
A Single Amino Acid Residue R144 of SNX16 Affects Its Ability to Inhibit the Replication of Influenza A Virus

 ,
,  and
and 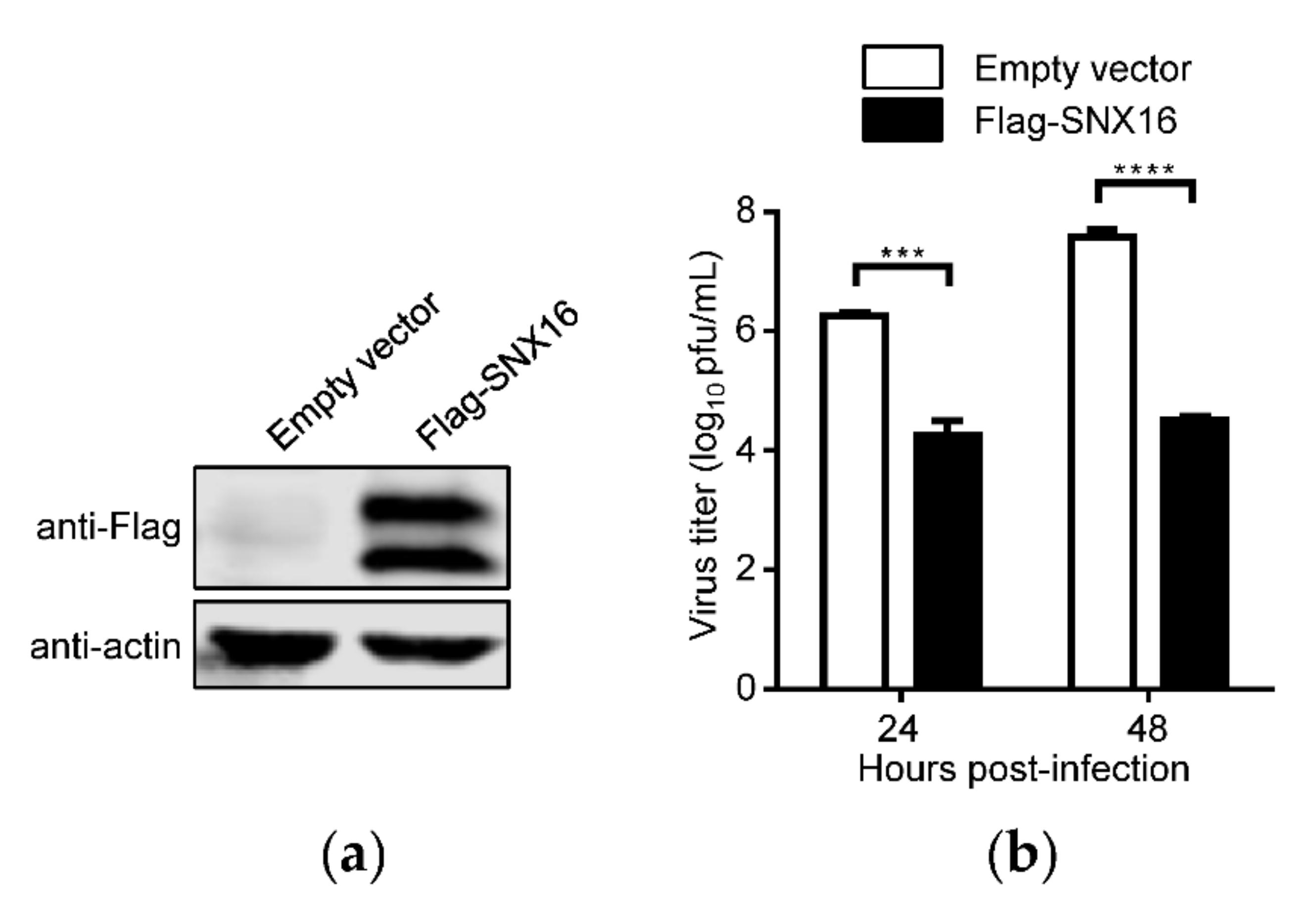{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Plasmids
2.3. Antibodies
2.4. siRNA Knockdown, Plasmid Transfection and Virus Infection
2.5. qRT-PCR Assays
2.6. Western Blotting
2.7. Cell Viability Assay
2.8. Confocal Microscopy
2.9. Plaque Assay
3. Results
3.1. Overexpression of SNX16 Inhibits the Replication of IAV
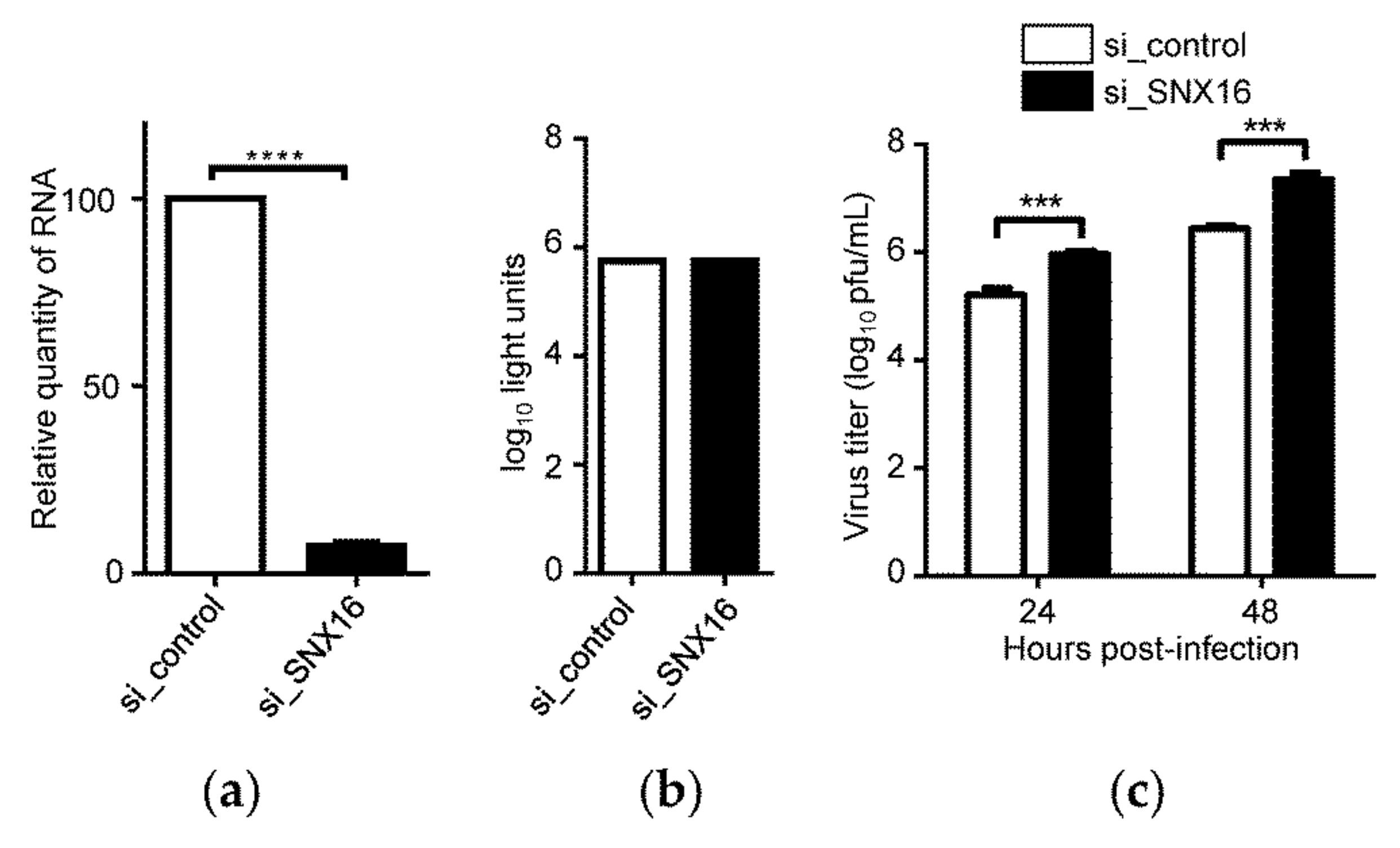
3.2. Downregulation of SNX16 Expression Promotes the Replication of IAV
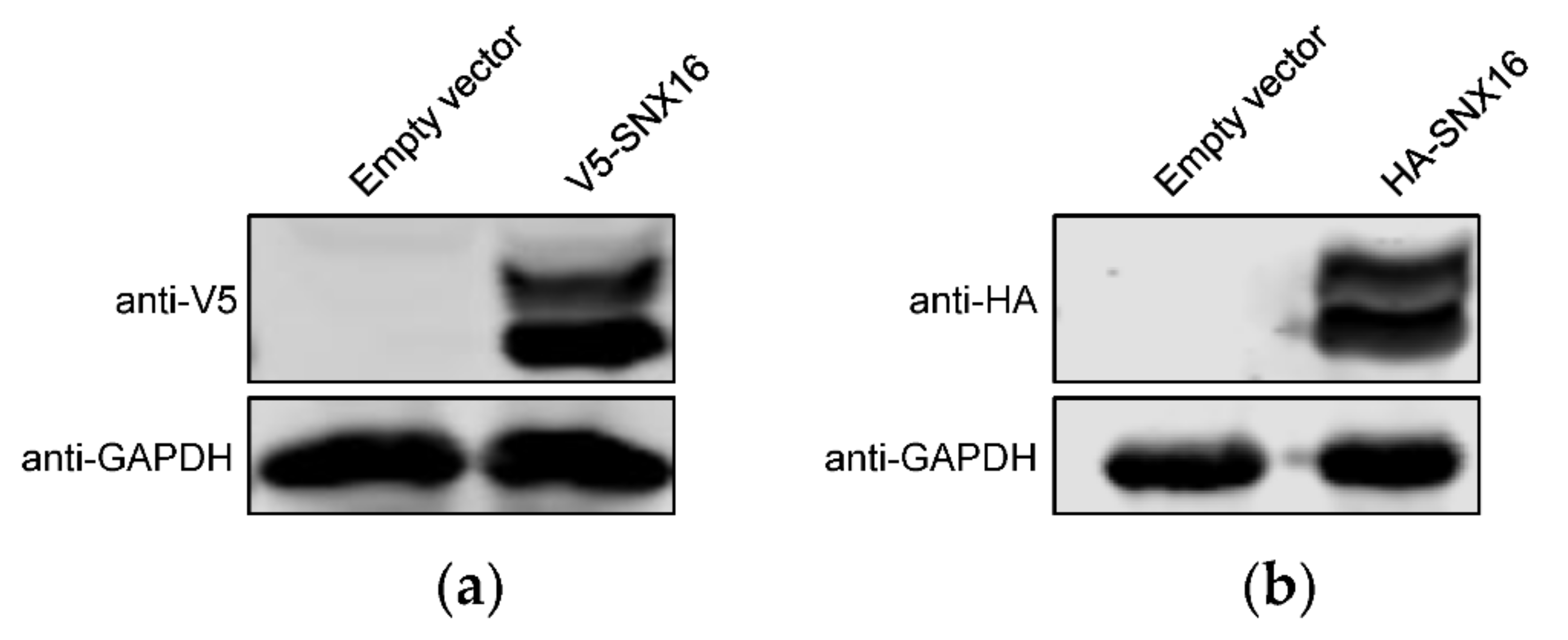
3.3. Transient Overexpression of SNX16 Exhibits Two Obvious Bands
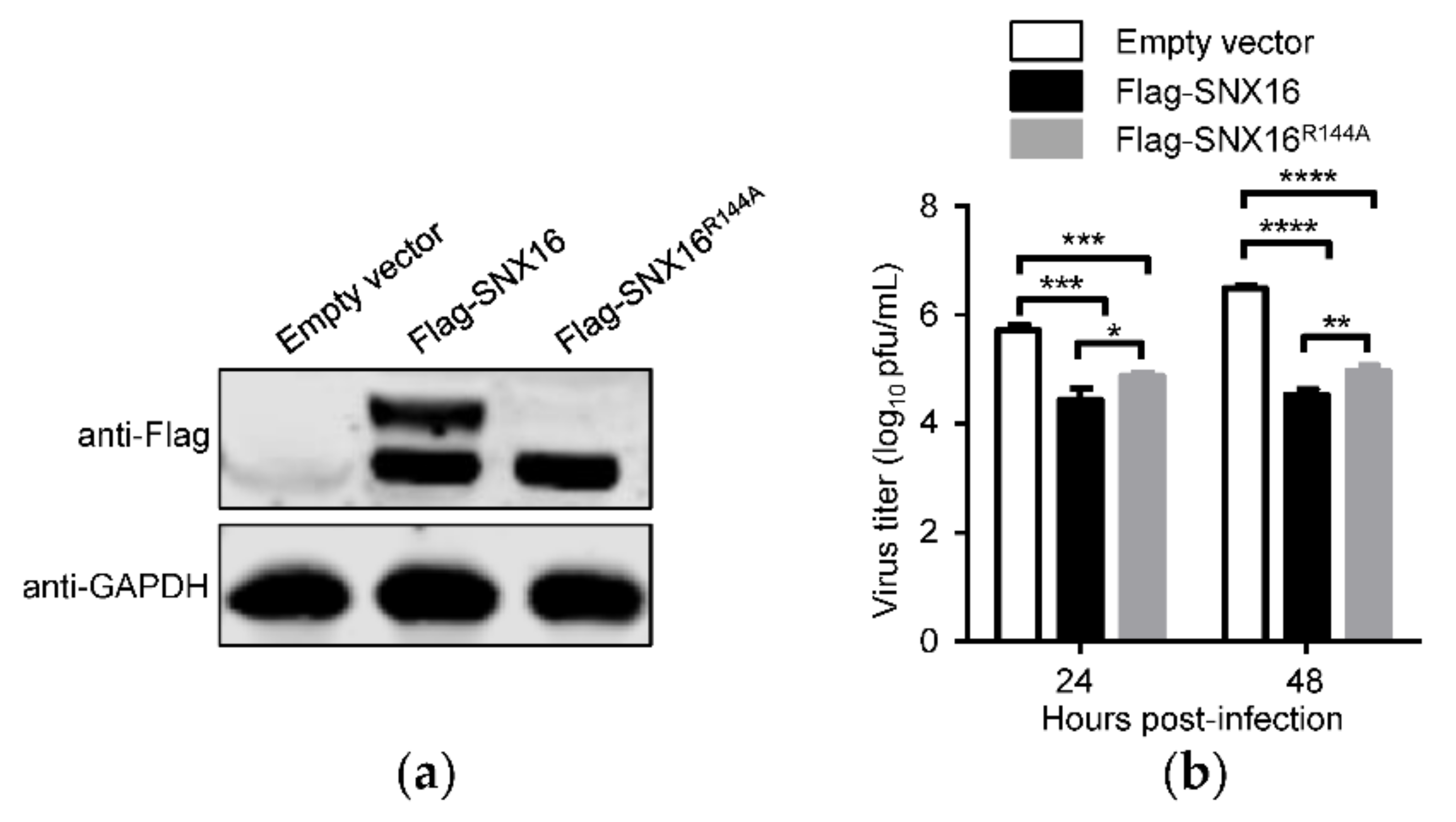
3.4. A Single Amino Acid Residue R144 Is Critical for the Expression and Distribution of SNX16
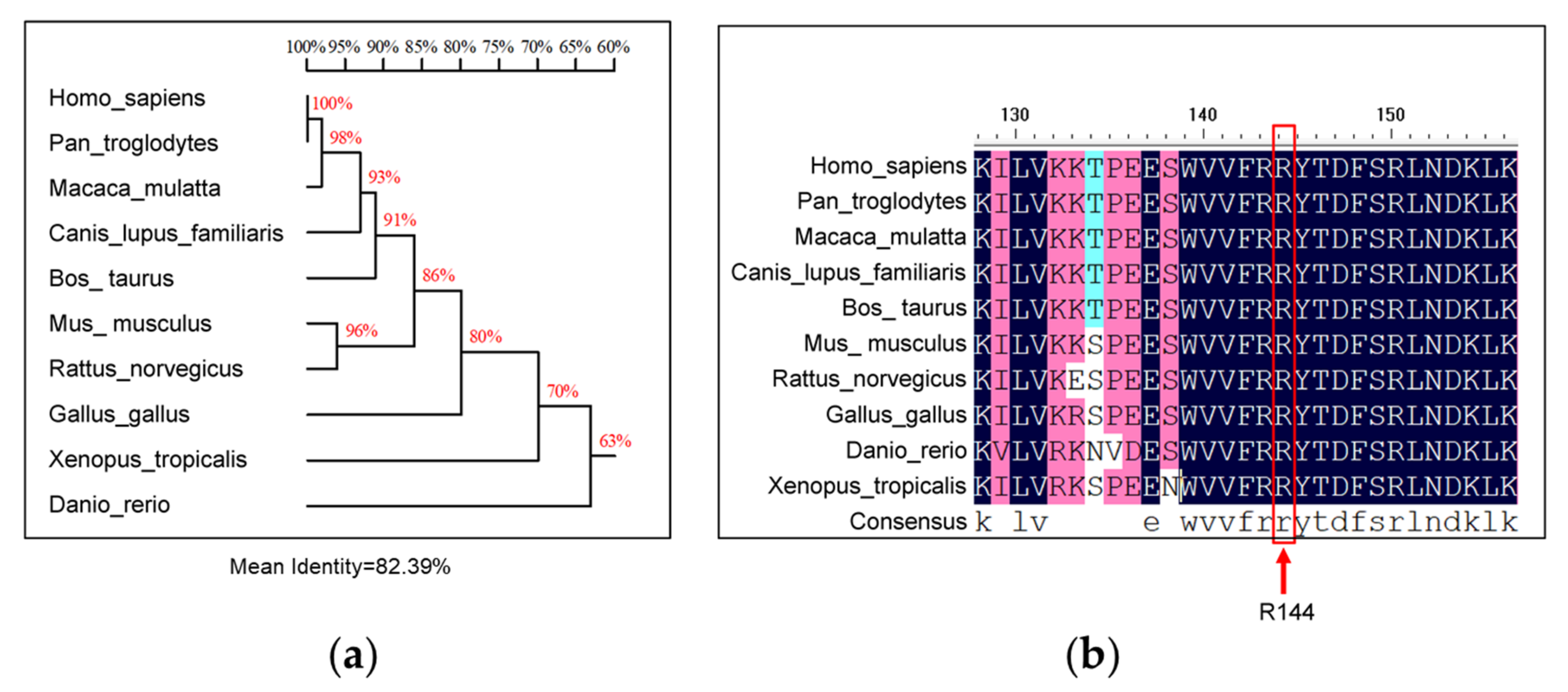
3.5. Homology Analysis of R144 of SNX16
3.6. R144A Mutation of SNX16 Partially Attenuates Its Inhibitory Effect on IAV Replication
3.7. SNX16 Functions in the Early Stage of IAV Replication Cycle
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hilleman, M.R. Realities and enigmas of human viral influenza: Pathogenesis, epidemiology and control. Vaccine 2002, 20, 3068–3087. [Google Scholar] [CrossRef]
- Hause, B.M.; Collin, E.A.; Liu, R.; Huang, B.; Sheng, Z.; Lu, W.; Wang, D.; Nelson, E.A.; Li, F. Characterization of a novel influenza virus in cattle and Swine: Proposal for a new genus in the Orthomyxoviridae family. mBio 2014, 5, e00031-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, A.M.Q.; Lefkowitz, E.J.; Mushegian, A.R.; Adams, M.J.; Dutilh, B.E.; Gorbalenya, A.E.; Harrach, B.; Harrison, R.L.; Junglen, S.; Knowles, N.J.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2018). Arch. Virol. 2018, 163, 2601–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchinson, E.C. Influenza Virus. Trends Microbiol. 2018, 26, 809–810. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, V.N.; Russell, C.A. The evolution of seasonal influenza viruses. Nat. Rev. Microbiol. 2018, 16, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Bu, Z.; Chen, H. Avian influenza vaccines against H5N1 ’bird flu’. Trends Biotechnol. 2014, 32, 147–156. [Google Scholar] [CrossRef]
- Arranz, R.; Coloma, R.; Chichon, F.J.; Conesa, J.J.; Carrascosa, J.L.; Valpuesta, J.M.; Ortin, J.; Martin-Benito, J. The structure of native influenza virion ribonucleoproteins. Science 2012, 338, 1634–1637. [Google Scholar] [CrossRef]
- Moeller, A.; Kirchdoerfer, R.N.; Potter, C.S.; Carragher, B.; Wilson, I.A. Organization of the influenza virus replication machinery. Science 2012, 338, 1631–1634. [Google Scholar] [CrossRef] [Green Version]
- Pleschka, S.; Jaskunas, R.; Engelhardt, O.G.; Zurcher, T.; Palese, P.; Garcia-Sastre, A. A plasmid-based reverse genetics system for influenza A virus. J. Virol. 1996, 70, 4188–4192. [Google Scholar] [CrossRef] [Green Version]
- Neumann, G.; Watanabe, T.; Ito, H.; Watanabe, S.; Goto, H.; Gao, P.; Hughes, M.; Perez, D.R.; Donis, R.; Hoffmann, E.; et al. Generation of influenza A viruses entirely from cloned cDNAs. Proc. Natl. Acad. Sci. USA 1999, 96, 9345–9350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matlin, K.S.; Reggio, H.; Helenius, A.; Simons, K. Infectious entry pathway of influenza virus in a canine kidney cell line. J. Cell Biol. 1981, 91, 601–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vries, E.; Tscherne, D.M.; Wienholts, M.J.; Cobos-Jimenez, V.; Scholte, F.; Garcia-Sastre, A.; Rottier, P.J.; de Haan, C.A. Dissection of the influenza A virus endocytic routes reveals macropinocytosis as an alternative entry pathway. PLoS Pathog. 2011, 7, e1001329. [Google Scholar] [CrossRef] [PubMed]
- White, J.; Kartenbeck, J.; Helenius, A. Membrane fusion activity of influenza virus. EMBO J. 1982, 1, 217–222. [Google Scholar] [CrossRef]
- Miyake, Y.; Keusch, J.J.; Decamps, L.; Ho-Xuan, H.; Iketani, S.; Gut, H.; Kutay, U.; Helenius, A.; Yamauchi, Y. Influenza virus uses transportin 1 for vRNP debundling during cell entry. Nat. Microbiol. 2019, 4, 578–586. [Google Scholar] [CrossRef]
- Lakadamyali, M.; Rust, M.J.; Zhuang, X. Endocytosis of influenza viruses. Microb. Infect. 2004, 6, 929–936. [Google Scholar] [CrossRef] [Green Version]
- Tian, G.; Zhang, S.; Li, Y.; Bu, Z.; Liu, P.; Zhou, J.; Li, C.; Shi, J.; Yu, K.; Chen, H. Protective efficacy in chickens, geese and ducks of an H5N1-inactivated vaccine developed by reverse genetics. Virology 2005, 341, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Chen, P.; Liu, L.; Deng, G.; Li, Y.; Shi, J.; Kong, H.; Feng, H.; Bai, J.; Li, X.; et al. Protective Efficacy of an H5N1 Inactivated Vaccine Against Challenge with Lethal H5N1, H5N2, H5N6, and H5N8 Influenza Viruses in Chickens. Avian Dis. 2016, 60, 253–255. [Google Scholar] [CrossRef]
- Sano, K.; Ainai, A.; Suzuki, T.; Hasegawa, H. The road to a more effective influenza vaccine: Up to date studies and future prospects. Vaccine 2017, 35, 5388–5395. [Google Scholar] [CrossRef]
- Astrahan, P.; Arkin, I.T. Resistance characteristics of influenza to amino-adamantyls. Biochim. Biophys. Acta 2011, 1808, 547–553. [Google Scholar] [CrossRef] [Green Version]
- Samson, M.; Pizzorno, A.; Abed, Y.; Boivin, G. Influenza virus resistance to neuraminidase inhibitors. Antiv. Res. 2013, 98, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.C.; Kumar, G.; Barman, S.; Najera, I.; White, S.W.; Webby, R.J.; Govorkova, E.A. Identification of the I38T PA Substitution as a Resistance Marker for Next-Generation Influenza Virus Endonuclease Inhibitors. mBio 2018, 9, e00430-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.H.; Hong, W.P.; Kim, M.J.; Kim, J.H.; Ryu, S.H.; Suh, P.G. Sorting nexin 16 regulates EGF receptor trafficking by phosphatidylinositol-3-phosphate interaction with the Phox domain. J. Cell Sci. 2004, 117, 4209–4218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhao, Z.; Rodal, A.A. Higher-order assembly of Sorting Nexin 16 controls tubulation and distribution of neuronal endosomes. J. Cell Biol. 2019, 218, 2600–2618. [Google Scholar] [CrossRef] [Green Version]
- Hanson, B.J.; Hong, W. Evidence for a role of SNX16 in regulating traffic between the early and later endosomal compartments. J. Biol. Chem. 2003, 278, 34617–34630. [Google Scholar] [CrossRef] [Green Version]
- Brankatschk, B.; Pons, V.; Parton, R.G.; Gruenberg, J. Role of SNX16 in the dynamics of tubulo-cisternal membrane domains of late endosomes. PLoS ONE 2011, 6, e21771. [Google Scholar] [CrossRef] [Green Version]
- Rodal, A.A.; Blunk, A.D.; Akbergenova, Y.; Jorquera, R.A.; Buhl, L.K.; Littleton, J.T. A presynaptic endosomal trafficking pathway controls synaptic growth signaling. J. Cell Biol. 2011, 193, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Qin, D.; Hao, C.; Shu, X.; Pei, D. SNX16 negatively regulates the migration and tumorigenesis of MCF-7 cells. Cell Regen. 2013, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Li, Y.; Fang, Y.; Lin, M.; Feng, X.; Li, Z.; Zhan, Y.; Liu, Y.; Mou, T.; Lan, X.; et al. SNX16 activates c-Myc signaling by inhibiting ubiquitin-mediated proteasomal degradation of eEF1A2 in colorectal cancer development. Mol. Oncol. 2020, 14, 387–406. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wen, X.; Li, Q.; Li, J.; Wang, G.; Liang, L.; Wang, X.; Chen, H.; Li, C. Generation and application of two monoclonal antibodies targeting conserved linear epitopes in the NP protein of influenza A virus. J. Integr. Agric. 2021. [Google Scholar] [CrossRef]
- Zhu, P.; Liang, L.; Shao, X.; Luo, W.; Jiang, S.; Zhao, Q.; Sun, N.; Zhao, Y.; Li, J.; Wang, J.; et al. Host Cellular Protein TRAPPC6ADelta Interacts with Influenza A Virus M2 Protein and Regulates Viral Propagation by Modulating M2 Trafficking. J. Virol. 2017, 91, e01757-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Zhang, J.; Liang, L.; Wang, G.; Li, Q.; Zhu, P.; Zhou, Y.; Li, J.; Zhao, Y.; Sun, N.; et al. Phospholipid scramblase 1 interacts with influenza A virus NP, impairing its nuclear import and thereby suppressing virus replication. PLoS Pathog. 2018, 14, e1006851. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, I.; Luyet, P.P.; Pons, V.; Ferguson, C.; Emans, N.; Petiot, A.; Mayran, N.; Demaurex, N.; Faure, J.; Sadoul, R.; et al. Endosome-to-cytosol transport of viral nucleocapsids. Nat. Cell Biol. 2005, 7, 653–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, G.; Chen, H.; Gao, G.F.; Shu, Y.; Kawaoka, Y. H5N1 influenza viruses: Outbreaks and biological properties. Cell Res. 2010, 20, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, W.; Li, X.; Goraya, M.U.; Wang, S.; Chen, J.L. Evolution of Influenza A Virus by Mutation and Re-Assortment. Int. J. Mol. Sci. 2017, 18, 1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Deng, G.; Kong, H.; Gu, C.; Ma, S.; Yin, X.; Zeng, X.; Cui, P.; Chen, Y.; Yang, H.; et al. H7N9 virulent mutants detected in chickens in China pose an increased threat to humans. Cell Res. 2017, 27, 1409–1421. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Deng, G.; Ma, S.; Zeng, X.; Yin, X.; Li, M.; Zhang, B.; Cui, P.; Chen, Y.; Yang, H.; et al. Rapid Evolution of H7N9 Highly Pathogenic Viruses that Emerged in China in 2017. Cell Host Microbe 2018, 24, 558–568.e7. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Chen, H. Enhancement of influenza virus transmission by gene reassortment. Curr. Top. Microbiol. Immunol. 2014, 385, 185–204. [Google Scholar] [CrossRef]
- Schotsaert, M.; Garcia-Sastre, A. Influenza vaccines: A moving interdisciplinary field. Viruses 2014, 6, 3809–3826. [Google Scholar] [CrossRef] [Green Version]
- Tenforde, M.W.; Kondor, R.J.G.; Chung, J.R.; Zimmerman, R.K.; Nowalk, M.P.; Jackson, M.L.; Jackson, L.A.; Monto, A.S.; Martin, E.T.; Belongia, E.A.; et al. Effect of Antigenic Drift on Influenza Vaccine Effectiveness in the United States-2019–2020. Clin. Infect. Dis. 2021, 73, e4244–e4250. [Google Scholar] [CrossRef]
- Gouma, S.; Weirick, M.; Hensley, S.E. Potential Antigenic Mismatch of the H3N2 Component of the 2019 Southern Hemisphere Influenza Vaccine. Clin. Infect. Dis. 2020, 70, 2432–2434. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Jiang, L.; Wang, J.; Zhang, J.; Kong, F.; Li, Q.; Yan, Y.; Huang, S.; Zhao, Y.; Liang, L.; et al. The G Protein-Coupled Receptor FFAR2 Promotes Internalization during Influenza A Virus Entry. J. Virol. 2020, 94, e01707-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Huang, H.; Hu, Y.; Zhang, J.; Li, F.; Yin, X.; Shi, J.; Li, Y.; Li, C.; Zhao, D.; et al. A genome-wide CRISPR/Cas9 gene knockout screen identifies immunoglobulin superfamily DCC subclass member 4 as a key host factor that promotes influenza virus endocytosis. PLoS Pathog. 2021, 17, e1010141. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Sun, E.; Bujny, M.V.; Kim, D.; Davidson, M.W.; Zhuang, X. Dual function of CD81 in influenza virus uncoating and budding. PLoS Pathog. 2013, 9, e1003701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edinger, T.O.; Pohl, M.O.; Yanguez, E.; Stertz, S. Cathepsin W Is Required for Escape of Influenza A Virus from Late Endosomes. mBio 2015, 6, e00297. [Google Scholar] [CrossRef] [Green Version]
- Gschweitl, M.; Ulbricht, A.; Barnes, C.A.; Enchev, R.I.; Stoffel-Studer, I.; Meyer-Schaller, N.; Huotari, J.; Yamauchi, Y.; Greber, U.F.; Helenius, A.; et al. A SPOPL/Cullin-3 ubiquitin ligase complex regulates endocytic trafficking by targeting EPS15 at endosomes. eLife 2016, 5, e13841. [Google Scholar] [CrossRef] [Green Version]
- Su, W.C.; Chen, Y.C.; Tseng, C.H.; Hsu, P.W.; Tung, K.F.; Jeng, K.S.; Lai, M.M. Pooled RNAi screen identifies ubiquitin ligase Itch as crucial for influenza A virus release from the endosome during virus entry. Proc. Natl. Acad. Sci. USA 2013, 110, 17516–17521. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, W.; Jiang, L.; Ye, M.; Wang, B.; Chang, Y.; Shan, Z.; Wang, X.; Hu, Y.; Chen, H.; Li, C. A Single Amino Acid Residue R144 of SNX16 Affects Its Ability to Inhibit the Replication of Influenza A Virus. Viruses 2022, 14, 825. https://doi.org/10.3390/v14040825
Shi W, Jiang L, Ye M, Wang B, Chang Y, Shan Z, Wang X, Hu Y, Chen H, Li C. A Single Amino Acid Residue R144 of SNX16 Affects Its Ability to Inhibit the Replication of Influenza A Virus. Viruses. 2022; 14(4):825. https://doi.org/10.3390/v14040825
Chicago/Turabian StyleShi, Wenjun, Li Jiang, Miaomiao Ye, Bo Wang, Yu Chang, Zhibo Shan, Xuyuan Wang, Yuzhen Hu, Hualan Chen, and Chengjun Li. 2022. "A Single Amino Acid Residue R144 of SNX16 Affects Its Ability to Inhibit the Replication of Influenza A Virus" Viruses 14, no. 4: 825. https://doi.org/10.3390/v14040825
APA StyleShi, W., Jiang, L., Ye, M., Wang, B., Chang, Y., Shan, Z., Wang, X., Hu, Y., Chen, H., & Li, C. (2022). A Single Amino Acid Residue R144 of SNX16 Affects Its Ability to Inhibit the Replication of Influenza A Virus. Viruses, 14(4), 825. https://doi.org/10.3390/v14040825