
A Review of Viruses Infecting Yam (Dioscorea spp.)


Abstract
:1. Introduction
2. Family Potyviridae
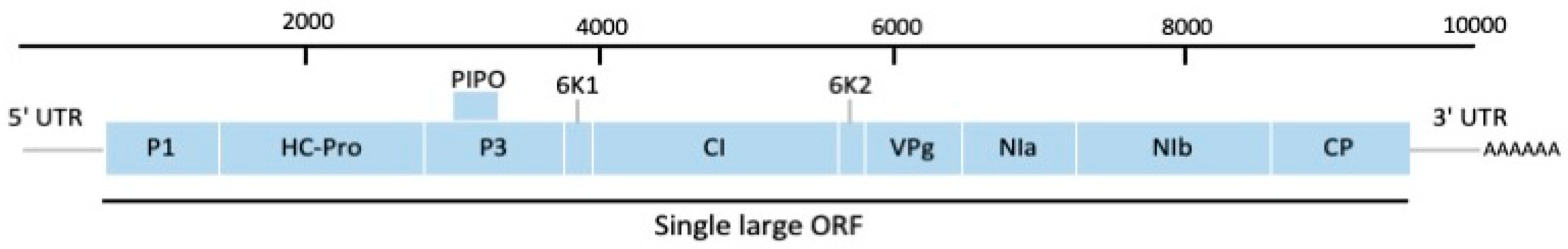
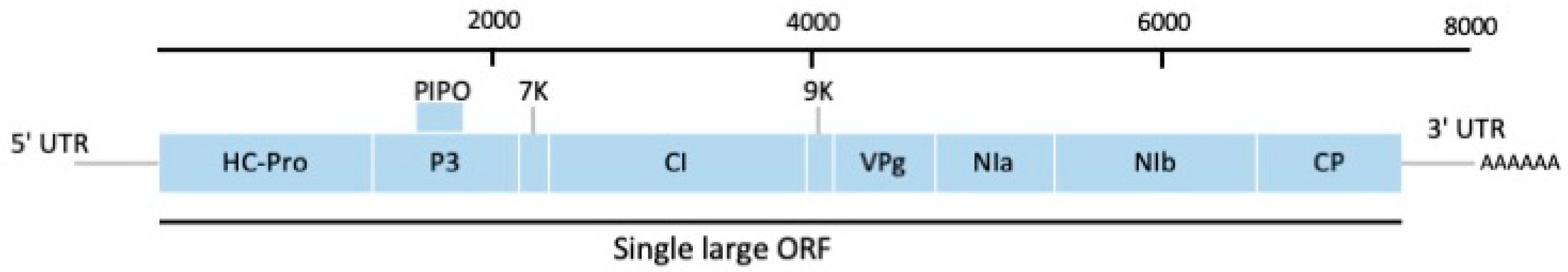
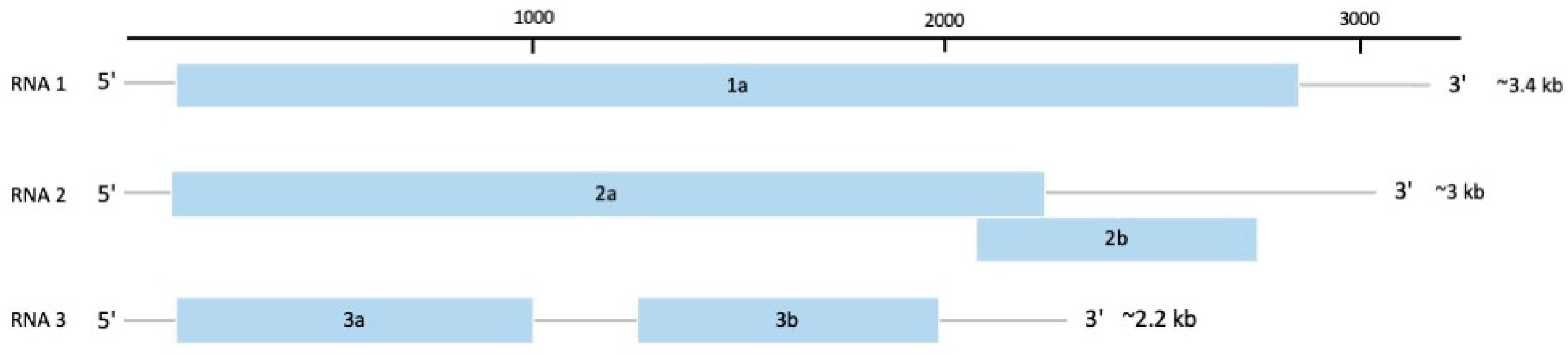
2.1. Genus Potyvirus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Name | Genome Organization | Primer Name | Gene Region/ORF | Primer Sequence (5′–3′) | Annealing Temperature (°C) | PCR Product Length (bp) | Reference |
|---|---|---|---|---|---|---|---|
| YMV | (+) ssRNA | YMV1 YMV2 | NIb, CP, 3′UTR | TGCGGAACTCRAAAGAAC TGCCATCAAATCCAAACA | 52 | 196 | [50] |
| YMV1 Poly T | NIb, CP, 3′UTR | TGCGGAACTCRAAAGAAC TTTTTTTTTTTTTTTTTTTTTTTTV | 52 | 450 | [50] | ||
| YMV CP 1F YMV UTR 1R | CP, 3′UTR | ATCCGGGATGTGGACAATGA TGGTCCTCCGCCACATCAAA | 55 | 586 | [52] | ||
| YMMV | (+) ssRNA | YMMV CP 2F YMMV UTR 1R | CP, 3′UTR | GGCACACATGCAAATGAAAGC CACCAGTAGAGTGAACATAG | 55 | 249 | [52] |
| YMMVCP-Bam FP YMMV-CP-EcoRP | CP | CAGAGAGGATCCGCAAGTAAGGAACAGACATTTG TTGATCGAATTCCTAGATATTGCGCACTCCAAGAAG | 55 | 798 | [54] | ||
| Macluraviruses | (+) ssRNA | YamMac4F YamMac5R | CP | CHGCAGCWATYGGKMGTG GGGTTGCTGAGCRTYGGA | 47 | 292 | [19] |
| CYNMV | (+) ssRNA | CYNMV-Det-Fw CYNMV-Det-Rv | 3′ proximal region | GTGTGCTAACAATGGTACATCATC GTGCGTTGAGGGTTGCTGAGC | 55 | 609 | [58] |
| YCNV | (+) ssRNA | YCNF504 YCNR1269 | 5′ region | TTAACGTCATAGTAGCTCTCAGC TAGTCGAGCTGAACCCTAACCAC | - | - | [12] |
| Badnaviruses | Circular dsDNA | BadnaFP BadnaRP | RT, RnaseH | ATGCCITTYGGIITIAARAAYGCICC CCAYTTRCAIACISCICCCCAICC | 49 | 579 | [59] |
| CMV | (+) ssRNA | CMV1-F CMV1-R | 3′UTR, CP | GTAGACATCTGTGACGCGA GCGCGAAACAAGCTTCTTATC | 55 | 540 | [60] |
| CMV primer 1 CMV primer 2 | CP | GCCGTAAGCTGGATGGACAA TATGATAAGAAGCTTGTTTCGCG | 55 | 500 | [61] | ||
| CMV-F IITA CMV-R | Replicase | GCCGTAAGCTGGATGGACAA CCGCTTGTGCGTTTAATGGCT | 54 | 520 | [1] | ||
| Potexviruses | (+) ssRNA | Potex-2RC Potex-5 | ORF1 | AGCATRGCNSCRTCYTG CAYCARCARGCMAARGAYGA | 49 | 584 | [62] |
| YamX-3F yamX-5R | RdRp domain | CICARTGGGTIAAGAAARWKGA GRTCRAAIGCIGTRAARTCATTIGC | 43 | 255 | [19] | ||
| YaV1 | (+) ssRNA | DiosClos-F DiosClos-R | 3′end ORF8, 3′NCR | CTCTTTAGGTTTCCCATTTATCA TGGTTCTACATTACTAGACTAC | 49 | 285 | [63] |
| DMaV | (+) ssRNA | Seco-1F Seco-1R | RdRp | AACTCCWTCWGGWTTYGCTYTGAC CCCACTTYCTY TGAGAAAATCAA | 46 | 323 | [19] |
| YVY | (+) ssRNA | YVY-RdRp1-PF YVY-RdRp1-PR | RdRp | GTAATTGAAAATCACAGTGAGC CTTCAAGTGCATAATTGTCTAT | 55 | 790 | [20] |
| YVY-CP-F YVY-CP-R | CP, 3′UTR | TTGATTAGTTAAGTATTTAGC CCAGTTTTTCCTGCTGGCAAAC | 55 | 788 | [20] | ||
| DNUaV | Circular dsDNA | DNUaV-ORF4-FP1 DNUaV-ORF4-RP1 | ORF4 sequence | CCGGGTTGCCAGTACAGAAT CGTGAAGCACCCAAACCTTG | 57 | 450 | [18] |
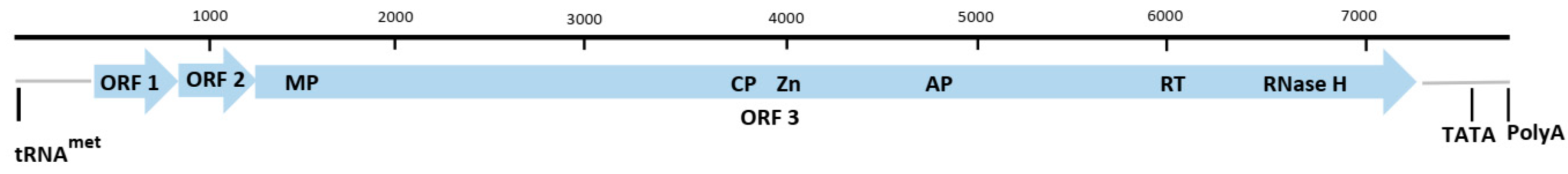
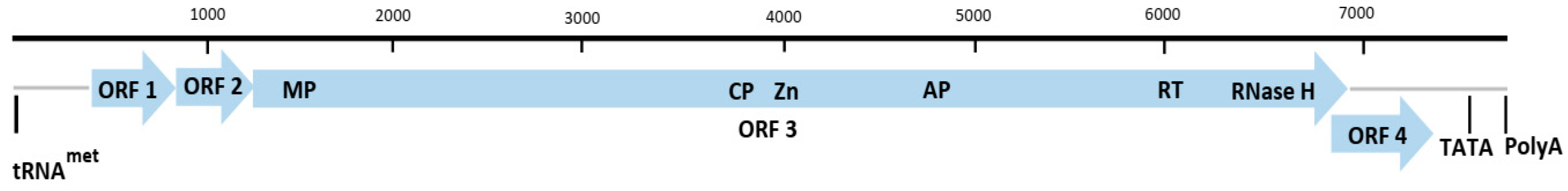
2.2. Genus Macluravirus
3. Family Caulimoviridae
4. Family Bromoviridae, Genus Cucumovirus
5. New Yam Viruses
6. Yam Virus Sanitation
7. Yam Virus Status in the Pacific
8. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azeteh, I.N.; Hanna, R.; Njukeng, A.P.; Oresanya, A.O.; Sakwe, P.N.; Kumar, P.L. Distribution and diversity of viruses infecting yams (Dioscorea spp.) in Cameroon. Virus Dis. 2019, 30, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Ita, E.E.; Uyoh, E.A.; Nakamura, I.; Ntui, V.O. Efficient elimination of Yam mosaic virus (YMV) from white yam (Dioscorea rotundata Poir.) by cryotherapy of axillary buds. South Afr. J. Bot. 2020, 130, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Tamiru, M.; Natsume, S.; Takagi, H.; White, B.; Yaegashi, H.; Shimizu, M.; Yoshida, K.; Uemura, A.; Oikawa, K.; Abe, A.; et al. Genome sequencing of the staple food crop white Guinea yam enables the development of a molecular marker for sex determination. BMC Biol. 2017, 15, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tostain, S.; Agbangla, C.; Scarcelli, N.; Mariac, C.; Dainou, O.; Berthaud, J.; Pham, J.-L. Genetic diversity analysis of yam cultivars (Dioscorea rotundata Poir.) in Benin using simple sequence repeat (SSR) markers. Plant Genet. Resour. Charact. Util. 2007, 5, 71–81. [Google Scholar] [CrossRef]
- Wang, M.; Li, F.; Zhou, G.; Lan, P.; Xu, D.; Li, R. Molecular Detection and characterization of Chinese yam mild mosaic virus isolates. J. Phytopathol. 2014, 163, 1036–1040. [Google Scholar] [CrossRef]
- Adenji, M.O.; Shoyinka, S.A.; Ikotun, T.; Asiedu, R.; Hughes, J.D.; Odu, B.O. Yield loss in guinea yam (Dioscorea rotundata Poir.) due to infection by YMV genus Potyvirus. Ife J. Sci. 2012, 14, 237–244. [Google Scholar]
- Amusa, N.; Adigbite, A.; Muhammed, S.; Baiyewu, R. Yam diseases and its management in Nigeria. Afr. J. Biotechnol. 2003, 2, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Lebot, V.; Malapa, R.; Abraham, K. The Pacific yam (Dioscorea nummularia Lam.), an under-exploited tuber crop from Melanesia. Genet. Resour. Crop Evol. 2017, 64, 217–235. [Google Scholar] [CrossRef]
- Guimarães, K.M.C.; Silva, S.J.C.; Melo, A.M.; Ramos-Sobrinho, R.; Lima, J.S.; Zerbini, F.M.; Assunção, I.P.; Lima, G.S.A. Genetic variability of badnaviruses infecting yam (Dioscorea spp.) in northeastern Brazil. Trop. Plant Pathol. 2015, 40, 111–118. [Google Scholar] [CrossRef]
- Kondo, T.; Fujita, T. Complete nucleotide sequence and construction of an infectious clone of Chinese yam necrotic mosaic virus suggest that macluraviruses have the smallest genome among members of the family Potyviridae. Arch. Virol. 2012, 157, 2299–2307. [Google Scholar] [CrossRef]
- Florence, M.; Ngo, S.; Omokolo, D.N.; Joly, S. Evolution and Phylogenetic Diversity of Yam Species (Dioscorea spp.): Implication for Conservation and Agricultural Practices. PLoS ONE 2015, 10, e0145364. [Google Scholar] [CrossRef] [Green Version]
- Lan, P.; Meng, Y.; Shen, P.; Li, R.; Ma, Y.; Tan, S.; Chen, H.; Cao, M.; Li, F. Complete genome sequence of yam chlorotic necrosis virus, a novel macluravirus infecting yam. Arch. Virol. 2018, 163, 2275–2278. [Google Scholar] [CrossRef] [PubMed]
- Mambole, I.A.; Bonheur, L.; Dumas, L.S.; Filloux, D.; Gomez, R.M.; Faure, C.; Lange, D.; Anzala, F.; Pavis, C.; Marais, A.; et al. Molecular characterization of yam virus X, a new potexvirus infecting yams (Dioscorea spp.) and evidence for the existence of at least three distinct potexviruses infecting yams. Arch. Virol. 2014, 159, 3421–3426. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QV (accessed on 15 August 2021).
- Yeyeh, T.; Marie, N.; Diallo, H.A.; Akinbade, S.A.; Séka, K.; Kumar, P.L. Distribution, incidence and severity of viral diseases of yam (Dioscorea spp.) in Côte d’Ivoire. Afr. J. Biotechnol. 2014, 13, 465–470. [Google Scholar] [CrossRef]
- Njukeng, A.P.; Azeteh, I.N.; Mbong, G.A. Article Survey of the incidence and distribution of two viruses infecting yam (Dioscorea spp.) in two agro-ecological zones of Cameroon. Int. J. Curr. Microbiol. App. Sci 2014, 3, 1153–1166. [Google Scholar]
- Kenyon, L.; Lebas, B.S.M.; Seal, S.E. Yams (Dioscorea spp.) from the South Pacific Islands contain many novel badnaviruses: Implications for international movement of yam germplasm. Arch. Virol. 2008, 153, 877–889. [Google Scholar] [CrossRef]
- Sukal, A.C.; Kidanemariam, D.B.; Dale, J.L.; Harding, R.M.; James, A.P. Characterization of a novel member of the family Caulimoviridae infecting Dioscorea nummularia in the pacific, which may represent a new genus of dsDNA plant viruses. PLoS ONE 2018, 13, e0203038. [Google Scholar] [CrossRef]
- Umber, M.; Filloux, D.; Gélabale, S.; Gomez, R.M.; Marais, A.; Gallet, S.; Gamiette, F.; Pavis, C.; Teycheney, P.Y. Molecular viral diagnosis and sanitation of yam genetic resources: Implications for safe yam germplasm exchange. Viruses 2020, 12, 1101. [Google Scholar] [CrossRef]
- Silva, G.; Bömer, M.; Rathnayake, A.I.; Sewe, S.O.; Visendi, P.; Oyekanmi, J.O.; Quain, M.D.; Akomeah, B.; Kumar, P.L.; Seal, S.E. Molecular characterization of a new virus species identified in yam (Dioscorea spp.) by high-throughput sequencing. Plants 2019, 8, 167. [Google Scholar] [CrossRef] [Green Version]
- Eni, A.O.; Kumar, P.L.; Asiedu, R.; Alabi, O.J.; Naidu, R.A.; Hughes, J.A. Characterization of cucumber mosaic virus isolated from yam (Dioscorea spp.) in West Africa. Afr. J. Biotechnol. 2013, 12, 3472–3480. [Google Scholar] [CrossRef]
- Odedara, O.O.; Ayo-john, E.I.; Gbuyiro, M.M.; Falade, F.O.; Agbebi, S.E. Serological detection of yam viruses in farmers’ fields in Ogun State, Nigeria. Arch. Phytopathol. Plant Prot. 2012, 45, 840–845. [Google Scholar] [CrossRef]
- Odu, B.O.; Asiedu, R.; Hughes, J.A.; Shoyinka, S.A.; Oladiran, O.A. Identification of resistance to Yam mosaic virus (YMV), genus Potyvirus in white Guinea yam (Dioscorea rotundata Poir.). Field Crop. Res. 2004, 89, 97–105. [Google Scholar] [CrossRef]
- Silva, G.; Bömer, M.; Nkere, C.; Lava Kumar, P.; Seal, S.E. Rapid and specific detection of yam mosaic virus by reverse-transcription recombinase polymerase amplification. J. Virol. Methods 2015, 222, 138–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakayoko, Y.; Kouakou, A.M.; Kouassi, A.B.; Gomez, R.; Dibi, K.E.B.B.; Essis, B.S.; N’Zué, B.; Adebola, P.; N’Guetta, A.S.P.; Umber, M. Detection and diversity of viruses infecting African yam (Dioscorea rotundata) in a collection and F 1 progenies in Côte d’Ivoire shed light to plant-to-plant viral transmission. Plant Pathol. 2021, 70, 1486–1495. [Google Scholar] [CrossRef]
- Thouvenel, J.C.; Fauquet, C. Yam mosaic, a new potyvirus infecting Dioscorea cayenensis in the Ivory Coast. Ann. Appl. Biol. 1979, 93, 279–283. [Google Scholar] [CrossRef]
- Odu, B.O.; Hughes, J.D.; Shoyinka, S.A.; Dongo, L.N. Isolation, characterization and identification of a potyvirus from Dioscorea alata L. (water yam) in Nigeria. Ann. Appl. Biol. 1999, 134, 65–71. [Google Scholar] [CrossRef]
- Phillips, S.; Briddon, R.W.; Brunt, A.A.; Hull, R. The Partial characterization of a badnavirus infecting the greater asiatic or water yam (Dioscorea alata). J. Phytopathol. 1999, 147, 265–269. [Google Scholar] [CrossRef]
- Bömer, M.; Rathnayake, A.I.; Visendi, P.; Silva, G.; Seal, S.E. Complete genome sequence of a new member of the genus Badnavirus, Dioscorea bacilliform RT virus 3, reveals the first evidence of recombination in yam badnaviruses. Arch. Virol. 2018, 163, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Filho, F.d.A.C.R.; Nicolini, C.; Resende, R.d.O.; de Andrade, G.P.; Pio-Ribeiro, G.; Nagata, T. The complete genome sequence of a Brazilian isolate of yam mild mosaic virus. Arch. Virol. 2013, 158, 515–518. [Google Scholar] [CrossRef]
- Fuji, S.; Nakamae, H. Complete nucleotide sequence of the genomic RNA of a Japanese yam mosaic virus, a new potyvirus in Japan. Arch. Virol. 1999, 144, 231–240. [Google Scholar] [CrossRef]
- Fuji, S.; Mitobe, I.; Nakamae, H.; Natsuaki, K.T. Nucleotide sequence of coat protein gene of yam mild mosaic virus, isolated in Papua New Guinea. Arch. Virol. 1999, 144, 1415–1419. [Google Scholar] [CrossRef] [PubMed]
- Nkere, C.K.; Oyekanmi, J.O.; Silva, G.; Bömer, M.; Atiri, G.I.; Onyeka, J.; Maroya, N.G.; Seal, S.E.; Kumar, P.L. Chromogenic detection of yam mosaic virus by closed-tube reverse transcription loop-mediated isothermal amplification (CT-RT-LAMP). Arch. Virol. 2018, 163, 1057–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bousalem, M.; Dallot, S.; Fuji, S.; Natsuaki, K.T. Origin, worldwide dispersion, bio-geographical diversification, radiation and recombination: An evolutionary history of Yam mild mosaic virus (YMMV). Infect. Genet. Evol. 2003, 3, 189–206. [Google Scholar] [CrossRef]
- Seal, S.; Turaki, A.; Muller, E.; Kumar, P.L.; Kenyon, L.; Filloux, D.; Galzi, S.; Lopez-montes, A.; Iskra-caruana, M. The prevalence of badnaviruses in West African yams (Dioscorea cayenensis-rotundata) and evidence of endogenous pararetrovirus sequences in thier genomes. Virus Res. 2014, 186, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Wylie, S.J.; Adams, M.; Chalam, C.; Kreuze, J.; Jos, J.; Ohshima, K.; Praveen, S.; Rabenstein, F.; Stenger, D.; Wang, A.; et al. ICTV Virus Taxonomy Profile: Potyviridae. J. Gen. Virol. 2017, 352–354. [Google Scholar] [CrossRef]
- Chung, B.Y.W.; Miller, W.A.; Atkins, J.F.; Firth, A.E. An overlapping essential gene in the Potyviridae. Proc. Natl. Acad. Sci. USA 2008, 105, 5897–5902. [Google Scholar] [CrossRef] [Green Version]
- Gadhave, K.R.; Gautam, S.; Rasmussen, D.A.; Srinivasan, R. Aphid transmission of potyvirus: The largest plant-infecting RNA virus genus. Viruses 2020, 12, 733. [Google Scholar] [CrossRef]
- Rybicki, E.P. A Top Ten list for economically important plant viruses. Arch. Virol. 2015, 160, 17–20. [Google Scholar] [CrossRef]
- Nigam, D.; LaTourrette, K.; Souza, P.F.N.; Garcia-Ruiz, H. Genome-wide variation in Potyviruses. Front. Plant Sci. 2019, 10, 1439. [Google Scholar] [CrossRef] [Green Version]
- Ayisah, K.D.; Gumedzoe, Y.M.D. Genetic diversity among yam mosaic virus (YMV) isolates infecting yam of the complex Dioscorea cayenensis-rotundata in Togo. Int. J. Biol. Chem. Sci. 2012, 6, 1090–1101. [Google Scholar] [CrossRef]
- SPYN. Yam: Cultivar Selection for Disease Resistance & Commercial Potential in Pacific Islands; CIRAD: Montpellier, France, 2003; pp. 68–88. [Google Scholar]
- Aleman, M.E.; Marcos, J.F.; Brugidou, C.; Beachy, R.N.; Fauquet, C. The complete nucleotide sequence of yam mosaic virus (Ivory Coast isolate) genomic RNA. Arch. Virol. 1996, 141, 1259–1278. [Google Scholar] [CrossRef] [PubMed]
- Bömer, M.; Rathnayake, A.I.; Visendi, P.; Sewe, S.O.; Paolo, J.; Sicat, A.; Silva, G.; Kumar, P.L.; Seal, S.E. Tissue culture and next-generation sequencing: A combined approach for detecting yam (Dioscorea spp.) viruses. Physiol. Mol. Plant Pathol. 2019, 105, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Aleman-Verdaguer, M.E.; Goudou-Urbino, C.; Dubern, J.; Beachy, R.N.; Fauquet, C. Analysis of the sequence diversity of the P1, HC, P3, NIb and CP genomic regions of several yam mosaic potyvirus isolates: Implications for the intraspecies molecular diversity of potyviruses. J. Gen. Virol. 1997, 78, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Mignouna, H.D.; Abang, M.M.; Onasanya, A.; Agindotan, B.; Asiedu, R.; Iita, A.; Lambourn, L.W. Identification and potential use of RAPD markers linked to yam mosaic virus resistance in white yam (Dioscorea rotundata). Ann. Appl. Biol. 2002, 140, 163–169. [Google Scholar] [CrossRef]
- Odu, B.O.; Asiedu, R.; Shoyinka, S.A.; Hughes, J.D. Analysis of resistance to yam mosaic virus, genus Potyvirus in white Guinea yam (Dioscorea rotundata Poir.) genotypes. J. Agric. Sci. 2011, 56, 1–13. [Google Scholar] [CrossRef]
- Goudou-Urbino, C.; Givord, L.; Quiot, J.B.; Boeglin, M.; Konate, G.; Dubern, J. Differentiation of yam virus isolates using symptomatology, western blot assay, and monoclonal antibodies. J. Phytopathol. 1996, 144, 235–240. [Google Scholar] [CrossRef]
- Bousalem, M.; Douzery, E.J.P.; Fargette, D. High genetic diversity, distant phylogenetic relationships and intraspecies recombination events among natural populations of yam mosaic virus: A contribution to understanding potyvirus evolution. J. Gen. Virol. 2000, 81, 243–255. [Google Scholar] [CrossRef]
- Bousalem, M.; Dallot, S.; Guyader, S. The use of phylogenetic data to develop molecular tools for the detection and genotyping of yam mosaic virus. Potential application in molecular epidemiology. J. Virol. Methods 2000, 90, 25–36. [Google Scholar] [CrossRef]
- Mignouna, H.D.; Njuken, P.; Abang, M.; Asiedu, R. Inheritance of resistance to yam mosaic virus, genus Potyvirus, in white yam (Dioscorea rotundata). Theor. Appl. Genet. 2001, 103, 1196–2000. [Google Scholar] [CrossRef]
- Mumford, R.A.; Seal, S.E. Rapid single-tube immunocapture RT-PCR for the detection of two yam potyviruses. J. Virol. Methods 1997, 69, 73–79. [Google Scholar] [CrossRef]
- Eni, A.O.; Asiedu, R. Re-evaluation of yam mosaic virus (YMV) detection methods. Acad. J. Plant Sci. 2012, 5, 18–22. [Google Scholar] [CrossRef]
- Nkere, C.K.; Otoo, E.; Atiri, G.I.; Onyeka, J.; Silva, G.; Bömer, M.; Seal, S.E.; Kumar, P.L. Assessment of yam mild mosaic virus coat protein gene sequence diversity reveals the prevalence of cosmopolitan and African group of isolates in Ghana and Nigeria. Curr. Plant Biol. 2020, 23, 100156. [Google Scholar] [CrossRef]
- Oppong, A.; Lamptey, J.N.L.; Ofori, F.A.; Anno-Nyako, F.O.; Offei, S.K.; Dzomeku, B.M. Serological detection of Dioscorea alata potyvirus on white yams (Dioscorea rotundata) in Ghana. J. Plant Sci. 2007, 2, 630–634. [Google Scholar] [CrossRef]
- Zou, C.-W.; Meng, J.-R.; Yao, Z.-T.; Zhang, L.; Wang, Z.-Q.; Wei, B.-H.; Chen, B.-S. Genetic diversity and genome recombination in yam mild mosaic virus isolates. Phytopathol. Res. 2020, 2, 10. [Google Scholar] [CrossRef]
- Eni, A.O.; Hughes, J.D.A.; Rey, M.E.C. Survey of the incidence and distribution of five viruses infecting yams in the major yam-producing zones in Benin. Ann. Appl. Biol. 2008, 153, 223–232. [Google Scholar] [CrossRef]
- Kwon, S.; Cho, I.; Choi, S.; Yoon, J.; Choi, G. Complete sequence analysis of a Korean isolate of Chinese yam necrotic mosaic virus and generation of the virus-specific primers for molecular detection. Res. Plant Dis. 2016, 22, 194–197. [Google Scholar] [CrossRef] [Green Version]
- Yang, I.C.; Hafner, G.J.; Dale, J.L.; Harding, R.M. Genomic characterization of taro bacilliform virus. Arch. Virol. 2003, 148, 937–949. [Google Scholar] [CrossRef]
- De Blas, C.; Borja, M.J.; Saiz, M.; Romero, J. Broad-spectrum detection of cucumber mosaic virus (CMV) using the polymerase chain reaction. J. Phytopathol. 1994, 141, 323–329. [Google Scholar] [CrossRef]
- Wylie, S.; Wilson, C.R.; Jones, R.A.C.; Jones, M.G.K. A polymerase chain reaction assay for cucumber mosaic virus in lupin seeds. Aust. J. Agric. Res. 1993, 44, 41–51. [Google Scholar] [CrossRef]
- Van der Vlugt, R.A.A.; Berendsen, M. Development of a general potexvirus detection method. Eur. J. Plant Pathol. 2002, 108, 367–371. [Google Scholar] [CrossRef]
- Marais, A.; Umber, M.; Filloux, D.; Gomez, R.-M.; Faure, C.; Pavis, C.; Julian, C.; Roumagnac, P.; Acina-Mambole, I.; Bonheur, L.; et al. Yam asymptomatic virus 1, a novel virus infecting yams (Dioscorea spp.) with significant prevalence in a germplasm collection. Arch. Virol. 2020, 165, 2653–2657. [Google Scholar] [CrossRef] [PubMed]
- Lan, P.; Li, F.; Wang, M.; Li, R. Complete genome sequence of a divergent strain of Japanese yam mosaic virus from China. Arch. Virol. 2014, 160, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Son, C.-G.; Kwon, J.-B.; Nam, H.-H.; Kim, Y.-T.; Kim, M.K.; Lee, S.-H. Diagnosis and sequence analysis of Japanese yam mosaic virus from yam (Dioscorea opposita). Res. Plant Dis. 2016, 22, 289–292. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki, T.; Iwamoto, I.; Atarashi, A.; Ohki, S.T.; Kikukawa, H. Rapid and low-cost diagnosis of Japanese yam mosaic virus infection in Chinese yam (Dioscorea polystachya) leaves by a print-capture RT-PCR. J. Gen. Plant Pathol. 2017, 83, 189–196. [Google Scholar] [CrossRef]
- Fuji, S.; Nakamae, H. Complete nucleotide sequence of the genomic RNA of a mild strain of Japanese yam mosaic potyvirus. Arch. Virol. 2000, 145, 635–640. [Google Scholar] [CrossRef]
- Fukuta, S.; Iida, T.; Mizukami, Y.; Ishida, A.; Ueda, J.; Kanbe, M.; Ishimoto, Y. Detection of Japanese yam mosaic virus by RT-LAMP. Arch. Virol. 2003, 148, 1713–1720. [Google Scholar] [CrossRef]
- Wang, H.; Qi, M.; Cutler, A.J. A simple method of preparing plant samples for PCR. Nucleic Acids Res. 1993, 21, 4153–4154. [Google Scholar] [CrossRef] [Green Version]
- Kondo, T. The 3-terminal sequence of Chinese yam necrotic mosaic virus genomic RNA: A close relationship with macluravirus. Arch. Virol. 2001, 146, 1527–1535. [Google Scholar] [CrossRef]
- Zhang, P.; Peng, J.; Guo, H.; Chen, J.; Chen, S.; Wang, J. Complete genome sequence of yam chlorotic necrotic mosaic virus from Dioscorea parviflora. Arch. Virol. 2016, 161, 1715–1717. [Google Scholar] [CrossRef]
- Filloux, D.; Fernandez, E.; Loire, E.; Claude, L.; Galzi, S.; Candresse, T.; Winter, S.; Jeeva, M.L.; Makeshkumar, T.; Martin, D.P.; et al. Nanopore-based detection and characterization of yam viruses. Sci. Rep. 2018, 8, 17879. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Son, C.; Kwon, J.; Nam, H.; Kim, Y.; Lee, S.; Zhao, F.; Moon, S. Complete genome sequence of Chinese Yam necrotic mosaic virus from Dioscorea opposita in the Republic of Korea. Genome Annoucements 2016, 4, e00778-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filloux, D.; Bonheur, L.; Umber, M.; Pavis, C.; Fernandez, E.; Galzi, S.; Julian, C.; Daugrois, J.H.; Sukal, A.; Winter, S.; et al. Metagenomic discovery, worldwide distribution and genetic diversity of novel macluraviruses infecting yams (Dioscorea spp.). In Proceedings of the 15èmes Rencontres de Virologie Végétale, Aussois, France, 18–22 January 2015; p. 86. [Google Scholar]
- Teycheney, P.Y.; Geering, A.D.W.; Dasgupta, I.; Hull, R.; Kreuze, J.F.; Lockhart, B.; Muller, E.; Olszewski, N.; Pappu, H.; Pooggin, M.M.; et al. ICTV Virus taxonomy profile: Caulimoviridae. J. Gen. Virol. 2020, 101, 1025–1026. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.; Hohn, T.; Selvarajan, R. Badnaviruses: The current global scenario. Viruses 2016, 8, 177. [Google Scholar] [CrossRef] [Green Version]
- Sukal, A.C.; Kidanemariam, D.B.; Dale, J.L.; Harding, R.M.; James, A.P. Characterization and genetic diversity of Dioscorea bacilliform viruses present in a Pacific yam germplasm collection. Plant Pathol. 2020, 69, 576–584. [Google Scholar] [CrossRef]
- Galzi, S.; Scutt, R.; Prophète, P.; Roumagnac, P.; Filloux, D. Assessment and characterization of the genetic diversity of viruses infecting cultivated yams (Dioscorea spp.) in Haïti. In Proceedings of the 14èmes Rencontres de Virologie Végétale, Aussois, France, 13–17 January 2013; p. 70. [Google Scholar]
- Bömer, M.; Turaki, A.; Silva, G.; Kumar, P.; Seal, S. A sequence-independent strategy for amplification and characterization of episomal badnavirus sequences reveals three previously uncharacterized yam badnaviruses. Viruses 2016, 8, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umber, M.; Gomez, R.; Gelabale, S.; Bonheur, L.; Pavis, C.; Teycheney, P. The genome sequence of Dioscorea bacilliform TR virus, a member of the genus Badnavirus infecting Dioscorea spp., sheds light on the possible function of endogenous Dioscorea bacilliform viruses. Arch. Virol. 2016, 162, 517–521. [Google Scholar] [CrossRef]
- Mantell, S.H.; Haque, S.Q. Internal brown spot disease of yams. Yam Virus Proj. Bull. (Trinidad Tobago) 1979, ill, 1–13. [Google Scholar]
- Harrison, B.; Roberts, I. Association of virus-like particles with internal brown spot of yam (Dioscorea alata). Trop. Agric. 1979, 50, 335–340. [Google Scholar]
- Eni, A.O.; Hughes, J.D.A.; Asiedu, R.; Rey, M.E.C. Sequence diversity among badnavirus isolates infecting yam (Dioscorea spp.) in Ghana, Togo, Benin and Nigeria. Arch. Virol. 2008, 153, 2263–2272. [Google Scholar] [CrossRef]
- Briddon, R.W.; Phillips, S.; Brunt, A.; Hull, R. Analysis of the sequence of Dioscorea alata bacilliform virus; comparison to other members of the badnavirus group. Virus Genes 1999, 18, 277–283. [Google Scholar] [CrossRef]
- Seal, S.; Muller, E. Molecular analysis of a full-length sequence of a new yam badnavirus from Dioscorea sansibarensis. Arch. Virol. 2007, 152, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Sukal, A.; Kidanemariam, D.; Dale, J.; James, A.; Harding, R. Characterization of badnaviruses infecting Dioscorea spp. in the Pacific reveals two putative novel species and the first report of Dioscorea bacilliform RT virus 2. Virus Res. 2017, 238, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bousalem, M.; Durand, O.; Scarcelli, N.; Lebas, B.S.M.; Kenyon, L.; Marchand, J.L.; Lefort, F.; Seal, S.E. Dilemmas caused by endogenous pararetroviruses regarding the taxonomy and diagnosis of yam (Dioscorea spp.) badnaviruses: Analyses to support safe germplasm movement. Arch. Virol. 2009, 154, 297–314. [Google Scholar] [CrossRef] [PubMed]
- Turaki, A.A.; Bömer, M.; Silva, G.; Lava Kumar, P.; Seal, S.E. PCR-DGGE analysis: Unravelling complex mixtures of badnavirus sequences present in yam germplasm. Viruses 2017, 9, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umber, M.; Filloux, D.; Muller, E.; Laboureau, N.; Galzi, S.; Roumagnac, P.; Pavis, C.; Teycheney, P.; Seal, S.E. The genome of African yam (Dioscorea cayenensis-rotundata complex) hosts endogenous sequences from four distinct badnavirus species. Mol. Plant Pathol. 2014, 15, 790–801. [Google Scholar] [CrossRef]
- Homs, M.; Kober, S.; Kepp, G.; Jeske, H. Mitochondrial plasmids of sugar beet amplified via rolling circle method detected during curtovirus screening. Virus Res. 2008, 136, 124–129. [Google Scholar] [CrossRef]
- Paprotka, T.; Boiteux, L.S.; Fonseca, M.E.N.; Resende, R.O.; Jeske, H.; Faria, J.C.; Ribeiro, S.G. Genomic diversity of sweet potato geminiviruses in a Brazilian germplasm bank. Virus Res. 2010, 149, 224–233. [Google Scholar] [CrossRef]
- James, A.P.; Geijskes, R.J.; Dale, J.L.; Harding, R.M. Development of a novel rolling-circle amplification technique to detect banana streak virus that also discriminates between integrated and episomal virus sequences. Plant Dis. 2011, 95, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Sukal, A.C.; Kidanemariam, D.B.; Dale, J.L.; Harding, R.M.; James, A.P. Assessment and optimization of rolling circle amplification protocols for the detection and characterization of badnaviruses. Virology 2019, 529, 73–80. [Google Scholar] [CrossRef]
- Cordoñer, F.M.; Elena, S.F. The promiscuous evolutionary history of the family Bromoviridae. J. Gen. Virol. 2008, 89, 1739–1747. [Google Scholar] [CrossRef]
- Bujarski, J.; Gallitelli, D.; García-Arenal, F.; Pallás, V.; Palukaitis, P.; Krishna Reddy, M.; Wang, A. ICTV virus taxonomy profile: Bromoviridae. J. Gen. Virol. 2019, 100, 1206–1207. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yu, C.; Yin, Y.; Gao, S.; Wang, F.; Jiao, C.; Yao, M. Pepper crop improvement against cucumber mosaic virus (CMV): A review. Front. Plant Sci. 2020, 11, 1932. [Google Scholar] [CrossRef] [PubMed]
- Jacquemond, M. Cucumber Mosaic Virus. Adv. Virus Res. 2012, 84, 439–504. [Google Scholar] [CrossRef] [PubMed]
- Migliori, A.M. A Virus De L’Igname (Dioscorea sp.). In Proceedings of the 14th Annual Meeting of the Caribbean Food Crops Society, Guadeloupe and Martinique, France, 27 June–2 July 1977; pp. 428–435. [Google Scholar]
- Eni, A.O.; Hughes, J.A.; Asiedu, R.; Rey, M.E.C. Survey of the incidence and distribution of viruses infecting yam (Dioscorea spp.) in Ghana and Togo. Ann. Appl. Biol. 2010, 156, 243–252. [Google Scholar] [CrossRef]
- Eni, A.O.; Hughes, J.A.; Rey, M.E.C. Production of polyclonal antibodies against a yam isolate of cucumber mosaic virus (CMV). Res. J. Agric. Biol. Sci. 2010, 6, 607–612. [Google Scholar]
- Hayashi, E.A.I.; Blawid, R.; Lucas de Melo, F.; Andrade, M.S.; Pio-Ribeiro, G.; de Andrade, G.P.; Nagata, T. Complete genome sequence of a putative new secovirus infecting yam (Dioscorea spp.) plants. Arch. Virol. 2016, 162, 317–319. [Google Scholar] [CrossRef]
- Dey, K.K.; Sugikawa, J.; Kerr, C.; Melzer, M.J. Air potato (Dioscorea bulbifera) plants displaying virus-like symptoms are co-infected with a novel potyvirus and a novel ampelovirus. Virus Genes 2018, 55, 117–121. [Google Scholar] [CrossRef]
- Wang, M.R.; Cui, Z.H.; Li, J.W.; Hao, X.Y.; Zhao, L.; Wang, Q.C. In vitro thermotherapy-based methods for plant virus eradication. Plant Methods 2018, 14, 87. [Google Scholar] [CrossRef] [Green Version]
- Rubio, L.; Galipienso, L.; Ferriol, I. Detection of plant viruses and disease management: Relevance of genetic diversity and evolution. Front. Plant Sci. 2020, 11, 1092. [Google Scholar] [CrossRef]
- Panattoni, A.; Luvisi, A.; Triolo, E. Review. Elimination of viruses in plants: Twenty years of progress. Spanish J. Agric. Res. 2013, 11, 173–188. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.; Igiraneza, C.; Dusengemungu, L. Major in vitro techniques for potato virus elimination and post eradication detection methods. A review. Am. J. Potato Res. 2019, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.H.; Kang, D.K.; Sohn, J.K. Production of yam mosaic virus (YMV)-free Dioscorea opposita plants by cryotherapy of shoot-tips. Cryo-Letters 2013, 34, 149–157. [Google Scholar] [PubMed]
- Jayaseelan, D.; Rajitha, M.; Dhanya, M.K.; Hegde, V.; Jeeva, M.L. Hot water treatment: An efficient method for elimination of yam mild mosaic virus in Dioscorea alata. J. Root Crops 2011, 37, 60–64. [Google Scholar]
- González Ramírez, J.E.; Cabrera Jova, M.; Robaina, A.; Rodríguez Pérez, D.; González Cadalso, A.; Portal, O. Water-dissolved ozone mediates potyvirus sanitation during in vitro propagation of Dioscorea cayenensis subsp. rotundata (Poir.) Miège. Ozone Sci. Eng. 2020, 42, 89–94. [Google Scholar] [CrossRef]
- Lebas, B.M. Diversity of Viruses Infecting Dioscorea Species in the South Pacific. Ph.D. Thesis, The University of Greenwich, London, UK, 2002. [Google Scholar]
- Siadjeu, C.; Pucker, B.; Viehöver, P.; Albach, D.C.; Weisshaar, B. High contiguity de novo genome sequence assembly of trifoliate yam (Dioscorea dumetorum) using long-read sequencing. Genes 2020, 11, 274. [Google Scholar] [CrossRef] [Green Version]
- Saski, C.A.; Bhattacharjee, R.; Scheffler, B.E.; Asiedu, R. Genomic Resources for Water Yam (Dioscorea alata L.): Analyses of EST-sequences, De Novo sequencing and GBS libraries. PLoS ONE 2015, 10, e0134031. [Google Scholar] [CrossRef] [Green Version]
- Cormier, F.; Lawac, F.; Maledon, E.; Gravillon, M.-C.; Nudol, E.; Mournet, P.; Vignes, H.; Chaïr, H.; Arnau, G. A reference high-density genetic map of greater yam (Dioscorea alata L.). Theor. Appl. Genet. 2019, 132, 1733–1744. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, G.-F.; Podolyan, A.; Kidanemariam, D.B.; Pilotti, C.; Houliston, G.; Sukal, A.C. A Review of Viruses Infecting Yam (Dioscorea spp.). Viruses 2022, 14, 662. https://doi.org/10.3390/v14040662
Luo G-F, Podolyan A, Kidanemariam DB, Pilotti C, Houliston G, Sukal AC. A Review of Viruses Infecting Yam (Dioscorea spp.). Viruses. 2022; 14(4):662. https://doi.org/10.3390/v14040662
Chicago/Turabian StyleLuo, Guo-Fu, Ana Podolyan, Dawit B. Kidanemariam, Carmel Pilotti, Gary Houliston, and Amit C. Sukal. 2022. "A Review of Viruses Infecting Yam (Dioscorea spp.)" Viruses 14, no. 4: 662. https://doi.org/10.3390/v14040662
APA StyleLuo, G. -F., Podolyan, A., Kidanemariam, D. B., Pilotti, C., Houliston, G., & Sukal, A. C. (2022). A Review of Viruses Infecting Yam (Dioscorea spp.). Viruses, 14(4), 662. https://doi.org/10.3390/v14040662