

Genomic Epidemiology of SARS-CoV-2 in Western Burkina Faso, West Africa

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Ethical Approval
2.3. Sampling and SARS-CoV-2 Screening
2.4. Sequencing and Genome Assembly
2.5. Phylogenetic and Phylogeographic Analyses
3. Results
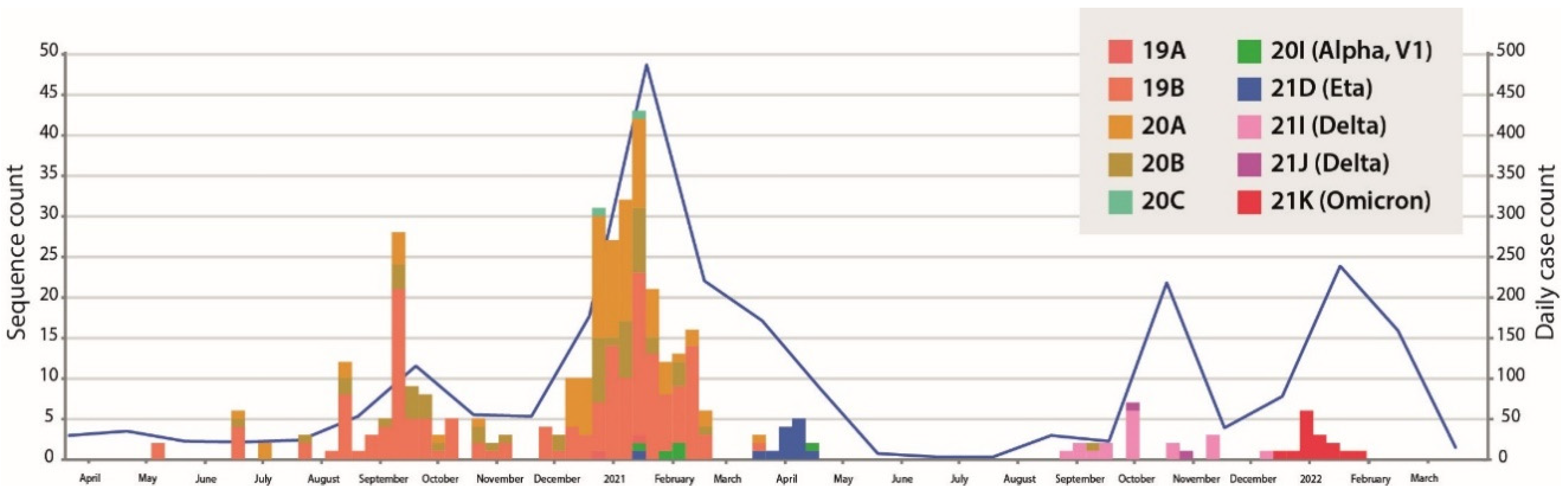
3.1. SARS-CoV-2 Genetic Diversity in Western Burkina Faso
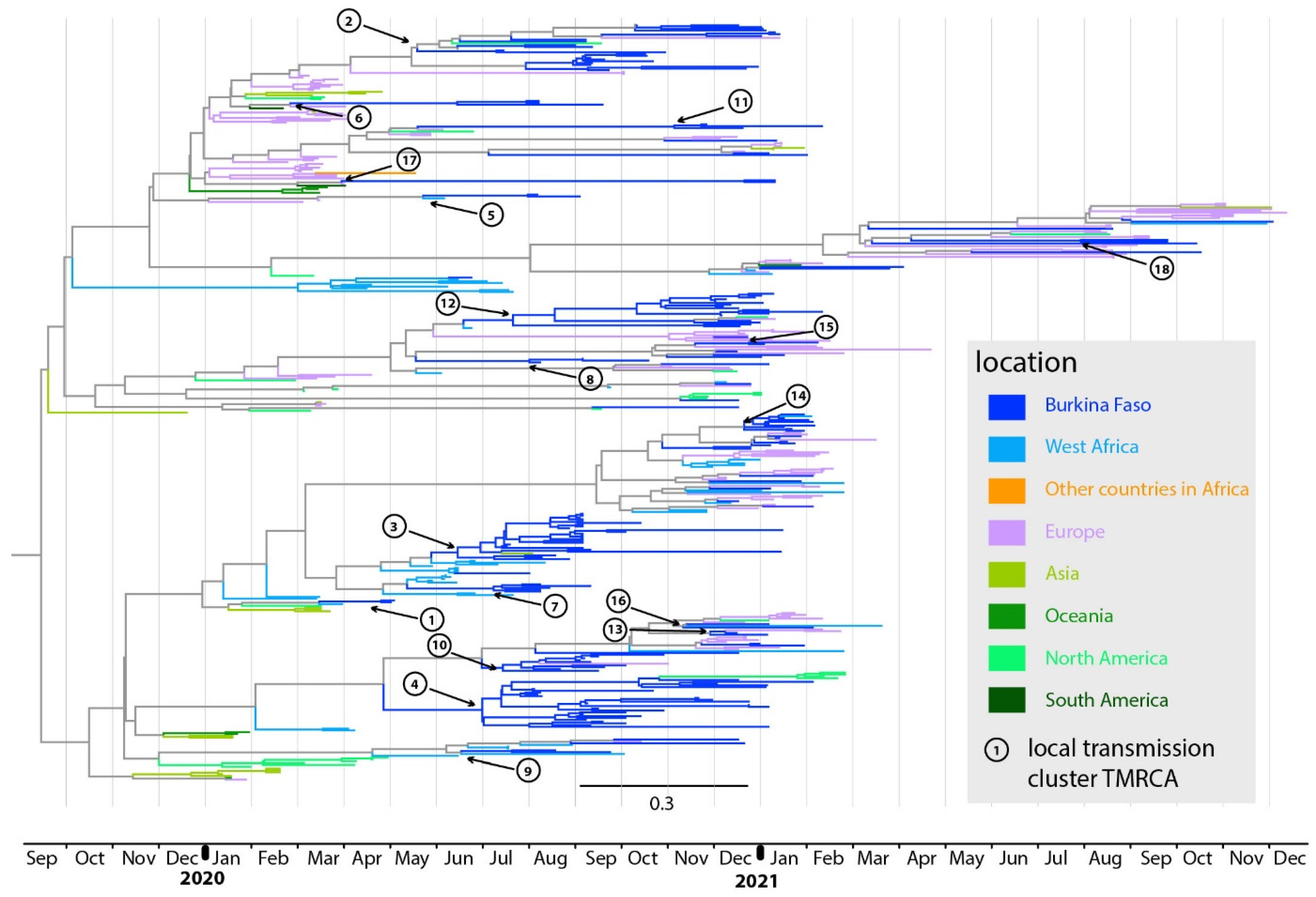
3.2. Geographic Sources of Virus Introduction into Western Burkina Faso
3.3. Timing of Virus Introductions into Western Burkina Faso
3.4. Geographic Distribution of Transmission Lineages in Western Burkina Faso
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Coronavirus Disease 2019 (COVID-19): Situation Report, 51; World Health Organization: Geneva, Switzerland, 2020. Available online: https://apps.who.int/iris/handle/10665/331475 (accessed on 6 December 2022).
- Nguendo-Yongsi, H.B.; Paka, E.E.; Makita-Ikouaya, E.; Niang-Diène, A.; Sop, M.S.; Bestandji, S.; Labii, B.; Zineddin, D.; Rouamba, J. Riposte Africaine à La Pandémie Du COVID-19: Une Urgence Sanitaire Pour Les Villes. Espace Géograph. Société Maroc. 2020, 40, 244–248. [Google Scholar]
- Lone, S.A.; Ahmad, A. COVID-19 Pandemic—An African Perspective. Emerg. Microbes Infect. 2020, 9, 1300–1308. [Google Scholar] [CrossRef] [PubMed]
- Plan de Préparation et de Riposte à l’épidemie de COVID-19 Au Burkina Faso Revisé|HumanitarianResponse. Available online: https://www.humanitarianresponse.info/en/op%C3%A9rations/burkina-faso/document/plan-de-pr%C3%A9paration-et-de-riposte-%C3%A0-l%C3%A9pidemie-de-covid-19-au (accessed on 4 May 2022).
- Burkina Faso—Décret N°2020-0271/PM/MDNAC/MATDC/MSECU/MS/MTMUSR Du 15 Avril 2020 Portant Restriction Temporaire de Libertés Au Titre Des Mesures Spéciales de Réduction de La Propagation Du COVID-19. Available online: http://www.droit-afrique.com/uploads/Burkina-Decret-2020-271-restriction-temporaire-liberte-pandemie.pdf (accessed on 6 December 2022).
- Réouverture des Frontières Aériennes: Le Premier Ministre Invite au Respect des Mesures Gouvernementales. Available online: https://www.gouvernement.gov.bf/informations/actualites/details?tx_news_pi1%5Baction%5D=detail&tx_news_pi1%5Bcontroller%5D=News&tx_news_pi1%5Bnews%5D=400&cHash=a67720b550e13a72aaef2351e817ffd0 (accessed on 4 May 2022).
- Décret Portant Réouverture Des Frontières Terrestres et Ferroviaires. Available online: https://www.sig.bf/wp-content/uploads/2021/11/D%C3%A9cret-portant-ouverture-des-fronti%C3%A8res-du-Burkina-Faso.pdf (accessed on 6 December 2022).
- Ritchie, H.; Mathieu, E.; Rodés-Guirao, L.; Appel, C.; Giattino, C.; Ortiz-Ospina, E.; Hasell, J.; Macdonald, B.; Beltekian, D.; Roser, M. Coronavirus Pandemic (COVID-19). Our World in Data 2020. Available online: https://ourworldindata.org/coronavirus (accessed on 20 October 2022).
- Shu, Y.; McCauley, J. GISAID: Global Initiative on Sharing All Influenza Data—From Vision to Reality. Eurosurveillance 2017, 22, 30494. [Google Scholar] [CrossRef] [Green Version]
- WHO COVID-19 Case Definition. Available online: https://www.who.int/publications-detail-redirect/WHO-2019-nCoV-Surveillance_Case_Definition-2022.1 (accessed on 20 October 2022).
- Quick, J. NCoV-2019 Sequencing Protocol v3 (LoCost). Available online: https://www.protocols.io/view/ncov-2019-sequencing-protocol-v3-locost-bh42j8ye (accessed on 20 October 2022).
- Nextstrain Clade Assignment. Available online: https://github.com/nextstrain/ncov/blob/9a1cf941872a421d7b69d98787c6aed73083e2cc/defaults/clades.tsv (accessed on 20 October 2022).
- Rambaut, A.; Holmes, E.C.; O’Toole, Á.; Hill, V.; McCrone, J.T.; Ruis, C.; du Plessis, L.; Pybus, O.G. A Dynamic Nomenclature Proposal for SARS-CoV-2 Lineages to Assist Genomic Epidemiology. Nat. Microbiol. 2020, 5, 1403–1407. [Google Scholar] [CrossRef]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.; Russell, T.W.; Tully, D.C.; Washburne, A.D. Estimated Transmissibility and Impact of SARS-CoV-2 Lineage B. 1.1. 7 in England. Science 2021, 372, eabg3055. [Google Scholar] [CrossRef]
- Davies, N.G.; Jarvis, C.I.; Edmunds, W.J.; Jewell, N.P.; Diaz-Ordaz, K.; Keogh, R.H. Increased Mortality in Community-Tested Cases of SARS-CoV-2 Lineage B. 1.1. 7. Nature 2021, 593, 270–274. [Google Scholar] [CrossRef]
- Funk, T.; Pharris, A.; Spiteri, G.; Bundle, N.; Melidou, A.; Carr, M.; Gonzalez, G.; Garcia-Leon, A.; Crispie, F.; O’Connor, L. Characteristics of SARS-CoV-2 Variants of Concern B. 1.1. 7, B. 1.351 or P. 1: Data from Seven EU/EEA Countries, Weeks 38/2020 to 10/2021. Eurosurveillance 2021, 26, 2100348. [Google Scholar] [CrossRef]
- Lemey, P.; Hong, S.L.; Hill, V.; Baele, G.; Poletto, C.; Colizza, V.; O’toole, Á.; McCrone, J.T.; Andersen, K.G.; Worobey, M. Accommodating Individual Travel History and Unsampled Diversity in Bayesian Phylogeographic Inference of SARS-CoV-2. Nat. Commun. 2020, 11, 5110. [Google Scholar] [CrossRef]
- Villabona-Arenas, C.; Hanage, W.P.; Tully, D.C. Phylogenetic Interpretation during Outbreaks Requires Caution. Nat. Microbiol. 2020, 5, 876–877. [Google Scholar] [CrossRef]
- Worobey, M.; Pekar, J.; Larsen, B.B.; Nelson, M.I.; Hill, V.; Joy, J.B.; Rambaut, A.; Suchard, M.A.; Wertheim, J.O.; Lemey, P. The Emergence of SARS-Cov-2 in Europe and North America. Science 2020, 370, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, E.; Giovanetti, M.; Tegally, H.; San, J.E.; Lessells, R.; Cuadros, D.; Martin, D.P.; Rasmussen, D.A.; Zekri, A.-R.N.; Sangare, A.K. A Year of Genomic Surveillance Reveals How the SARS-CoV-2 Pandemic Unfolded in Africa. Science 2021, 374, 423–431. [Google Scholar] [CrossRef] [PubMed]
- OCDE; ISSP. Interactions Entre Politiques Publiques, Migrations et Développement Au Burkina Faso; OCDE: Paris, France, 2017.
- Bugembe, D.L.; Kayiwa, J.; Phan, M.V.; Tushabe, P.; Balinandi, S.; Dhaala, B.; Lexow, J.; Mwebesa, H.; Aceng, J.; Kyobe, H. Main Routes of Entry and Genomic Diversity of SARS-CoV-2, Uganda. Emerg. Infect. Dis. 2020, 26, 2411. [Google Scholar] [CrossRef] [PubMed]
- Githinji, G.; de Laurent, Z.R.; Mohammed, K.S.; Omuoyo, D.O.; Macharia, P.M.; Morobe, J.M.; Otieno, E.; Kinyanjui, S.M.; Agweyu, A.; Maitha, E. Tracking the Introduction and Spread of SARS-CoV-2 in Coastal Kenya. Nat. Commun. 2021, 12, 4809. [Google Scholar] [CrossRef] [PubMed]
- Butera, Y.; Mukantwari, E.; Artesi, M.; O’Toole, Á.N.; Hill, V.; Rooke, S.; Hong, S.L.; Dellicour, S.; Majyambere, O.; Bontems, S. Genomic Sequencing of SARS-CoV-2 in Rwanda Reveals the Importance of Incoming Travelers on Lineage Diversity. Nature Commun. 2021, 12, 5705. [Google Scholar] [CrossRef]
- Chouikha, A.; Lagare, A.; Ghedira, K.; Diallo, A.; Njouom, R.; Sankhe, S.; Derrar, F.; Victoir, K.; Dellagi, K.; Triki, H. SARS-CoV-2 Lineage A. 27: New Data from African Countries and Dynamics in the Context of the COVID-19 Pandemic. Viruses 2022, 14, 1007. [Google Scholar] [CrossRef]
- Onwuamah, C.K.; Kanteh, A.; Abimbola, B.S.; Ahmed, R.A.; Okoli, C.L.; Shaibu, J.O.; James, A.B.; Ajibaye, O.; Okwuraiwe, A.P.; Fowora, M. SARS-CoV-2 Sequencing Collaboration in West Africa Shows Best Practices. Lancet Glob. Health 2021, 9, e1499–e1500. [Google Scholar] [CrossRef]
- Jain, M.; Olsen, H.E.; Paten, B.; Akeson, M. The Oxford Nanopore MinION: Delivery of Nanopore Sequencing to the Genomics Community. Genome Biol. 2016, 17, 239. [Google Scholar] [CrossRef] [Green Version]
- Quick, J.; Loman, N.J.; Duraffour, S.; Simpson, J.T.; Severi, E.; Cowley, L.; Bore, J.A.; Koundouno, R.; Dudas, G.; Mikhail, A. Real-Time, Portable Genome Sequencing for Ebola Surveillance. Nature 2016, 530, 228–232. [Google Scholar] [CrossRef] [Green Version]
- Quick, J.; Grubaugh, N.D.; Pullan, S.T.; Claro, I.M.; Smith, A.D.; Gangavarapu, K.; Oliveira, G.; Robles-Sikisaka, R.; Rogers, T.F.; Beutler, N.A. Multiplex PCR Method for MinION and Illumina Sequencing of Zika and Other Virus Genomes Directly from Clinical Samples. Nat. Protoc. 2017, 12, 1261–1276. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, W.; Bivins, A.; Smith, W.J.; Metcalfe, S.; Stephens, M.; Jennison, A.V.; Moore, F.A.; Bourke, J.; Schlebusch, S.; McMahon, J. Detection of the Omicron (B. 1.1. 529) Variant of SARS-CoV-2 in Aircraft Wastewater. Sci. Total Environ. 2022, 820, 153171. [Google Scholar] [CrossRef] [PubMed]
- Bull, R.A.; Adikari, T.N.; Ferguson, J.M.; Hammond, J.M.; Stevanovski, I.; Beukers, A.G.; Naing, Z.; Yeang, M.; Verich, A.; Gamaarachchi, H. Analytical Validity of Nanopore Sequencing for Rapid SARS-CoV-2 Genome Analysis. Nat. Commun. 2020, 11, 6272. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Transmission Cluster | Pango Lineage | Geographic Origin | TMRCA with the Most Likely Source Populations from Outside of the Country | Cluster Median TMRCA | Earliest Collection Date | Last Collection Date | Detection Lag (in Days) | Town |
|---|---|---|---|---|---|---|---|---|
| 1 (n = 2) | A | Unknown | ND | 1 May 2020 | 8 May 2020 | 10 May 2020 | ND | Bobo Dioulasso |
| 2 (n = 26) | B.1.1.404 | England | 8 March 2020 | 20 May 2020 | 21 July 2020 | 22 January 2021 | 73 | Bobo Dioulasso; Boromo; Gaoua; Houndé |
| 3 (n = 25) | A.19 | Cote d’Ivoire | 2 June 2020 | 20 June 2020 | 23 July 2020 | 20 January 2021 | 18 | Bobo Dioulasso; N’Dorola; Orodara; Diebougou |
| 4 (n = 27) | A.21 | Unknown | ND | 6 July 2020 | 10 August 2020 | 9 February 2021 | ND | Bobo Dioulasso; Orodara |
| 5 (n = 2) | B.1.388 | Unknown | ND | 5 August 2020 | 12 August 2020 | 9 September 2020 | ND | Bobo Dioulasso; Houndé |
| 6 (n = 3) | B.1.1 | England | 8 March 2020 | 11 July 2020 | 13 August 2020 | 24 September 2020 | 125 | Bobo Dioulasso; Houndé |
| 7 (n = 5) | A.19 | Cote d’Ivoire | 1 May 2020 | 13 July 2020 | 14 August 2020 | 16 September 2020 | 73 | Bobo Dioulasso |
| 8 (n = 6) | B.1 | Unknown | ND | 1 August 2020 | 14 August 2020 | 6 January 2021 | ND | Bobo Dioulasso; Banfora |
| 9 (n = 2) | A.18 | Cote d’Ivoire | 22 July 2020 | 25 July 2020 | 24 August 2020 | 29 September 2020 | 3 | Bobo Dioulasso |
| 10 (n = 10) | A.21 | Unknown | ND | 5 July 2020 | 27 August 2020 | 22 December 2020 | ND | Bobo Dioulasso |
| 11 (n = 3) | B.1.1 | England | 24 May 2020 | 9 November 2020 | 1 December 2020 | 15 February 2021 | 169 | Bobo Dioulasso |
| 12 (n = 17) | B.1 | Cote d’Ivoire | 23 June 2020 | 26 July 2020 | 7 December 2020 | 15 February 2021 | 33 | Bobo Dioulasso; Boromo |
| 13 (n = 2) | A.21 | Unknown | ND | 2 December 2020 | 21 December 2020 | 10 January 2021 | ND | Bobo Dioulasso |
| 14 (n = 16) | A.27 | Niger | 22 October 2020 | 1 November 2020 | 30 December 2020 | 10 February 2021 | 10 | Bobo Dioulasso |
| 15 (n = 2) | B.1 | Unknown | ND | 28 December 2020 | 8 January 2021 | 12 February 2021 | ND | Bobo Dioulasso; Batié |
| 16 (n = 2) | A.21 | Unknown | ND | 15 November 2020 | 11 January 2021 | 9 February 2021 | ND | Bobo Dioulasso |
| 17 (n = 2) | B.1.1.359 | Scotland | 4 April 2020 | 25 December 2020 | 15 January 2021 | 15 January 2021 | 265 | Bobo Dioulasso |
| 18 (n = 2) | AY.133 | Denmark | 14 March 2021 | 2 August 2021 | 29 september 2021 | 29 September 2021 | 141 | Bobo Dioulasso |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawadogo, Y.; Galal, L.; Belarbi, E.; Zongo, A.; Schubert, G.; Leendertz, F.; Kanteh, A.; Sesay, A.K.; Erhart, A.; Bañuls, A.-L.; et al. Genomic Epidemiology of SARS-CoV-2 in Western Burkina Faso, West Africa. Viruses 2022, 14, 2788. https://doi.org/10.3390/v14122788
Sawadogo Y, Galal L, Belarbi E, Zongo A, Schubert G, Leendertz F, Kanteh A, Sesay AK, Erhart A, Bañuls A-L, et al. Genomic Epidemiology of SARS-CoV-2 in Western Burkina Faso, West Africa. Viruses. 2022; 14(12):2788. https://doi.org/10.3390/v14122788
Chicago/Turabian StyleSawadogo, Yacouba, Lokman Galal, Essia Belarbi, Arsène Zongo, Grit Schubert, Fabian Leendertz, Abdoulie Kanteh, Abdul Karim Sesay, Annette Erhart, Anne-Laure Bañuls, and et al. 2022. "Genomic Epidemiology of SARS-CoV-2 in Western Burkina Faso, West Africa" Viruses 14, no. 12: 2788. https://doi.org/10.3390/v14122788
APA StyleSawadogo, Y., Galal, L., Belarbi, E., Zongo, A., Schubert, G., Leendertz, F., Kanteh, A., Sesay, A. K., Erhart, A., Bañuls, A.-L., Tarnagda, Z., Godreuil, S., Tinto, H., & Ouedraogo, A.-S. (2022). Genomic Epidemiology of SARS-CoV-2 in Western Burkina Faso, West Africa. Viruses, 14(12), 2788. https://doi.org/10.3390/v14122788