
Brain Inflammation and Intracellular α-Synuclein Aggregates in Macaques after SARS-CoV-2 Infection

 , , ,
, , ,  , , , , ,
, , , , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and SARS-CoV-2 Infection
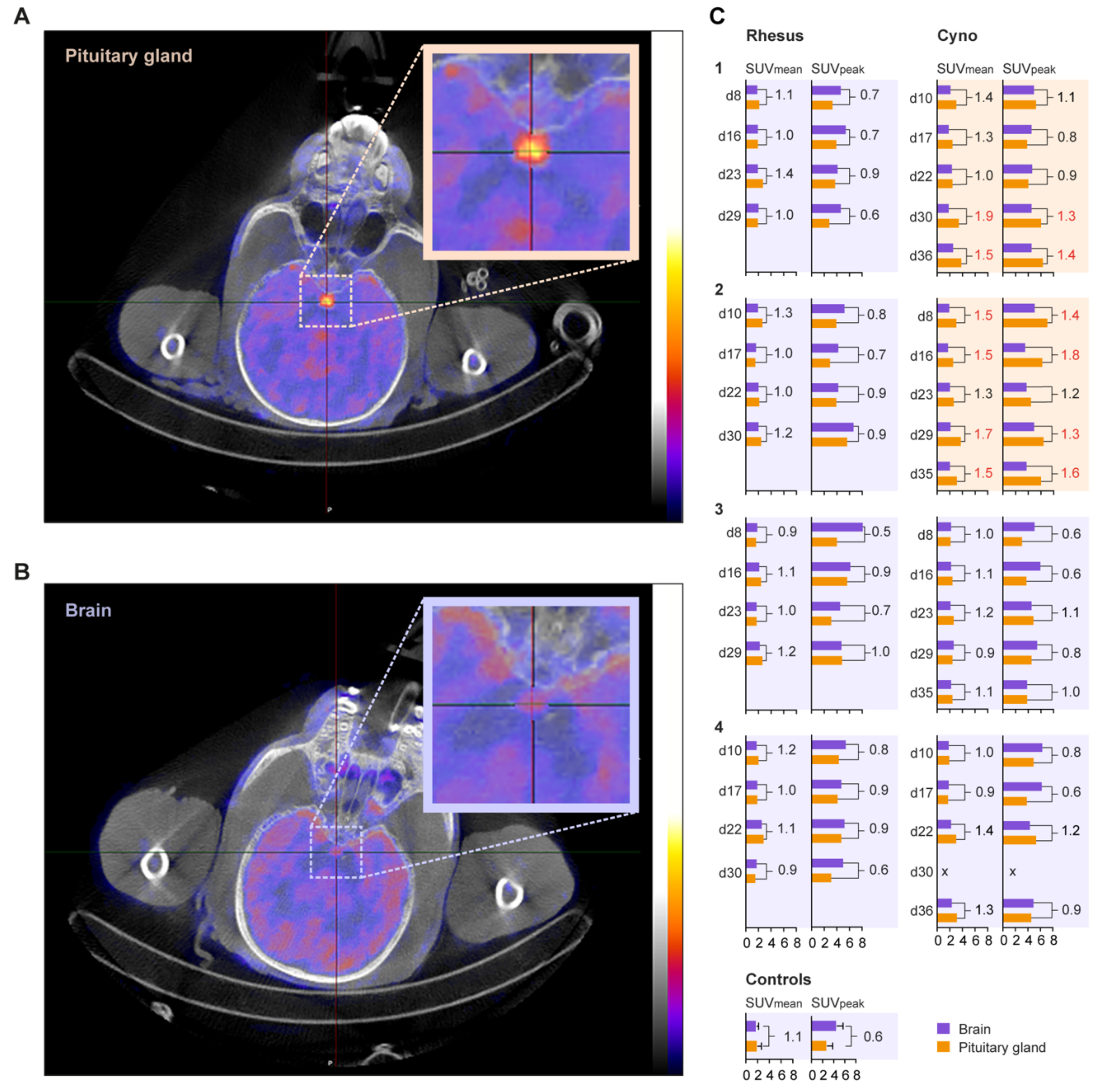
2.2. Positron Emission Tomography-Computed Tomography of the Brain
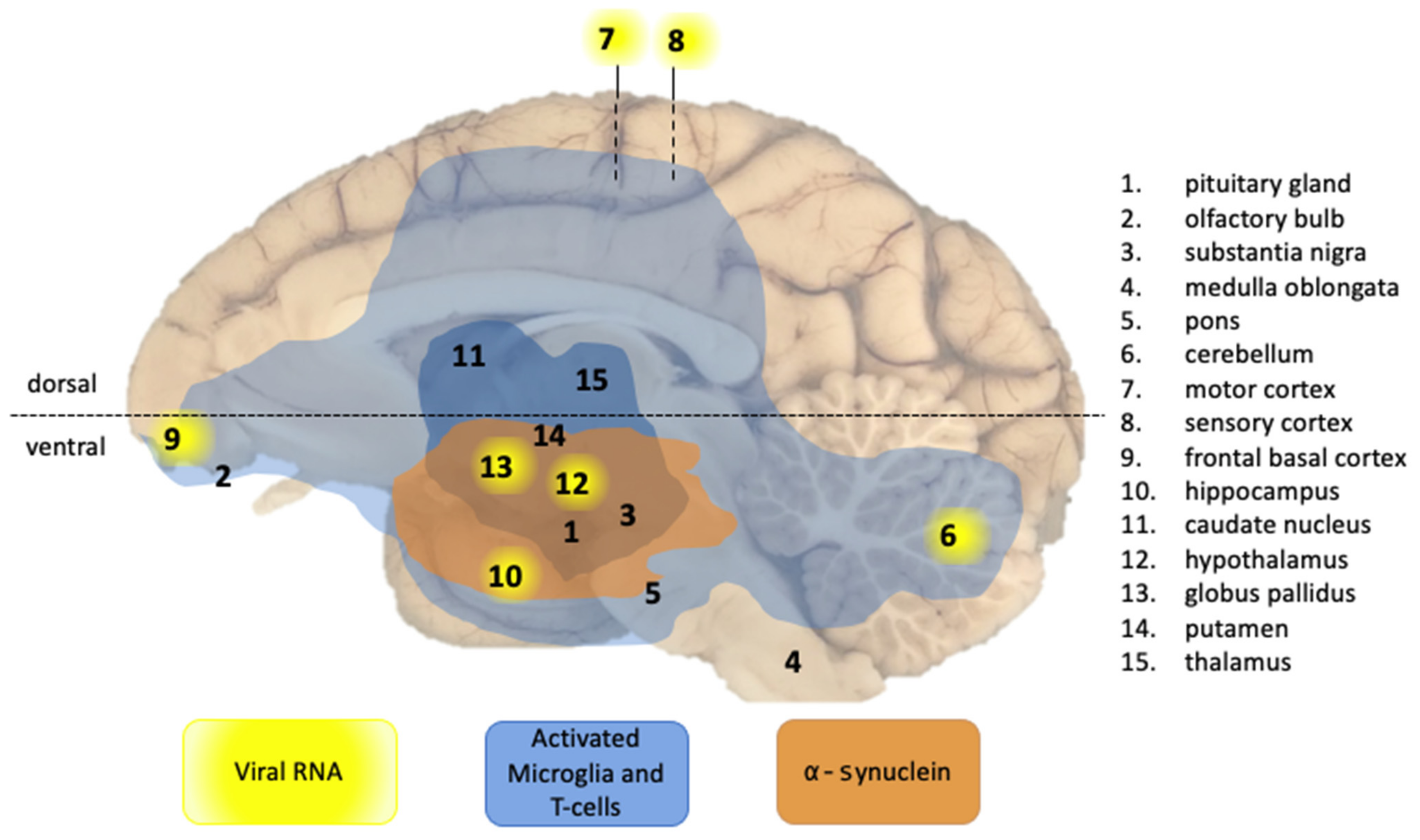
2.3. Brain Tissue Sampling
2.4. Viral RNA Detection in Brain Tissue
2.5. Immunohistochemistry
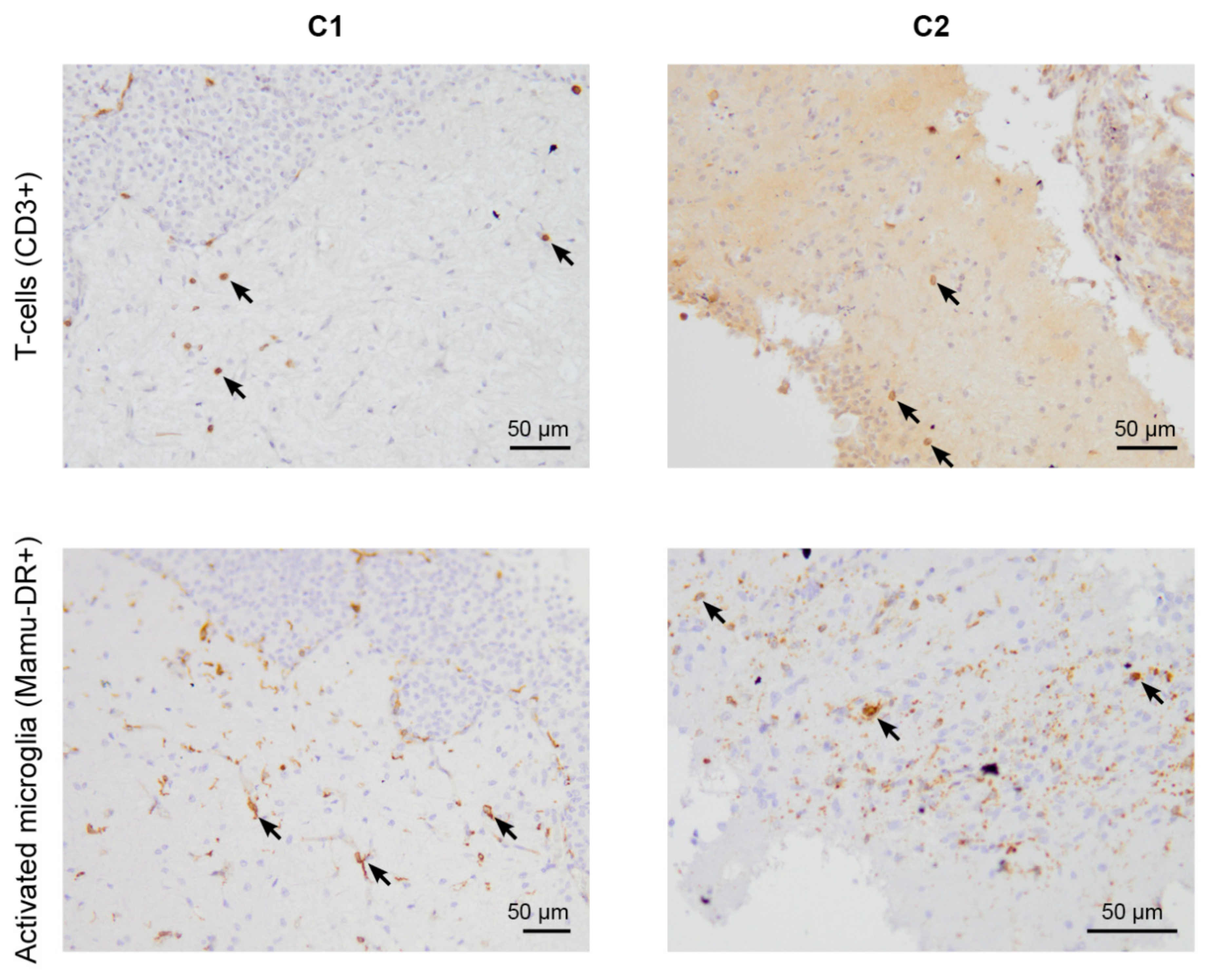
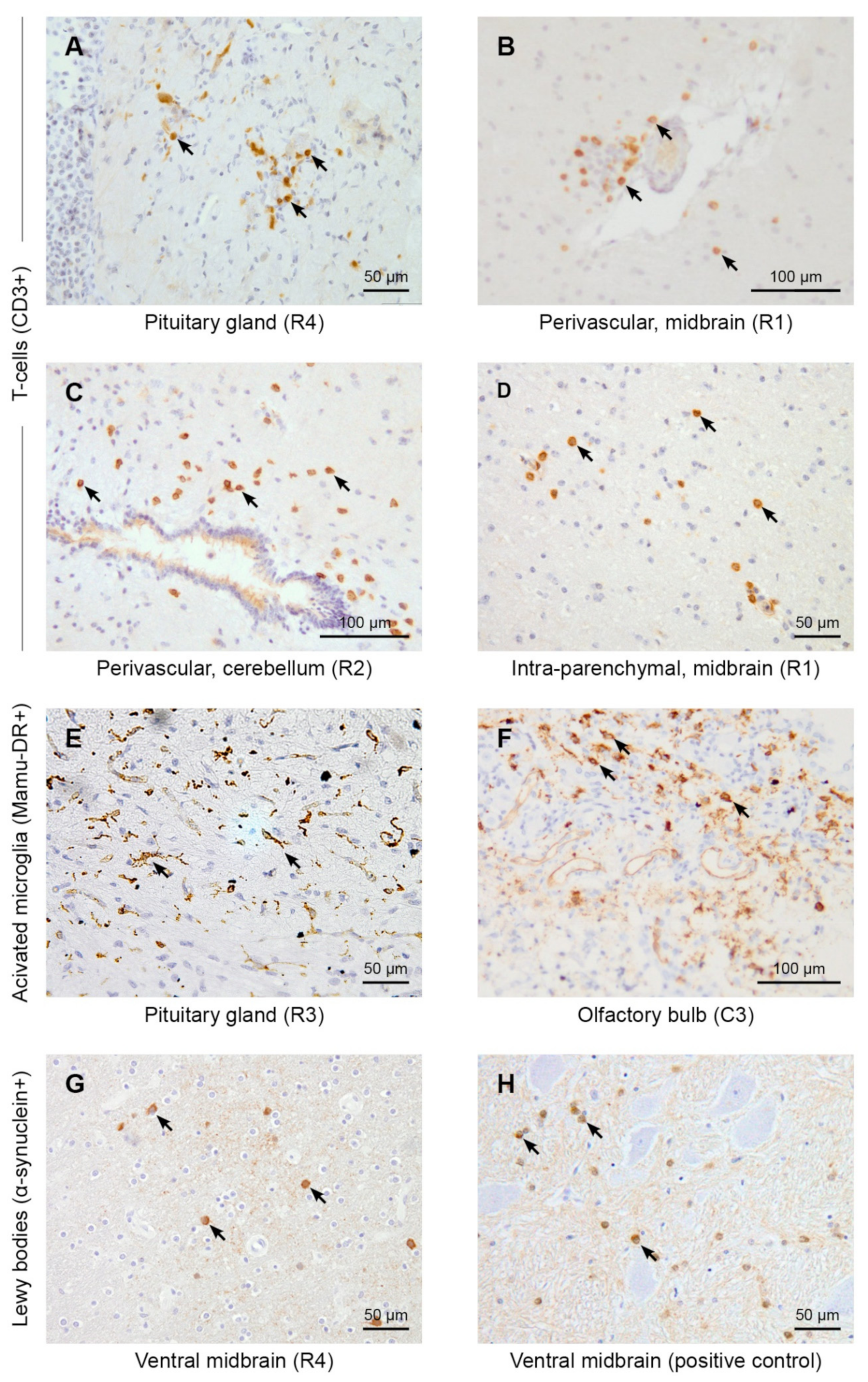
2.5.1. CD3 and CD20
2.5.2. Mamu-DR Staining
2.5.3. α-Synuclein
2.5.4. SARS-CoV-2
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fotuhi, M.; Mian, A.; Meysami, S.; Raji, C.A. Neurobiology of COVID-19. J. Alzheimers Dis. 2020, 76, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Romoli, M.; Jelcic, I.; Bernard-Valnet, R.; Garcia Azorin, D.; Mancinelli, L.; Akhvlediani, T.; Monaco, S.; Taba, P.; Sellner, J.; Infectious Disease Panel of the European Academy of Neurology. A systematic review of neurological manifestations of SARS-CoV-2 infection: The devil is hidden in the details. Eur. J. Neurol. 2020, 27, 1712–1726. [Google Scholar] [CrossRef] [PubMed]
- Merello, M.; Bhatia, K.P.; Obeso, J.A. SARS-CoV-2 and the risk of Parkinson’s disease: Facts and fantasy. Lancet Neurol. 2021, 20, 94–95. [Google Scholar] [CrossRef]
- Cohen, M.E.; Eichel, R.; Steiner-Birmanns, B.; Janah, A.; Ioshpa, M.; Bar-Shalom, R.; Paul, J.J.; Gaber, H.; Skrahina, V.; Bornstein, N.M.; et al. A case of probable Parkinson’s disease after SARS-CoV-2 infection. Lancet Neurol. 2020, 19, 804–805. [Google Scholar] [CrossRef]
- Faber, I.; Brandao, P.R.P.; Menegatti, F.; de Carvalho Bispo, D.D.; Maluf, F.B.; Cardoso, F. Coronavirus Disease 2019 and Parkinsonism: A Non-post-encephalitic Case. Mov. Disord. 2020, 35, 1721–1722. [Google Scholar] [CrossRef]
- Mendez-Guerrero, A.; Laespada-Garcia, M.I.; Gomez-Grande, A.; Ruiz-Ortiz, M.; Blanco-Palmero, V.A.; Azcarate-Diaz, F.J.; Rabano-Suarez, P.; Alvarez-Torres, E.; de Fuenmayor-Fernandez de la Hoz, C.P.; Vega Perez, D.; et al. Acute hypokinetic-rigid syndrome following SARS-CoV-2 infection. Neurology 2020, 95, e2109–e2118. [Google Scholar] [CrossRef]
- Schurink, B.; Roos, E.; Radonic, T.; Barbe, E.; Bouman, C.S.C.; de Boer, H.H.; de Bree, G.J.; Bulle, E.B.; Aronica, E.M.; Florquin, S.; et al. Viral presence and immunopathology in patients with lethal COVID-19: A prospective autopsy cohort study. Lancet Microbe 2020, 1, e290–e299. [Google Scholar] [CrossRef]
- Yang, A.C.; Kern, F.; Losada, P.M.; Agam, M.R.; Maat, C.A.; Schmartz, G.P.; Fehlmann, T.; Stein, J.A.; Schaum, N.; Lee, D.P.; et al. Dysregulation of brain and choroid plexus cell types in severe COVID-19. Nature 2021, 595, 565–571. [Google Scholar] [CrossRef]
- Zeiss, C.J.; Compton, S.; Veenhuis, R.T. Animal Models of COVID-19. I. Comparative Virology and Disease Pathogenesis. ILAR J. 2021. [Google Scholar] [CrossRef]
- Wan, D.; Du, T.; Hong, W.; Chen, L.; Que, H.; Lu, S.; Peng, X. Neurological complications and infection mechanism of SARS-CoV-2. Signal Transduct. Target Ther. 2021, 6, 406. [Google Scholar] [CrossRef]
- Vidal, E.; Lopez-Figueroa, C.; Rodon, J.; Perez, M.; Brustolin, M.; Cantero, G.; Guallar, V.; Izquierdo-Useros, N.; Carrillo, J.; Blanco, J.; et al. Chronological brain lesions after SARS-CoV-2 infection in hACE2-transgenic mice. Vet. Pathol. 2021, 03009858211066841. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, L.; Bao, L.; Liu, J.; Zhu, H.; Lv, Q.; Liu, R.; Chen, W.; Tong, W.; Wei, Q.; et al. SARS-CoV-2 crosses the blood-brain barrier accompanied with basement membrane disruption without tight junctions alteration. Signal Transduct. Target Ther. 2021, 6, 337. [Google Scholar] [CrossRef] [PubMed]
- Golden, J.W.; Li, R.; Cline, C.R.; Zeng, X.; Mucker, E.M.; Fuentes-Lao, A.J.; Spik, K.W.; Williams, J.A.; Twenhafel, N.; Davis, N.; et al. Hamsters Expressing Human Angiotensin-Converting Enzyme 2 Develop Severe Disease following Exposure to SARS-CoV-2. mBio 2022, 13, e02906-21. [Google Scholar] [CrossRef] [PubMed]
- Fagre, A.; Lewis, J.; Eckley, M.; Zhan, S.; Rocha, S.M.; Sexton, N.R.; Burke, B.; Geiss, B.; Peersen, O.; Bass, T.; et al. SARS-CoV-2 infection, neuropathogenesis and transmission among deer mice: Implications for spillback to New World rodents. PLoS Pathog. 2021, 17, e1009585. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, L.; Bishop, E.; Jay, B.; Lafoux, B.; Minoves, M.; Passaes, C. COVID-19 Research: Lessons from Non-Human Primate Models. Vaccines 2021, 9, 886. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; Kanevsky, I.; Yildiz, S.; Sellers, R.S.; Swanson, K.A.; Franks, T.; Rathnasinghe, R.; Munoz-Moreno, R.; Jangra, S.; Gonzalez, O.; et al. Modeling SARS-CoV-2: Comparative Pathology in Rhesus Macaque and Golden Syrian Hamster Models. Toxicol. Pathol. 2022, 01926233211072767. [Google Scholar] [CrossRef]
- Jiao, L.; Yang, Y.; Yu, W.; Zhao, Y.; Long, H.; Gao, J.; Ding, K.; Ma, C.; Li, J.; Zhao, S.; et al. The olfactory route is a potential way for SARS-CoV-2 to invade the central nervous system of rhesus monkeys. Signal Transduct. Target Ther. 2021, 6, 169. [Google Scholar] [CrossRef]
- Böszörményi, K.P.; Stammes, M.A.; Fagrouch, Z.C.; Kiemenyi-Kayere, G.; Niphuis, H.; Mortier, D.; van Driel, N.; Nieuwenhuis, I.; Vervenne, R.A.W.; Haaksma, T.; et al. The Post-Acute Phase of SARS-CoV-2 Infection in Two Macaque Species Is Associated with Signs of Ongoing Virus Replication and Pathology in Pulmonary and Extrapulmonary Tissues. Viruses 2021, 13, 1673. [Google Scholar] [CrossRef]
- Tulisiak, C.T.; Mercado, G.; Peelaerts, W.; Brundin, L.; Brundin, P. Can infections trigger alpha-synucleinopathies? Prog. Mol. Biol. Transl. Sci. 2019, 168, 299–322. [Google Scholar] [CrossRef]
- Stammes, M.A.; Bakker, J.; Vervenne, R.A.W.; Zijlmans, D.G.M.; van Geest, L.; Vierboom, M.P.M.; Langermans, J.A.M.; Verreck, F.A.W. Recommendations for Standardizing Thorax PET–CT in Non-Human Primates by Recent Experience from Macaque Studies. Animals 2021, 11, 204. [Google Scholar] [CrossRef]
- Paxinos, G.; Huang, X.F.; Petrides, M.; Toga, A. The Rhesus Monkey Brain in Stereotaxic Coordinates, 2nd ed.; Elsevier Science Publishing Co. Inc.: San Diego, CA, USA, 2008; p. 416. [Google Scholar]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brunink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar] [CrossRef] [Green Version]
- Wolfel, R.; Corman, V.M.; Guggemos, W.; Seilmaier, M.; Zange, S.; Muller, M.A.; Niemeyer, D.; Jones, T.C.; Vollmar, P.; Rothe, C.; et al. Virological assessment of hospitalized patients with COVID-2019. Nature 2020, 581, 465–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pijl, J.P.; Nienhuis, P.H.; Kwee, T.C.; Glaudemans, A.; Slart, R.; Gormsen, L.C. Limitations and Pitfalls of FDG-PET/CT in Infection and Inflammation. Semin. Nucl. Med. 2021, 51, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.A.; Kim, J.I.; Lee, J.W.; Cho, Y.J.; Lee, B.H.; Chung, H.W.; Park, K.K.; Han, J.S. Effects of various anesthetic protocols on 18F-flurodeoxyglucose uptake into the brains and hearts of normal miniature pigs (Sus scrofa domestica). J. Am. Assoc. Lab. Anim. Sci. 2012, 51, 246–252. [Google Scholar] [PubMed]
- Meyer, M.; Allenbach, G.; Nicod Lalonde, M.; Schaefer, N.; Prior, J.O.; Gnesin, S. Increased (18)F-FDG signal recovery from small physiological structures in digital PET/CT and application to the pituitary gland. Sci. Rep. 2020, 10, 368. [Google Scholar] [CrossRef]
- Goutal, S.; Tournier, N.; Guillermier, M.; Van Camp, N.; Barret, O.; Gaudin, M.; Bottlaender, M.; Hantraye, P.; Lavisse, S. Comparative test-retest variability of outcome parameters derived from brain [18F]FDG PET studies in non-human primates. PLoS ONE 2020, 15, e0240228. [Google Scholar] [CrossRef]
- Boellaard, R. Standards for PET image acquisition and quantitative data analysis. J. Nucl. Med. 2009, 50 (Suppl. 1), 11S–20S. [Google Scholar] [CrossRef] [Green Version]
- Ju, H.; Zhou, J.; Pan, Y.; Lv, J.; Zhang, Y. Evaluation of pituitary uptake incidentally identified on (18)F-FDG PET/CT scan. Oncotarget 2017, 8, 55544–55549. [Google Scholar] [CrossRef] [Green Version]
- Hyun, S.H.; Choi, J.Y.; Lee, K.H.; Choe, Y.S.; Kim, B.T. Incidental focal 18F-FDG uptake in the pituitary gland: Clinical significance and differential diagnostic criteria. J. Nucl. Med. 2011, 52, 547–550. [Google Scholar] [CrossRef] [Green Version]
- Chua, M.W.J.; Chua, M.P.W. Delayed Onset of Central Hypocortisolism in a Patient Recovering From COVID-19. AACE Clin. Case Rep. 2021, 7, 2–5. [Google Scholar] [CrossRef]
- Tan, T.; Khoo, B.; Mills, E.G.; Phylactou, M.; Patel, B.; Eng, P.C.; Thurston, L.; Muzi, B.; Meeran, K.; Prevost, A.T.; et al. Association between high serum total cortisol concentrations and mortality from COVID-19. Lancet Diabetes Endocrinol. 2020, 8, 659–660. [Google Scholar] [CrossRef]
- Alshebri, M.S.; Alshouimi, R.A.; Alhumidi, H.A.; Alshaya, A.I. Neurological Complications of SARS-CoV, MERS-CoV, and COVID-19. SN Compr. Clin. Med. 2020, 2, 2037–2047. [Google Scholar] [CrossRef] [PubMed]
- Zubair, A.S.; McAlpine, L.S.; Gardin, T.; Farhadian, S.; Kuruvilla, D.E.; Spudich, S. Neuropathogenesis and Neurologic Manifestations of the Coronaviruses in the Age of Coronavirus Disease 2019: A Review. JAMA Neurol. 2020, 77, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Sulzer, D.; Antonini, A.; Leta, V.; Nordvig, A.; Smeyne, R.J.; Goldman, J.E.; Al-Dalahmah, O.; Zecca, L.; Sette, A.; Bubacco, L.; et al. COVID-19 and possible links with Parkinson’s disease and parkinsonism: From bench to bedside. NPJ Parkinsons Dis. 2020, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Philippens, I.H.; Ormel, P.R.; Baarends, G.; Johansson, M.; Remarque, E.J.; Doverskog, M. Acceleration of Amyloidosis by Inflammation in the Amyloid-Beta Marmoset Monkey Model of Alzheimer’s Disease. J. Alzheimers Dis. 2017, 55, 101–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douaud, G.; Lee, S.; Alfaro-Almagro, F.; Arthofer, C.; Wang, C.; Lange, F.; Andersson, J.L.R.; Griffanti, L.; Duff, E.; Jbabdi, S.; et al. Brain imaging before and after COVID-19 in UK Biobank. medRxiv 2021. [Google Scholar] [CrossRef]
- Song, E.; Zhang, C.; Israelow, B.; Lu-Culligan, A.; Prado, A.V.; Skriabine, S.; Lu, P.; Weizman, O.E.; Liu, F.; Dai, Y.; et al. Neuroinvasion of SARS-CoV-2 in human and mouse brain. J. Exp. Med. 2021, 218, e20202135. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Age (years) | Weight (kg) | Cum VL 1 | Cum CT 2 | Shedding (dpi) 3 | Seroconversion (dpi) 4 | Euthanasia (dpi) | |
|---|---|---|---|---|---|---|---|---|
| Rhesus | ||||||||
| R14002 | R1 | 6 | 8.2 | 2.10 × 107 | 12 | 1–10 | 16 | 36 |
| R15080 | R2 | 5 | 7.9 | 2.69 × 106 | 8 | 1–4 | 16 | 35 |
| R15090 | R3 | 5 | 7.8 | 4.64 × 105 | 4 | 1–5 | 16 | 36 |
| R15096 | R4 | 5 | 8.7 | 1.70 × 104 | 22 | 1 | 16 | 35 |
| Cynomolgus | ||||||||
| J16004 | C1 | 4 | 5.7 | 1.28 × 105 | 14 | 1–4 | 12 | 42 |
| J16012 | C2 | 4 | 3.3 | 3.00 × 105 | 30 | 1–2 | 12 | 38 |
| J16017 | C3 | 4 | 4.9 | 1.92 × 108 | 9 | 1–6 | 12 | 38 |
| Ji04080 | C4 | 16 | 9.7 | 1.19 × 105 | 18 | 1–3 | 17 | 42 |
| Marker | Brain Area | Rhesus | Cynomolgus | Controls | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R1 | R2 | R3 | R4 | C1 | C2 | C3 | C4 | R5 | R6 | C5 | C6 | |||||||
| T-cells (CD3+) | intra-parenchymal | pituitary gland | ||||||||||||||||
| olfactory bulb | ||||||||||||||||||
| front brain | ||||||||||||||||||
| dorsal | ||||||||||||||||||
| ventral | ⊙ | |||||||||||||||||
| cerebellum | ||||||||||||||||||
| perivasculair | pituitary gland | |||||||||||||||||
| olfactory bulb | ||||||||||||||||||
| front brain | ||||||||||||||||||
| dorsal | ||||||||||||||||||
| ventral | ||||||||||||||||||
| cerebellum | ||||||||||||||||||
| nodules | pituitary gland | |||||||||||||||||
| olfactory bulb | ||||||||||||||||||
| front brain | ||||||||||||||||||
| dorsal | ||||||||||||||||||
| ventral | ||||||||||||||||||
| cerebellum | ||||||||||||||||||
| meninges | pituitary gland | |||||||||||||||||
| olfactory bulb | ||||||||||||||||||
| front brain | ||||||||||||||||||
| dorsal | ||||||||||||||||||
| ventral | ||||||||||||||||||
| cerebellum | ||||||||||||||||||
| Activated microglia (Mamu-DR+) | presence | pituitary gland | ||||||||||||||||
| olfactory bulb | ||||||||||||||||||
| front brain | ||||||||||||||||||
| dorsal | ||||||||||||||||||
| ventral | ||||||||||||||||||
| cerebellum | ||||||||||||||||||
| morphology: ramified/ amoeboid | pituitary gland | |||||||||||||||||
| olfactory bulb | ||||||||||||||||||
| front brain | ||||||||||||||||||
| dorsal | ||||||||||||||||||
| ventral | ||||||||||||||||||
| cerebellum | ||||||||||||||||||
| nodules | pituitary gland | |||||||||||||||||
| olfactory bulb | ||||||||||||||||||
| front brain | ||||||||||||||||||
| dorsal | ||||||||||||||||||
| ventral | Observation of indicated marker: | |||||||||||||||||
| cerebellum | ||||||||||||||||||
| meninges | pituitary gland | light (inlc. ramified microglia) | ||||||||||||||||
| olfactory bulb | ||||||||||||||||||
| front brain | moderate (incl. amoeboid microglia) | |||||||||||||||||
| dorsal | ||||||||||||||||||
| ventral | moderate to severe | |||||||||||||||||
| cerebellum | ||||||||||||||||||
| α-synuclein+ | inclusions | ventral midbrain | ⊙ | infarction | ||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Philippens, I.H.C.H.M.; Böszörményi, K.P.; Wubben, J.A.M.; Fagrouch, Z.C.; van Driel, N.; Mayenburg, A.Q.; Lozovagia, D.; Roos, E.; Schurink, B.; Bugiani, M.; et al. Brain Inflammation and Intracellular α-Synuclein Aggregates in Macaques after SARS-CoV-2 Infection. Viruses 2022, 14, 776. https://doi.org/10.3390/v14040776
Philippens IHCHM, Böszörményi KP, Wubben JAM, Fagrouch ZC, van Driel N, Mayenburg AQ, Lozovagia D, Roos E, Schurink B, Bugiani M, et al. Brain Inflammation and Intracellular α-Synuclein Aggregates in Macaques after SARS-CoV-2 Infection. Viruses. 2022; 14(4):776. https://doi.org/10.3390/v14040776
Chicago/Turabian StylePhilippens, Ingrid H. C. H. M., Kinga P. Böszörményi, Jacqueline A. M. Wubben, Zahra C. Fagrouch, Nikki van Driel, Amber Q. Mayenburg, Diana Lozovagia, Eva Roos, Bernadette Schurink, Marianna Bugiani, and et al. 2022. "Brain Inflammation and Intracellular α-Synuclein Aggregates in Macaques after SARS-CoV-2 Infection" Viruses 14, no. 4: 776. https://doi.org/10.3390/v14040776
APA StylePhilippens, I. H. C. H. M., Böszörményi, K. P., Wubben, J. A. M., Fagrouch, Z. C., van Driel, N., Mayenburg, A. Q., Lozovagia, D., Roos, E., Schurink, B., Bugiani, M., Bontrop, R. E., Middeldorp, J., Bogers, W. M., de Geus-Oei, L. -F., Langermans, J. A. M., Verschoor, E. J., Stammes, M. A., & Verstrepen, B. E. (2022). Brain Inflammation and Intracellular α-Synuclein Aggregates in Macaques after SARS-CoV-2 Infection. Viruses, 14(4), 776. https://doi.org/10.3390/v14040776