
Anti-Influenza Activity of Medicinal Material Extracts from Qinghai–Tibet Plateau

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of the Medicinal Plant, Lichen, and Mushroom Extracts
2.2. Cells
2.3. Virus
2.4. Cytotoxicity Studies of Extracts
2.5. In Vitro Microinhibition Assay
2.6. Hemagglutination Inhibition (HI) Test
3. Results
3.1. Characteristics of Extracts Obtained from Medicinal Plants, Lichens, and Fungi
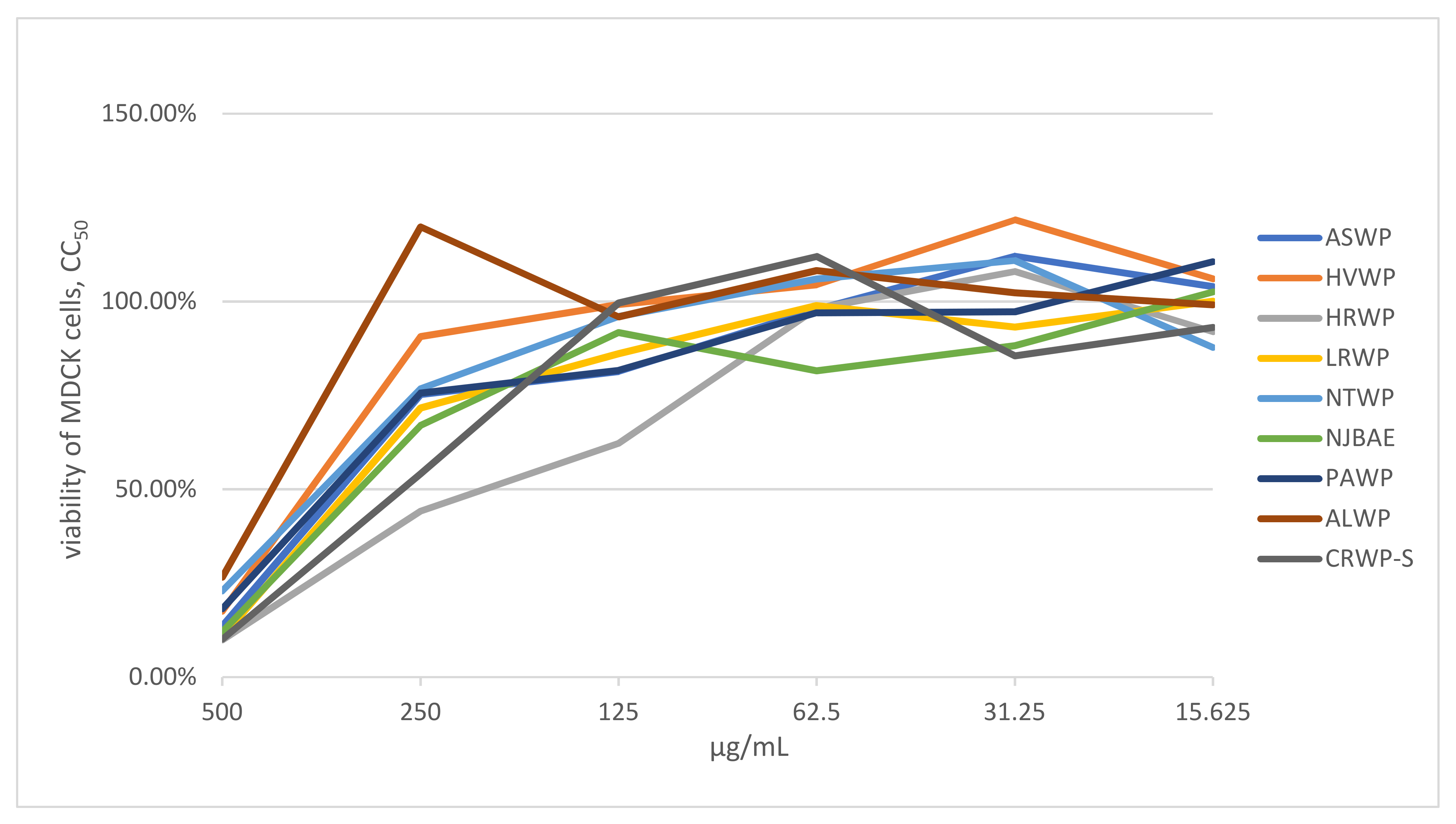
3.2. Cytotoxicity Studies of Medicinal Plant, Lichen, and Mushroom Extracts
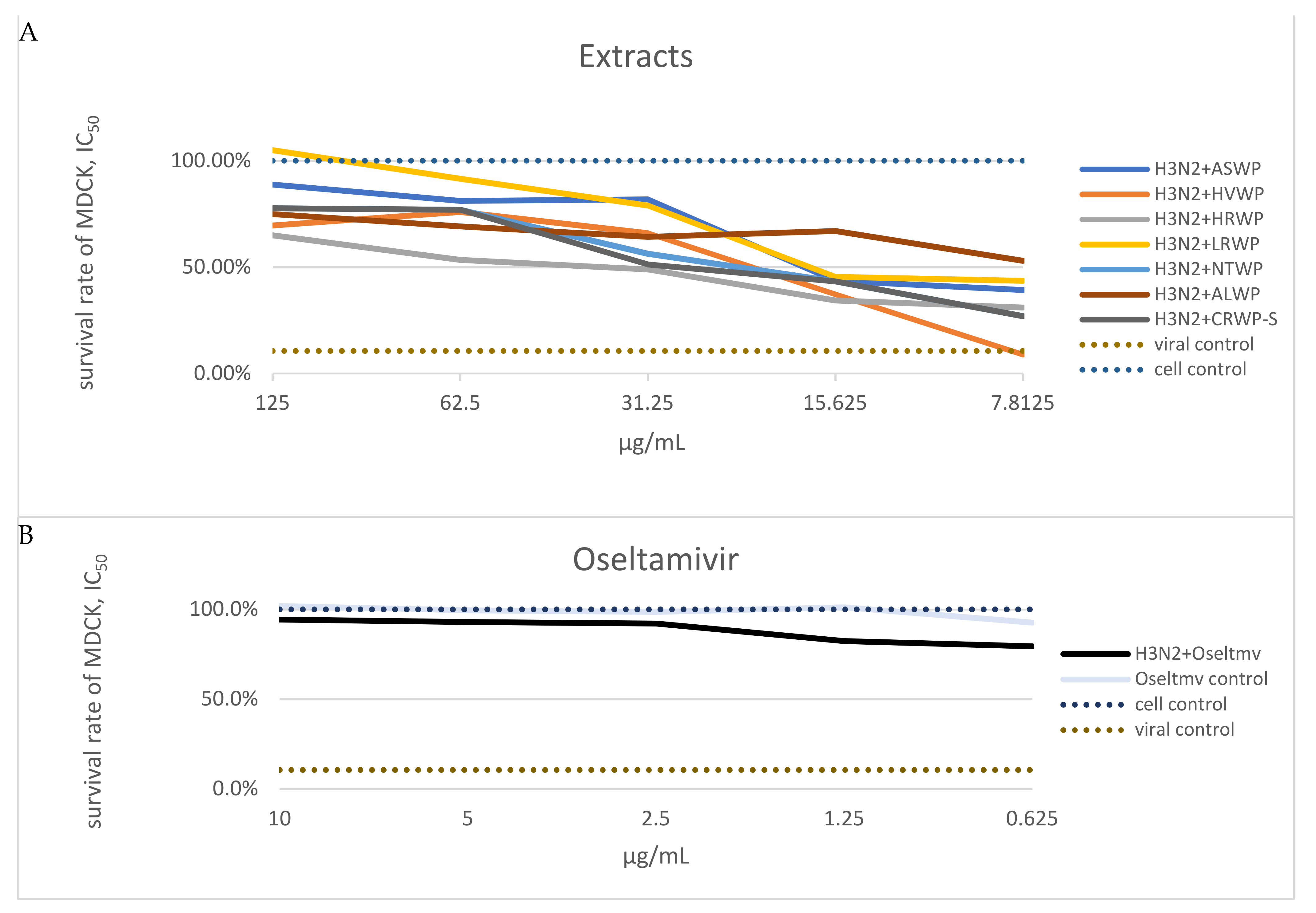
3.3. Inhibitory Effects of Plant, Lichen, and Mushroom Extracts on Influenza Virus
3.4. Inhibitory Effects of Medicinal Plant, Lichen, and Mushroom Extracts on Influenza-Virus-Induced Hemagglutination
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McCaughey, C. Influenza: A virus of our times. Ulst. Med. J. 2010, 79, 46–51. [Google Scholar]
- Biosciences, V. Delta NS1 Vaccine Platform. Available online: https://vivaldibiosciences.com/the-need (accessed on 18 December 2021).
- Ison, M. Antiviral Treatments. Clin. Chest Med. 2016, 38, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Yang, T. Baloxavir Marboxil: The First Cap-Dependent Endonuclease Inhibitor for the Treatment of Influenza. Ann. Pharmacother. 2019, 53, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.; Seong, B. Novel antiviral drug discovery strategies to tackle drug-resistant mutants of influenza virus strains. Expert Opin. Drug Discov. 2019, 14, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, D.; Palombo, E.; Yeo, T.; Ley, D.; Tu, C.; Malherbe, F.; Grollo, L. Identification of Traditional Medicinal Plant Extracts with Novel Anti-Influenza Activity. PLoS ONE 2013, 8, e79293. [Google Scholar] [CrossRef] [Green Version]
- Molinari, G. Natural products in drug discovery: Present status and perspectives. Adv. Exp. Med. Biol. 2009, 655, 13–27. [Google Scholar]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Sun, Y.; Si, J.; Liu, J.; Sun, G.; Sun, X.; Cao, L. Cryptoporus volvatus extract inhibits influenza virus replication in vitro and in vivo. PLoS ONE 2014, 9, e113604. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Tan, D.; Hsu, W.; Jong, T.; Wen, C.; Hsu, S.; Chang, P. Antiinfluenza virus activity of the ethanolic extract from Peperomia sui. J. Ethnopharmacol. 2014, 155, 320–325. [Google Scholar] [CrossRef]
- Zhou, H.; Mu, Z.; Du, X.; He, J.; Cao, L.; Zhong, G. Traightened on Chinese endemic seed plant species of medicine plants used in Tibetan medicine. China J. Chin. Mater. Med. 2015, 40, 3463–3469. [Google Scholar]
- Hao, D.; Xiao, P. Genomics and Evolution in Traditional Medicinal Plants: Road to a Healthier Life. Evol. Bioinform. 2015, 11, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Li, B. Investigation and study on traditional Tibetan medicine plant resources of Gramineae in eastern Qinghai-Tibet Plateau. Chin. Wild Plant Res. 2016, 5, 61–62. [Google Scholar]
- Dierma, D. Crystal Pearl of Materia Medica; Shanghai Scientific and Technical Publishers: Shanghai, China, 2012. [Google Scholar]
- Ni, W.; Gao, T.; Wang, H.; Du, Y.; Li, J.; Li, C.; Wei, L.; Bi, H. Anti-fatigue activity of polysaccharides from the fruits of four Tibetan plateau indigenous medicinal plants. J. Ethnopharmacol. 2013, 150, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Ma, J.; Bi, H.; Song, J.; Yang, H.; Xia, Z.; Du, Y.; Gao, T.; Wei, L. Characterization and cardioprotective activity of anthocyanins from Nitraria tangutorum Bobr. by-products. Food Funct. 2017, 8, 2771–2782. [Google Scholar] [CrossRef]
- Hu, T.; Shuai, X.; Chen, J.; Wei, Y.; Zheng, R. Protective effect of a Potentilla anserine polysaccharide on oxidative damages in mice. Int. J. Biol. Macromol. 2009, 45, 279–283. [Google Scholar] [CrossRef]
- Chen, J.; Yang, Z.; Hu, T.; Yan, Z.; Niu, T.; Wang, L.; Cui, D.; Wang, M. Immunomodulatory activity in vitro and in vivo of polysaccharide from Potentilla anserine. Fitoterapia 2010, 81, 1117–1124. [Google Scholar] [CrossRef]
- Hu, T.; Wei, X.; Zhang, X.; Cheng, F.; Shuai, X.; Zhang, L.; Kang, L. Protective effect of Potentilla anserine polysaccharide (PAP) on hydrogen peroxide induced apoptosis in murine splenic lymphocytes. Carbohydr. Polym. 2010, 79, 356–361. [Google Scholar] [CrossRef]
- Zhu, Y. Chinese Materia Medica (CMM); Shanghai: Shanghai Scientific and Technical Publishers: Shanghai, China, 1999. [Google Scholar]
- Huang, X.; Ma, J.; Wei, L.; Song, J.; Li, C.; Yang, H.; Du, Y.; Gao, T.; Bi, H. An antioxidant α-glucan from Cladina rangiferina (L.) Nyl. and its protective effect on alveolar epithelial cells from Pb2+-induced oxidative damage. Int. J. Biol. Macromol. 2018, 112, 101–109. [Google Scholar] [CrossRef]
- Yang, J.; Chen, Y.; Feng, X.; Yu, D.; He, C.; Zheng, Q.; Yang, J.; Liang, X. Isolation and structure elucidation of Armillaricin. Planta Med. 1989, 55, 564–565. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Shao, Y.; Tao, Y.; Wen, H. Optimization of dynamic microwave-assisted extraction of Armillaria polysaccharides using RSM, and their biological activity. LWT-Food Sci. Technol. 2015, 64, 1263–1269. [Google Scholar] [CrossRef]
- Gangopadhyay, N.; Hossain, M.; Rai, D.; Brunton, N. Optimisation of yield and molecular weight of b-glucan from barley flour using response surface methodology. J. Cereal Sci. 2015, 62, 38–44. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative-determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Sedmak, J.; Grossberg, S. Rapid, sensitive, and versatile assay forprotein using coomassie brilliant blue G250. Anal. Biochem. 1977, 79, 544–552. [Google Scholar] [CrossRef]
- Lavelli, V.; Sri-Harsha, P.; Spigno, G. Modelling the stability of maltodextrin-encapsulated grape skin phenolics used as a new ingredient in apple puree. Food Chem. 2016, 209, 323–331. [Google Scholar] [CrossRef]
- Sun, H.; Pan, H. Immunological adjuvant effect of Glycyrrhiza uralensis saponins on the immune responses to ovalbumin in mice. Vaccine 2006, 24, 1914–1920. [Google Scholar] [CrossRef] [PubMed]
- Groves, H.; Piché-Renaud, P.; Peci, A.; Farrar, D.; Buckrell, S.; Bancej, C.; Sevenhuysen, C.; Campigotto, A.; Gubbay, J.; Morris, S. The impact of the COVID-19 pandemic on influenza, respiratory syncytial virus, and other seasonal respiratory virus circulation in Canada: A population-based study. Lancet Reg. Health 2021, 1, 100015. [Google Scholar] [CrossRef]
- Olsen, S.; Winn, A.; Budd, A.; Prill, M.; Steel, J.; Midgley, C.; Kniss, K.; Burns, E.; Rowe, T.; Foust, A.; et al. Changes in Influenza and Other Respiratory Virus Activity during the COVID-19 Pandemic—United States, 2020–2021. CDC. Morbidity and Mortality Weekly Report (MMWR). Available online: https://www.cdc.gov/mmwr/volumes/70/wr/mm7029a1.htm#F1_down (accessed on 9 February 2022).
- Varela, F.; Scotta, M.; Polese-Bonatto, M.; Sartor, I.; Ferreira, C.; Fernandes, I.; Zavaglia, G.; Ferreira de Almeida, W.; Arakaki-Sanchez, D.; Pinto, L.; et al. Absence of detection of RSV and influenza during the COVID-19 pandemic in a Brazilian cohort: Likely role of lower transmission in the community. J. Glob. Health 2021, 11, 05007. [Google Scholar] [CrossRef]
- Xia, L.; Sun, Z.; Li, G.; Suo, Y.; You, J. A sensitive analytical method for the component monosaccharides of the polysaccharides from a Tibetan herb Potentilla anserine L. by capillary zone electrophoresis with UV detector. Eur. Food Res. Technol. 2010, 230, 715–722. [Google Scholar] [CrossRef]
- Wang, H.; Gao, T.; Du, Y.; Yang, H.; Wei, L.; Bi, H.; Ni, W. Anticancer and immunostimulating activities of a novel homogalacturonan from Hippophae rhamnoides L. berry. Carbohydr. Polym. 2015, 131, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M. Cereal beta-glucans: An underutilized health endorsing food ingredient. Crit. Rev. Food Sci. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Reynertson, K.; Garay, M.; Nebus, J.; Chon, S.; Kaur, S.; Mahmood, K.; Kizoulis, M.; Southall, M. Anti-inflammatory activities of colloidal oatmeal (Avena sativa) contribute to the effectiveness of oats in treatment of itch associated with dry, irritated skin. J. Drugs Dermatol. 2015, 14, 43–48. [Google Scholar]



{kind=link}
{kind=link}
| No. | Origin | Extract Name |
|---|---|---|
| 1 | Bran of Avena sativa L. | ASWP |
| 2 | Bran of Hordeum vulgare Linn. var. nudum Hook. f. | HVWP |
| 3 | Fruit of plant Hippophae rhamnoides Linn. | HRWP |
| 4 | Fruit of plant Lycium ruthenicum Murr. | LRWP |
| 5 | Fruit of plant Nitraria tangutorum Bobr. | NTWP |
| 6 | Anthocyanins from plant Nitraria tangutorum Bobr. by-products | NJBAE |
| 7 | Root of plant Potentilla anserina L. | PAWP |
| 8 | Dendritic part of lichen Cladina rangiferina (L.) Nyl. | CRWP-S |
| 9 | Fruit body of mushroom Armillaria luteo-virens (Aalb.et Schw:Fr) Sacc. | ALWP |
| No. | Extract Name | Total Sugar Content (%) | Uronic Acid Content (%) | Total Protein and Peptide Content (%) | Total Anthocyanin Content (mg CGE per g) | Total Endotoxin Content (EU)/mL | CC50 (μg/mL) | IC50 (μg/mL) |
|---|---|---|---|---|---|---|---|---|
| 1 | ASWP | 97.42 | n.d. | n.d. | n.d. | <0.5 | 395.46 | 19.53 ± 0.41 |
| 2 | HVWP | 93.20 | n.d. | n.d. | n.d. | <0.5 | 354.56 | 23.73 ± 0.62 |
| 3 | HRWP | 94.73 | 61.93 | 1.32 | 0.24 | <0.5 | 186.36 | 36.46 ± 1.12 |
| 4 | LRWP | 95.37 | 3.32 | 0.01 | 5.53 | <0.5 | 409.11 | 17.58 ± 0.38 |
| 5 | NTWP | 97.25 | 2.78 | 2.10 | 6.12 | <0.5 | 372.74 | 23.80 ± 0.38 |
| 6 | NJBAE | 5.52 | n.d. | n.d. | 723.60 | <0.5 | 286.37 | n.d. |
| 7 | PAWP | 69.35 | 9.51 | 0.07 | 6.79 | <0.5 | 386.38 | n.d. |
| 8 | ALWP | 51.43 | 0.08 | 10.22 | 7.33 | <0.5 | 440.90 | 7.81 ± 0.12 |
| 9 | CRWP-S | 92.16 | 0.01 | 2.92 | n.d. | <0.5 | 277.28 | 31.25 ± 2.51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurskaya, O.; Prokopyeva, E.; Bi, H.; Sobolev, I.; Murashkina, T.; Shestopalov, A.; Wei, L.; Sharshov, K. Anti-Influenza Activity of Medicinal Material Extracts from Qinghai–Tibet Plateau. Viruses 2022, 14, 360. https://doi.org/10.3390/v14020360
Kurskaya O, Prokopyeva E, Bi H, Sobolev I, Murashkina T, Shestopalov A, Wei L, Sharshov K. Anti-Influenza Activity of Medicinal Material Extracts from Qinghai–Tibet Plateau. Viruses. 2022; 14(2):360. https://doi.org/10.3390/v14020360
Chicago/Turabian StyleKurskaya, Olga, Elena Prokopyeva, Hongtao Bi, Ivan Sobolev, Tatyana Murashkina, Alexander Shestopalov, Lixin Wei, and Kirill Sharshov. 2022. "Anti-Influenza Activity of Medicinal Material Extracts from Qinghai–Tibet Plateau" Viruses 14, no. 2: 360. https://doi.org/10.3390/v14020360
APA StyleKurskaya, O., Prokopyeva, E., Bi, H., Sobolev, I., Murashkina, T., Shestopalov, A., Wei, L., & Sharshov, K. (2022). Anti-Influenza Activity of Medicinal Material Extracts from Qinghai–Tibet Plateau. Viruses, 14(2), 360. https://doi.org/10.3390/v14020360