VII: Positive Control, burned, infected, treated with Silver sulfadiazine 1% (Silvirburn®, MUP Co., Cairo, Egypt).
- Group VIII: Positive Control, burned, infected, treated with Collagenase 0.6 IU (Iruxol ®, Abbott Co., Wiesbaden, Germany).
- Group IX: Normal Control, intact, non-infected, untreated.
2.7.3. Treatment
2.7.4. Histopathological Examination
2.8. Statistical Analysis
3. Results
3.1. Identification, Antimicrobial Susceptibility, Phenotypic and Genotypic Analysis of the Recovered P. aeruginosa Isolates
3.2. Recovery of Bacteriophages and Screening for the Activity against CRPA
3.3. Characterization of the Isolated Bacteriophages Showing Lytic Activities against CRPA
3.3.1. Host Range
3.3.2. Morphology of the Bacteriophages vB_Pae_SMP1 and vB_Pae_SMP5 as Presented by TEM
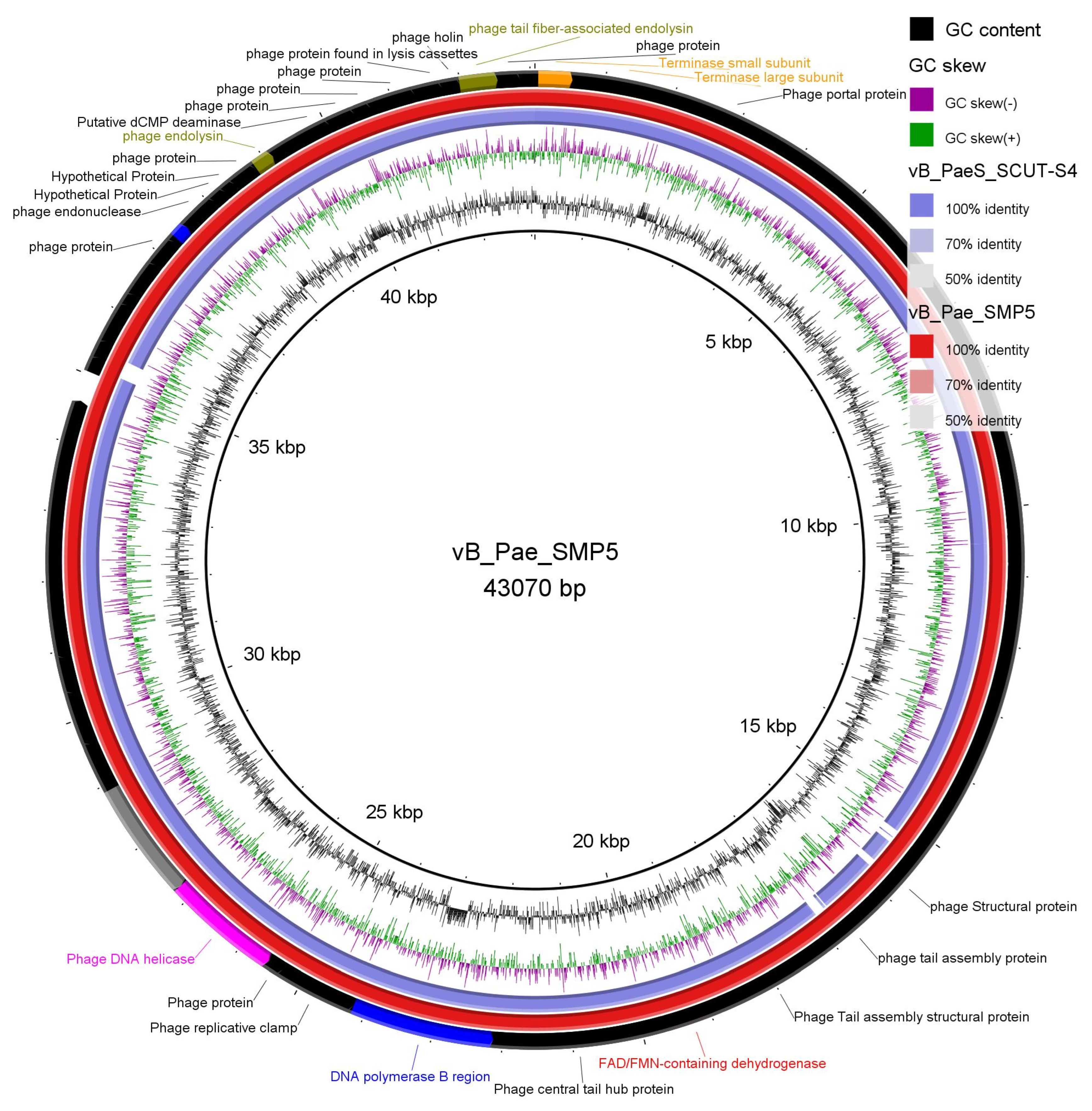
3.3.3. Phage Lysate Sequencing Results a Complete Genome of Phage vB_Pae_SMP5
3.4. In Vitro Anti-CRPA Activity of the Tested Hydrogels
3.5. In Vivo Anti-CRPA Activity of the Formulated Tested Hydrogels
3.5.1. Survival Rate
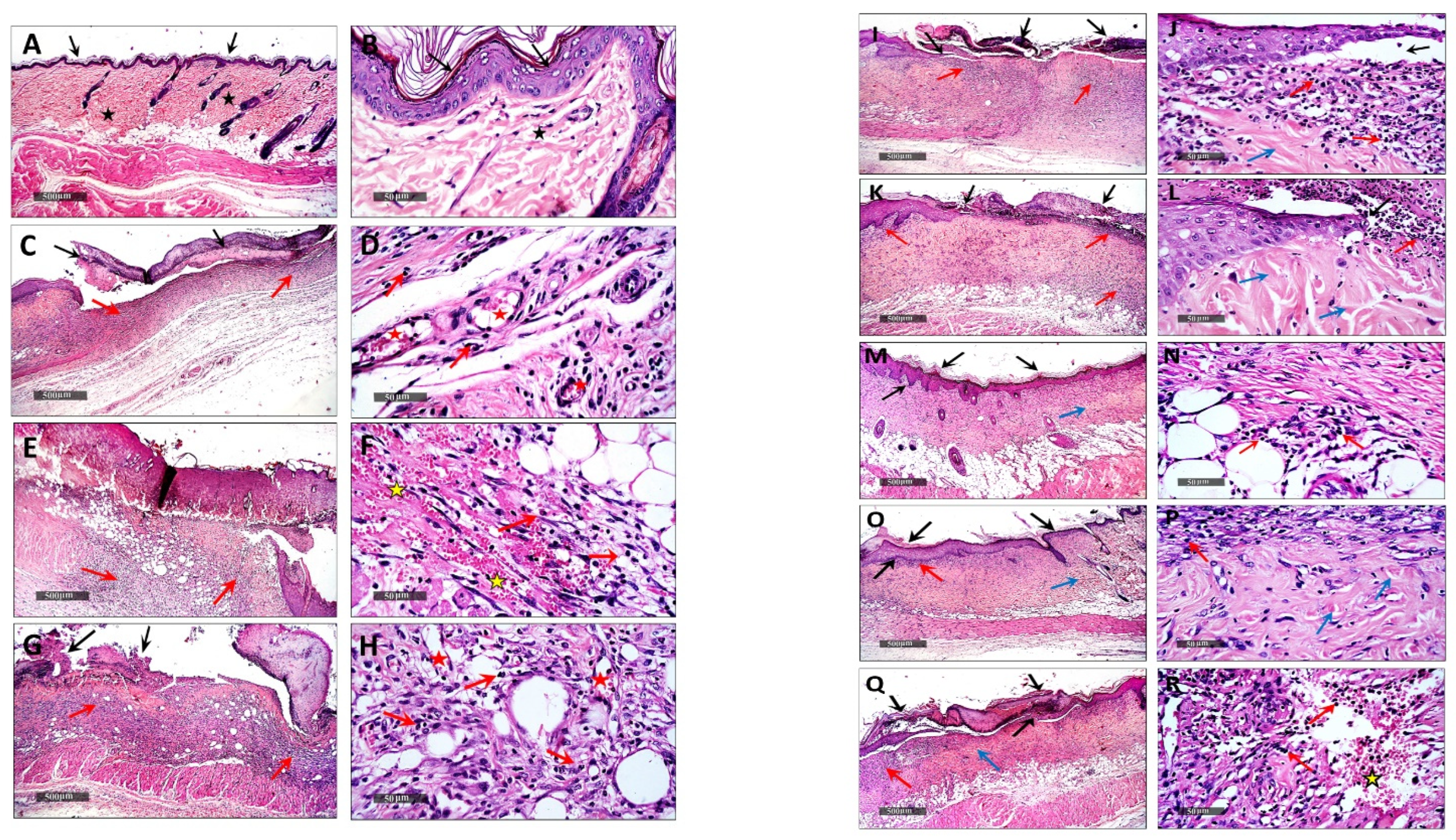
3.5.2. Histopathological Examination
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diekema, D.J.; Hsueh, P.-R.; Mendes, R.E.; Pfaller, M.A.; Rolston, K.V.; Sader, H.S.; Jones, R.N. The Microbiology of Bloodstream Infection: 20-Year Trends from the SENTRY Antimicrobial Surveillance Program. Antimicrob. Agents Chemother. 2019, 63, e00355-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiner, L.M.; Webb, A.K.; Limbago, B.; Dudeck, M.A.; Patel, J.; Kallen, A.J.; Edwards, J.R.; Sievert, D.M. Antimicrobial-Resistant Pathogens Associated with Healthcare-Associated Infections: Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2011–2014. Infect. Control Hosp. Epidemiol. 2016, 37, 1288–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadri, S.S. Key Takeaways from the US CDC’s 2019 Antibiotic Resistance Threats Report for Frontline Providers. Crit. Care Med. 2020, 48, 939–945. [Google Scholar] [PubMed]
- Motbainor, H.; Bereded, F.; Mulu, W. Multi-Drug Resistance of Blood Stream, Urinary Tract and Surgical Site Nosocomial Infections of Acinetobacter baumannii and Pseudomonas aeruginosa among Patients Hospitalized at Felegehiwot Referral Hospital, Northwest Ethiopia: A Cross-Sectional Study. BMC Infect. Dis. 2020, 20, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimer, N.A. Nosocomial Infection and Antibiotic-Resistant Threat in the Middle East. Infect. Drug Resist. 2022, 15, 631. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Garg, J.; Kumar, S.; Bhattacharya, A.; Agarwal, S.; Upadhyay, G.C. Molecular Epidemiology & Therapeutic Options of Carbapenem-Resistant Gram-Negative Bacteria. Indian J. Med. Res. 2019, 149, 285. [Google Scholar]
- Paul, M.; Carrara, E.; Retamar, P.; Tängdén, T.; Bitterman, R.; Bonomo, R.A.; De Waele, J.; Daikos, G.L.; Akova, M.; Harbarth, S. European Society of Clinical Microbiology and Infectious Diseases (ESCMID) Guidelines for the Treatment of Infections Caused by Multidrug-Resistant Gram-Negative Bacilli (Endorsed by European Society of Intensive Care Medicine). Clin. Microbiol. Infect. 2022, 28, 521–547. [Google Scholar] [CrossRef]
- Zhen, X.; Stålsby Lundborg, C.; Sun, X.; Gu, S.; Dong, H. Clinical and Economic Burden of Carbapenem-Resistant Infection or Colonization Caused by Klebsiella Pneumoniae, Pseudomonas aeruginosa, Acinetobacter baumannii: A Multicenter Study in China. Antibiotics 2020, 9, 514. [Google Scholar] [CrossRef]
- Azimi, L.; Lari, A.R. Colistin-Resistant Pseudomonas aeruginosa Clinical Strains with Defective Biofilm Formation. GMS Hyg. Infect. Control 2019, 14, 1–6. [Google Scholar] [CrossRef]
- Erol, H.B.; Kaskatepe, B. Comparison of the in Vitro Efficacy of Commercial Bacteriophage Cocktails and Isolated Bacteriophage vB_Pa01 against Carbapenem Resistant Nosocomial Pseudomonas aeruginosa. J. Res. Pharm. 2021, 25, 407–414. [Google Scholar]
- Jahangiri, A.; Neshani, A.; Mirhosseini, S.A.; Ghazvini, K.; Zare, H.; Sedighian, H. Synergistic Effect of Two Antimicrobial Peptides, Nisin and P10 with Conventional Antibiotics against Extensively Drug-Resistant Acinetobacter baumannii and Colistin-Resistant Pseudomonas aeruginosa Isolates. Microb. Pathog. 2021, 150, 104700. [Google Scholar] [PubMed]
- Broncano-Lavado, A.; Santamaría-Corral, G.; Esteban, J.; García-Quintanilla, M. Advances in Bacteriophage Therapy against Relevant Multidrug-Resistant Pathogens. Antibiotics 2021, 10, 672. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Liu, Z.; Tan, X.; Wang, H.; Liang, Y.; Kong, Y.; Sun, W.; Sun, L.; Ma, Y.; Lu, H. Bacteriophage Therapy for Empyema Caused by Carbapenem-Resistant Pseudomonas aeruginosa. Biosci. Trends 2022, 16, 158–162. [Google Scholar] [PubMed]
- Fathima, B.; Archer, A.C. Bacteriophage Therapy: Recent Developments and Applications of a Renaissant Weapon. Res. Microbiol. 2021, 172, 103863. [Google Scholar]
- Erol, H.B.; Kaskatepe, B. Isolation of newly isolated vb_k1 bacteriophage and investigation of susceptibility on esbl positive Klebsiella spp. strains. J. Fac. Pharm. Ankara Univ. 2021, 45, 515–523. [Google Scholar] [CrossRef]
- Kalelkar, P.P.; Riddick, M.; García, A.J. Biomaterial-Based Antimicrobial Therapies for the Treatment of Bacterial Infections. Nat. Rev. Mater. 2022, 7, 39–54. [Google Scholar]
- Rivera, D.; Moreno-Switt, A.I.; Denes, T.G.; Hudson, L.K.; Peters, T.L.; Samir, R.; Aziz, R.K.; Noben, J.-P.; Wagemans, J.; Dueñas, F. Novel Salmonella Phage, vB_Sen_STGO-35-1, Characterization and Evaluation in Chicken Meat. Microorganisms 2022, 10, 606. [Google Scholar]
- Sakr, M.M.; Elkhatib, W.F.; Aboshanab, K.M.; Mantawy, E.M.; Yassien, M.A.; Hassouna, N.A. In Vivo Evaluation of a Recombinant N-Acylhomoserine Lactonase Formulated in a Hydrogel Using a Murine Model Infected with MDR Pseudomonas aeruginosa Clinical Isolate, CCASUP2. AMB Express 2021, 11, 109. [Google Scholar] [CrossRef]
- Abd-Allah, I.M.; El-Housseiny, G.S.; Alshahrani, M.Y.; El-Masry, S.S.; Aboshanab, K.M.; Hassouna, N.A. An Anti-MRSA Phage From Raw Fish Rinse: Stability Evaluation and Production Optimization. Front. Cell. Infect. Microbiol. 2022, 12, 904531. [Google Scholar] [CrossRef]
- Camens, S.; Liu, S.; Hon, K.; Bouras, G.S.; Psaltis, A.J.; Wormald, P.-J.; Vreugde, S. Preclinical Development of a Bacteriophage Cocktail for Treating Multidrug Resistant Pseudomonas aeruginosa Infections. Microorganisms 2021, 9, 2001. [Google Scholar] [CrossRef]
- Kim, H.Y.; Chang, R.Y.K.; Morales, S.; Chan, H.K. Bacteriophage-Delivering Hydrogels: Current Progress in Combating Antibiotic Resistant Bacterial Infection. Antibiotics 2021, 10, 130. [Google Scholar] [CrossRef] [PubMed]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A. Bergey’s Manual of Determinative Bacterology, 9th ed.; Lippincott Williams and Wilkins: Baltimore, Maryland, 1994; Available online: https://www.scirp.org/(S(i43dyn45teexjx455qlt3d2q))/reference/ReferencesPapers.aspx?ReferenceID=42336 (accessed on 7 December 2022).
- Shetty, N.; Hill, G.; Ridgway, G.L. The Vitek Analyser for Routine Bacterial Identification and Susceptibility Testing: Protocols, Problems, and Pitfalls. J. Clin. Pathol. 1998, 51, 316–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, M.P. Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021; ISBN 1684401046. [Google Scholar]
- Magiorakos, A.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B. Multidrug-resistant, Extensively Drug-resistant and Pandrug-resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Mabrouk, S.S.; Abdellatif, G.R.; El-Ansary, M.R.; Aboshanab, K.M.; Ragab, Y.M. Carbapenemase Producers Among Extensive Drug-Resistant Gram-Negative Pathogens Recovered from Febrile Neutrophilic Patients in Egypt. Infect. Drug Resist. 2020, 13, 3113. [Google Scholar] [CrossRef] [PubMed]
- Wommack, K.E.; Williamson, K.E.; Helton, R.R.; Bench, S.R.; Winget, D.M. Methods for the Isolation of Viruses from Environmental Samples. In Bacteriophages; Springer: Berlin/Heidelberg, Germany, 2009; pp. 3–14. [Google Scholar]
- Ackermann, H.-W. Basic Phage Electron Microscopy. In Bacteriophages; Springer: Berlin/Heidelberg, Germany, 2009; pp. 113–126. [Google Scholar]
- El-Dougdoug, N.; Nasr-Eldin, M.; Azzam, M.; Mohamed, A.; Hazaa, M. Improving Wastewater Treatment Using Dried Banana Leaves and Bacteriophage Cocktail. Egypt. J. Bot. 2020, 60, 199–212. [Google Scholar] [CrossRef]
- Hussein, Y.S.; El-Masry, S.S.; Faiesal, A.A.; El-Dougdoug, K.A.; Othman, B.A. Physico-chemical properties of some listeria phages. Arab Univ. J. Agric. Sci. 2019, 27, 175–183. [Google Scholar] [CrossRef]
- Adams, M.H. Bacteriophages; Citeseer: 1959; Wiley Interscience: New York, NY, USA; Available online: https://www.scirp.org/(S(351jmbntvnsjt1aadkposzje))/reference/ReferencesPapers.aspx?ReferenceID=1855130 (accessed on 7 December 2022).
- Anderson, B.; Rashid, M.H.; Carter, C.; Pasternack, G.; Rajanna, C.; Revazishvili, T.; Dean, T.; Senecal, A.; Sulakvelidze, A. Enumeration of Bacteriophage Particles: Comparative Analysis of the Traditional Plaque Assay and Real-Time QPCR-and Nanosight-Based Assays. Bacteriophage 2011, 1, 86–93. [Google Scholar] [CrossRef] [Green Version]
- Kropinski, A.M. Bacteriophages, Methods and Protocols, Volume 1: Isolation, Characterization, and Interactions; Clokie, M.R.J., Kropinski, A.M., Eds.; Part of the book series: Methods in Molecular Biology; Humana Totowa: New York, NY, USA, 2009; Volume 501. [Google Scholar] [CrossRef]
- Carlson, K. Working with Bacteriophages: Common Techniques and Methodological Approaches; CRC Press: Boca Raton, FL, USA, 2005; Volume 1. [Google Scholar]
- Kutter, E. Phage Host Range and Efficiency of Plating. In Bacteriophages; Springer: Berlin/Heidelberg, Germany, 2009; pp. 141–149. [Google Scholar]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M. The PATRIC Bioinformatics Resource Center: Expanding Data and Analysis Capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef] [Green Version]
- Parrello, B.; Butler, R.; Chlenski, P.; Pusch, G.D.; Overbeek, R. Supervised Extraction of Near-Complete Genomes from Metagenomic Samples: A New Service in PATRIC. PLoS ONE 2021, 16, e0250092. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M. The RAST Server: Rapid Annotations Using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- McNair, K.; Aziz, R.K.; Pusch, G.D.; Overbeek, R.; Dutilh, B.E.; Edwards, R. Phage Genome Annotation Using the RAST Pipeline. In Bacteriophages; Springer: Berlin/Heidelberg, Germany, 2018; pp. 231–238. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Alikhan, N.-F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple Prokaryote Genome Comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khullar, R.; Saini, S.; Seth, N.; Rana, A.C. Emulgels: A Surrogate Approach for Topically Used Hydrophobic Drugs. Int. J. Pharm. Bio. Sci. 2011, 1, 117–128. [Google Scholar]
- Singh, B.; Sharma, S.; Dhiman, A. Design of Antibiotic Containing Hydrogel Wound Dressings: Biomedical Properties and Histological Study of Wound Healing. Int. J. Pharm. 2013, 457, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, P.E.; Abdelkader, N.F.; El Awdan, S.A.; El-Shabrawy, O.A.; Zaki, H.F. Liraglutide Ameliorated Peripheral Neuropathy in Diabetic Rats: Involvement of Oxidative Stress, Inflammation and Extracellular Matrix Remodeling. J. Neurochem. 2018, 146, 173–185. [Google Scholar] [CrossRef]
- Werner, M.; Chott, A.; Fabiano, A.; Battifora, H. Effect of Formalin Tissue Fixation and Processing on Immunohistochemistry. Am. J. Surg. Pathol. 2000, 24, 1016–1019. [Google Scholar] [CrossRef]
- Almukainzi, M.; El-Masry, T.A.; Negm, W.A.; Elekhnawy, E.; Saleh, A.; Sayed, A.E.; Khattab, M.A.; Abdelkader, D.H. Gentiopicroside PLGA Nanospheres: Fabrication, in Vitro Characterization, Antimicrobial Action, and in Vivo Effect for Enhancing Wound Healing in Diabetic Rats. Int. J. Nanomed. 2022, 17, 1203. [Google Scholar] [CrossRef]
- Culling, C.F.A. Handbook of Histopathological and Histochemical Techniques: Including Museum Techniques; Butterworth-Heinemann: Oxford, UK, 2013; ISBN 1483164799. [Google Scholar]
- Shrivastava, S.R.; Shrivastava, P.S.; Ramasamy, J. World Health Organization Releases Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. J. Med. Soc. 2018, 32, 76. [Google Scholar] [CrossRef]
- Abedon, S.T.; García, P.; Mullany, P.; Aminov, R. Phage Therapy: Past, Present and Future. Front. Microbiol. 2017, 8, 981. [Google Scholar] [CrossRef] [Green Version]
- Jennes, S.; Merabishvili, M.; Soentjens, P.; Pang, K.W.; Rose, T.; Keersebilck, E.; Soete, O.; François, P.-M.; Teodorescu, S.; Verween, G. Use of Bacteriophages in the Treatment of Colistin-Only-Sensitive Pseudomonas aeruginosa Septicaemia in a Patient with Acute Kidney Injury—A Case Report. Crit. Care 2017, 21, 129. [Google Scholar] [CrossRef] [Green Version]
- Nir-Paz, R.; Gelman, D.; Khouri, A.; Sisson, B.M.; Fackler, J.; Alkalay-Oren, S.; Khalifa, L.; Rimon, A.; Yerushalmy, O.; Bader, R. Successful Treatment of Antibiotic-Resistant, Poly-Microbial Bone Infection with Bacteriophages and Antibiotics Combination. Clin. Infect. Dis. 2019, 69, 2015–2018. [Google Scholar] [PubMed]
- Domingo-Calap, P.; Delgado-Martínez, J. Bacteriophages: Protagonists of a Post-Antibiotic Era. Antibiotics 2018, 7, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurie, C.D.; Hogan, B.K.; Murray, C.K.; Loo, F.L.; Hospenthal, D.R.; Cancio, L.C.; Kim, S.H.; Renz, E.M.; Barillo, D.; Holcomb, J.B. Contribution of Bacterial and Viral Infections to Attributable Mortality in Patients with Severe Burns: An Autopsy Series. Burns 2010, 36, 773–779. [Google Scholar]
- Chen, F.; Cheng, X.; Li, J.; Yuan, X.; Huang, X.; Lian, M.; Li, W.; Huang, T.; Xie, Y.; Liu, J. Novel Lytic Phages Protect Cells and Mice against Pseudomonas aeruginosa Infection. J. Virol. 2021, 95, e01832-20. [Google Scholar] [CrossRef]
- Aghaee, B.L.; Mirzaei, M.K.; Alikhani, M.Y.; Mojtahedi, A. Sewage and Sewage-Contaminated Environments Are the Most Prominent Sources to Isolate Phages against Pseudomonas aeruginosa. BMC Microbiol. 2021, 21, 132. [Google Scholar] [CrossRef]
- Bhat, T.; Cao, A.; Yin, J. Virus-like Particles: Measures and Biological Functions. Viruses 2022, 14, 383. [Google Scholar]
- Hyman, P. Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef]
- Park, D.-W.; Lim, G.; Lee, Y.; Park, J.-H. Characteristics of Lytic Phage vB_EcoM-ECP26 and Reduction of Shiga-Toxin Producing Escherichia coli on Produce Romaine. Appl. Biol. Chem. 2020, 63, 19. [Google Scholar]
- Vukotic, G.; Obradovic, M.; Novovic, K.; Di Luca, M.; Jovcic, B.; Fira, D.; Neve, H.; Kojic, M.; McAuliffe, O. Characterization, Antibiofilm, and Depolymerizing Activity of Two Phages Active on Carbapenem-Resistant Acinetobacter baumannii. Front. Med. 2020, 7, 426. [Google Scholar] [CrossRef]
- Duc, H.M.; Son, H.M.; Ngan, P.H.; Sato, J.; Masuda, Y.; Honjoh, K.; Miyamoto, T. Isolation and Application of Bacteriophages Alone or in Combination with Nisin against Planktonic and Biofilm Cells of Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2020, 104, 5145–5158. [Google Scholar] [CrossRef]
- Dakheel, K.H.; Rahim, R.A.; Neela, V.K.; Al-Obaidi, J.R.; Hun, T.G.; Isa, M.N.M.; Yusoff, K. Genomic Analyses of Two Novel Biofilm-Degrading Methicillin-Resistant Staphylococcus Aureus Phages. BMC Microbiol. 2019, 19, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klumpp, J.; Loessner, M.J. Listeria Phages: Genomes, Evolution, and Application. Bacteriophage 2013, 3, e26861. [Google Scholar] [PubMed] [Green Version]
- Ross, A.; Ward, S.; Hyman, P. More Is Better: Selecting for Broad Host Range Bacteriophages. Front. Microbiol. 2016, 7, 1352. [Google Scholar] [PubMed] [Green Version]
- de Jonge, P.A.; Nobrega, F.L.; Brouns, S.J.J.; Dutilh, B.E. Molecular and Evolutionary Determinants of Bacteriophage Host Range. Trends Microbiol. 2019, 27, 51–63. [Google Scholar]
- Turner, D.; Kropinski, A.M.; Adriaenssens, E.M. A Roadmap for Genome-Based Phage Taxonomy. Viruses 2021, 13, 506. [Google Scholar]
- Othman, B.A.; Askora, A.; Abo-Senna, A.S.M. Isolation and Characterization of a Siphoviridae Phage Infecting Bacillus megaterium from a Heavily Trafficked Holy Site in Saudi Arabia. Folia Microbiol. 2015, 60, 289–295. [Google Scholar]
- Ackermann, H.-W. Bacteriophage Electron Microscopy. Adv. Virus Res. 2012, 82, 1–32. [Google Scholar]
- King, A.M.Q.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: Amsterdam, The Netherlands, 2011; Volume 9, ISBN 0123846854. [Google Scholar]
- Sezer, S.; Şahin, İ.; Öztürk, K.; Şanko, V.; Koçer, Z.; Sezer, Ü.A. Cellulose-Based Hydrogels as Biomaterials. In Cellulose-Based Superabsorbent Hydrogels; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1177–1203. [Google Scholar]
- Alibolandi, M.; Bagheri, E.; Mohammadi, M.; Sameiyan, E.; Ramezani, M. Biopolymer-Based Hydrogel Wound Dressing. In Modeling and Control of Drug Delivery Systems; Elsevier: Amsterdam, The Netherlands, 2021; pp. 227–251. [Google Scholar]
- Li, Z.; He, C.; Yuan, B.; Dong, X.; Chen, X. Injectable Polysaccharide Hydrogels as Biocompatible Platforms for Localized and Sustained Delivery of Antibiotics for Preventing Local Infections. Macromol. Biosci. 2017, 17, 1600347. [Google Scholar]
- El-Gayar, M.H.; Ishak, R.A.H.; Esmat, A.; Aboulwafa, M.M.; Aboshanab, K.M. Evaluation of Lyophilized Royal Jelly and Garlic Extract Emulgels Using a Murine Model Infected with Methicillin-Resistant Staphylococcus aureus. AMB Express 2022, 12, 37. [Google Scholar] [CrossRef]
- Masson-Meyers, D.S.; Andrade, T.A.M.; Caetano, G.F.; Guimaraes, F.R.; Leite, M.N.; Leite, S.N.; Frade, M.A.C. Experimental Models and Methods for Cutaneous Wound Healing Assessment. Int. J. Exp. Pathol. 2020, 101, 21–37. [Google Scholar]
- Pires, D.P.; Vilas Boas, D.; Sillankorva, S.; Azeredo, J. Phage Therapy: A Step Forward in the Treatment of Pseudomonas aeruginosa Infections. J. Virol. 2015, 89, 7449–7456. [Google Scholar] [PubMed] [Green Version]
- McVay, C.S.; Velásquez, M.; Fralick, J.A. Phage Therapy of Pseudomonas aeruginosa Infection in a Mouse Burn Wound Model. Antimicrob. Agents Chemother. 2007, 51, 1934–1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghadam, M.T.; Khoshbayan, A.; Chegini, Z.; Farahani, I.; Shariati, A. Bacteriophages, a New Therapeutic Solution for Inhibiting Multidrug-Resistant Bacteria Causing Wound Infection: Lesson from Animal Models and Clinical Trials. Drug Des. Devel. Ther. 2020, 14, 1867. [Google Scholar]
- Jault, P.; Leclerc, T.; Jennes, S.; Pirnay, J.P.; Que, Y.-A.; Resch, G.; Rousseau, A.F.; Ravat, F.; Carsin, H.; Le Floch, R. Efficacy and Tolerability of a Cocktail of Bacteriophages to Treat Burn Wounds Infected by Pseudomonas aeruginosa (PhagoBurn): A Randomised, Controlled, Double-Blind Phase 1/2 Trial. Lancet Infect. Dis. 2019, 19, 35–45. [Google Scholar] [PubMed]
- Koehler, J.; Brandl, F.P.; Goepferich, A.M. Hydrogel Wound Dressings for Bioactive Treatment of Acute and Chronic Wounds. Eur. Polym. J. 2018, 100, 1–11. [Google Scholar] [CrossRef]
- Maura, D.; Bandyopadhaya, A.; Rahme, L.G. Animal Models for Pseudomonas aeruginosa Quorum Sensing Studies. In Quorum Sensing; Springer: Berlin/Heidelberg, Germany, 2018; pp. 227–241. [Google Scholar]
- Durmus, A.S.; Han, M.C.; Yaman, I. Comperative Evaluation of Collagenase and Silver Sulfadiazine on Burned Wound Healing in Rats. Firat Univ. Saglik Bilim. Vet. Derg. 2009, 23, 135–139. [Google Scholar]
- Mekkes, J.R.; Zeegelaar, J.E.; Westerhof, W. Quantitative and Objective Evaluation of Wound Debriding Properties of Collagenase and Fibrinolysin/Desoxyribonuclease in a Necrotic Ulcer Animal Model. Arch. Dermatol. Res. 1998, 290, 152–157. [Google Scholar]
- Khan, M.A.; Hussain, Z.; Ali, S.; Qamar, Z.; Imran, M.; Hafeez, F.Y. Fabrication of Electrospun Probiotic Functionalized Nanocomposite Scaffolds for Infection Control and Dermal Burn Healing in a Mice Model. ACS Biomater. Sci. Eng. 2019, 5, 6109–6116. [Google Scholar] [CrossRef]
- Pereira Beserra, F.; Sérgio Gushiken, L.F.; Vieira, A.J.; Augusto Bérgamo, D.; Luísa Bérgamo, P.; Oliveira de Souza, M.; Alberto Hussni, C.; Kiomi Takahira, R.; Henrique Nóbrega, R.; Monteiro Martinez, E.R. From Inflammation to Cutaneous Repair: Topical Application of Lupeol Improves Skin Wound Healing in Rats by Modulating the Cytokine Levels, NF-ΚB, Ki-67, Growth Factor Expression, and Distribution of Collagen Fibers. Int. J. Mol. Sci. 2020, 21, 4952. [Google Scholar] [CrossRef]
- Steele, A.; Stacey, H.J.; De Soir, S.; Jones, J.D. The Safety and Efficacy of Phage Therapy for Superficial Bacterial Infections: A Systematic Review. Antibiotics 2020, 9, 754. [Google Scholar]
- Mendes, J.J.; Leandro, C.; Corte-Real, S.; Barbosa, R.; Cavaco-Silva, P.; Melo-Cristino, J.; Górski, A.; Garcia, M. Wound Healing Potential of Topical Bacteriophage Therapy on Diabetic Cutaneous Wounds. Wound Repair Regen. 2013, 21, 595–603. [Google Scholar] [PubMed]
- Gupta, P.; Singh, H.S.; Shukla, V.K.; Nath, G.; Bhartiya, S.K. Bacteriophage Therapy of Chronic Nonhealing Wound: Clinical Study. Int. J. Low. Extrem. Wounds 2019, 18, 171–175. [Google Scholar] [PubMed]
- Khalid, F.; Siddique, A.B.; Nawaz, Z.; Shafique, M.; Zahoor, M.A.; Nisar, M.A.; Rasool, M.H. Efficacy of Bacteriophage against Multidrug Resistant Pseudomonas aeruginosa Isolates. Southeast Asian J. Trop. Med. Public Health 2017, 48, 1056–1062. [Google Scholar]
- Raz, A.; Serrano, A.; Hernandez, A.; Euler, C.W.; Fischetti, V.A. Isolation of Phage Lysins That Effectively Kill Pseudomonas aeruginosa in Mouse Models of Lung and Skin Infection. Antimicrob. Agents Chemother. 2019, 63, e00024-19. [Google Scholar] [PubMed] [Green Version]
- Pinto, A.M.; Cerqueira, M.A.; Bañobre-Lópes, M.; Pastrana, L.M.; Sillankorva, S. Bacteriophages for Chronic Wound Treatment: From Traditional to Novel Delivery Systems. Viruses 2020, 12, 235. [Google Scholar] [PubMed]
- Chhibber, S.; Kaur, J.; Kaur, S. Liposome Entrapment of Bacteriophages Improves Wound Healing in a Diabetic Mouse MRSA Infection. Front. Microbiol. 2018, 9, 561. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate Code | Susceptibility Pattern | MIC of IMP (μg /mL) | Phenotypic Tests | Carbapenemase Genes | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| AK | AT | CIP | LEV | IMP | MER | CDT | mCIM | BCT | |||
| CRPA1 | R | R | R | R | R | R | >1024 | − | − | + | blaKPC |
| CRPA2 | R | R | R | R | R | R | >512 | − | + | + | blaOXA-48 |
| CRPA3 | R | R | R | R | R | R | 32 | − | + | + | blaOXA-48 |
| Isolate Code | Microorganism | Spot Test | |
|---|---|---|---|
| Lysate vB_Pae_SMP1 | Lysate vB_Pae_SMP5 | ||
| CRPA1 | P. aeruginosa | − | + |
| CRPA2 | P. aeruginosa | + | + |
| CRPA3 | P. aeruginosa | + | − |
| Parameters | Phage vB_Pae_SMP5 |
|---|---|
| Molecular type | Genomic DNA |
| Genomic size (bp) | 43070 bp |
| Proteins/ORFs | 57 (46 coded by + frames and 11 coded by - frames) |
| Isolation Source | sewage |
| Host | XDR Pseudomonas aeruginosa clinical isolates CRPA1 and CRPA2 |
| Class | Caudoviricetes |
| Family | Currently unassigned (formerly Siphoviridae, suggested Septimatreviridae) |
| Genus | Septimatrevirus |
| Species | unclassified |
| Group | Description | Rats Survival % |
|---|---|---|
| I | Normal Control, intact, non-infected, untreated | 100 |
| II | Control, Burned, non-infected, untreated | 80 |
| III | Control, burned, infected, untreated | 40 |
| IV | Control, Burned, infected, treated with control hydrogel | 60 |
| V | Burned, infected, treated with Phage 1 hydrogel | 100 |
| VI | Burned, infected, treated with phage 5 hydrogel | 100 |
| VII | burned, infected, treated with phage cocktail hydrogel (vB_Pae_SMP1 + vB_Pae_SMP5) | 100 |
| VIII | Positive Control, burned, infected, treated with Silverburn® | 100 |
| IX | Positive Control, burned, infected, treated with Iruxol® | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
S. Mabrouk, S.; Abdellatif, G.R.; Abu Zaid, A.S.; Aziz, R.K.; Aboshanab, K.M. In Vitro and Pre-Clinical Evaluation of Locally Isolated Phages, vB_Pae_SMP1 and vB_Pae_SMP5, Formulated as Hydrogels against Carbapenem-Resistant Pseudomonas aeruginosa. Viruses 2022, 14, 2760. https://doi.org/10.3390/v14122760
S. Mabrouk S, Abdellatif GR, Abu Zaid AS, Aziz RK, Aboshanab KM. In Vitro and Pre-Clinical Evaluation of Locally Isolated Phages, vB_Pae_SMP1 and vB_Pae_SMP5, Formulated as Hydrogels against Carbapenem-Resistant Pseudomonas aeruginosa. Viruses. 2022; 14(12):2760. https://doi.org/10.3390/v14122760
Chicago/Turabian StyleS. Mabrouk, Samar, Ghada R. Abdellatif, Ahmed S. Abu Zaid, Ramy K. Aziz, and Khaled M. Aboshanab. 2022. "In Vitro and Pre-Clinical Evaluation of Locally Isolated Phages, vB_Pae_SMP1 and vB_Pae_SMP5, Formulated as Hydrogels against Carbapenem-Resistant Pseudomonas aeruginosa" Viruses 14, no. 12: 2760. https://doi.org/10.3390/v14122760
APA StyleS. Mabrouk, S., Abdellatif, G. R., Abu Zaid, A. S., Aziz, R. K., & Aboshanab, K. M. (2022). In Vitro and Pre-Clinical Evaluation of Locally Isolated Phages, vB_Pae_SMP1 and vB_Pae_SMP5, Formulated as Hydrogels against Carbapenem-Resistant Pseudomonas aeruginosa. Viruses, 14(12), 2760. https://doi.org/10.3390/v14122760