
Differential Transmission of Old and New World Begomoviruses by Middle East-Asia Minor 1 (MEAM1) and Mediterranean (MED) Cryptic Species of Bemisia tabaci
 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants, Virus Inoculum Sources, and B. tabaci
2.2. Bemisia tabaci Fitness on Tomato, Squash, and Prickly Sida Plants
2.3. Virus Detection
2.4. Virus Quantitation
2.5. TYLCV, CuLCrV, or SiGMV Transmission by B. tabaci MEAM1 and B. tabaci MED
2.6. Retention of TYLCV, CuLCrV, or SiGMV by B. tabaci MEAM1 and B. tabaci MED
2.7. TYLCV, CuLCrV, or SiGMV Localization and Accumulation in B. tabaci MEAM1 and B. tabaci MED Tissues
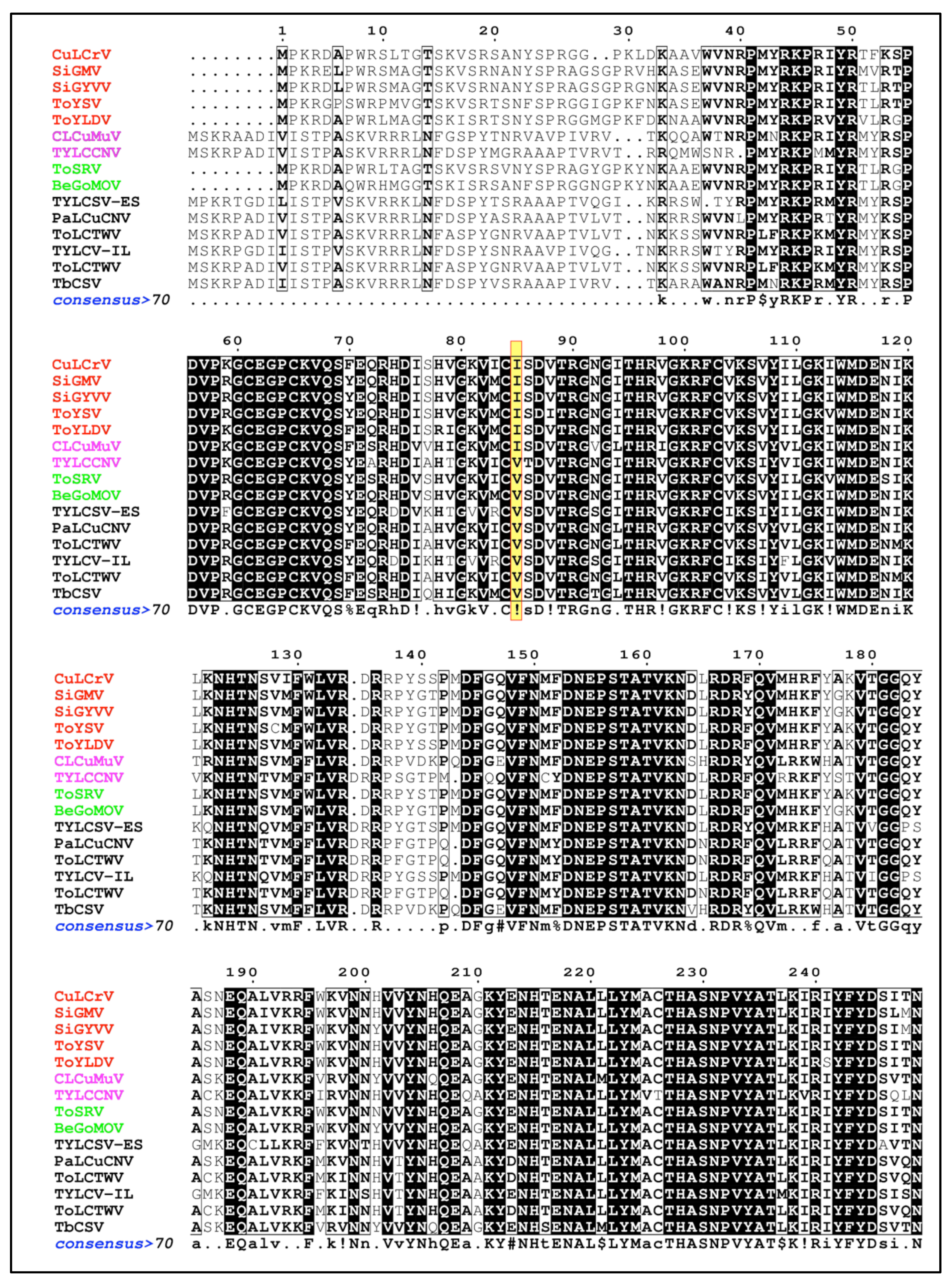
2.8. Comparison of Capsid Protein Sequences of B. tabaci MED Transmissible and Non-Transmissible Begomoviruses
2.9. Statistical Analyses
3. Results
3.1. Bemisia tabaci Performance on Tomato, Squash, or Prickly Sida Plants
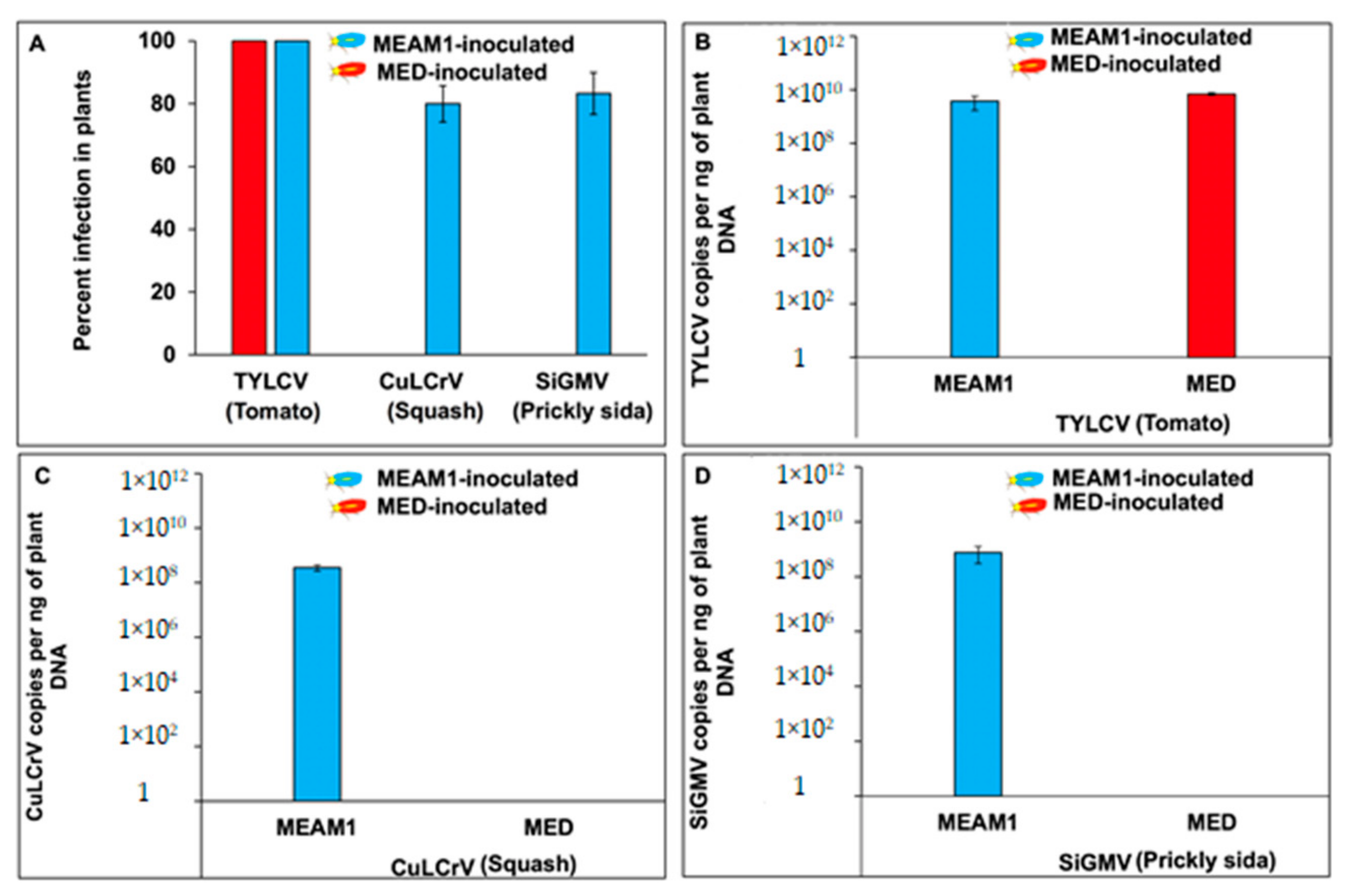
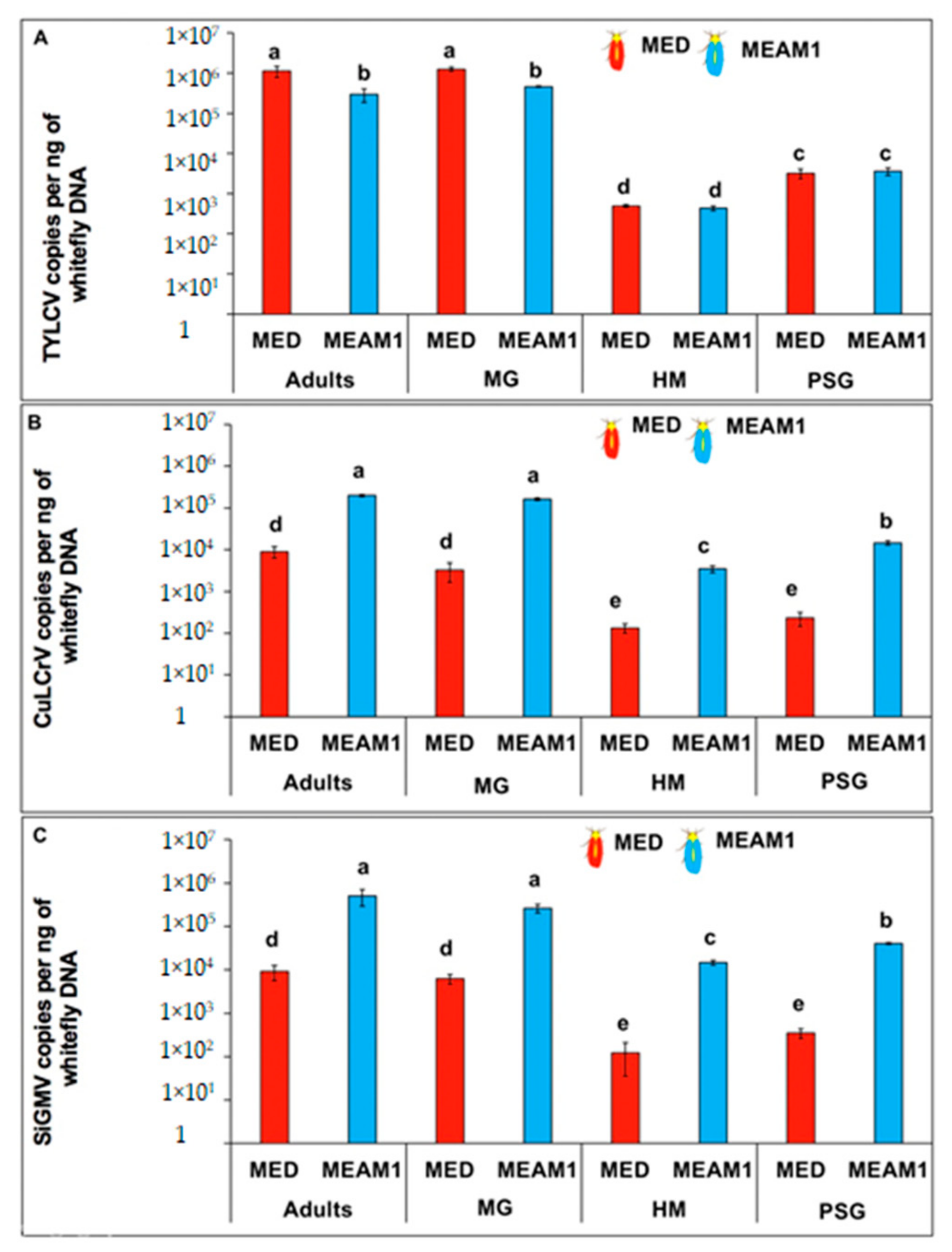
3.2. TYLCV, CuLCrV, or SiGMV Transmission and Quantitation
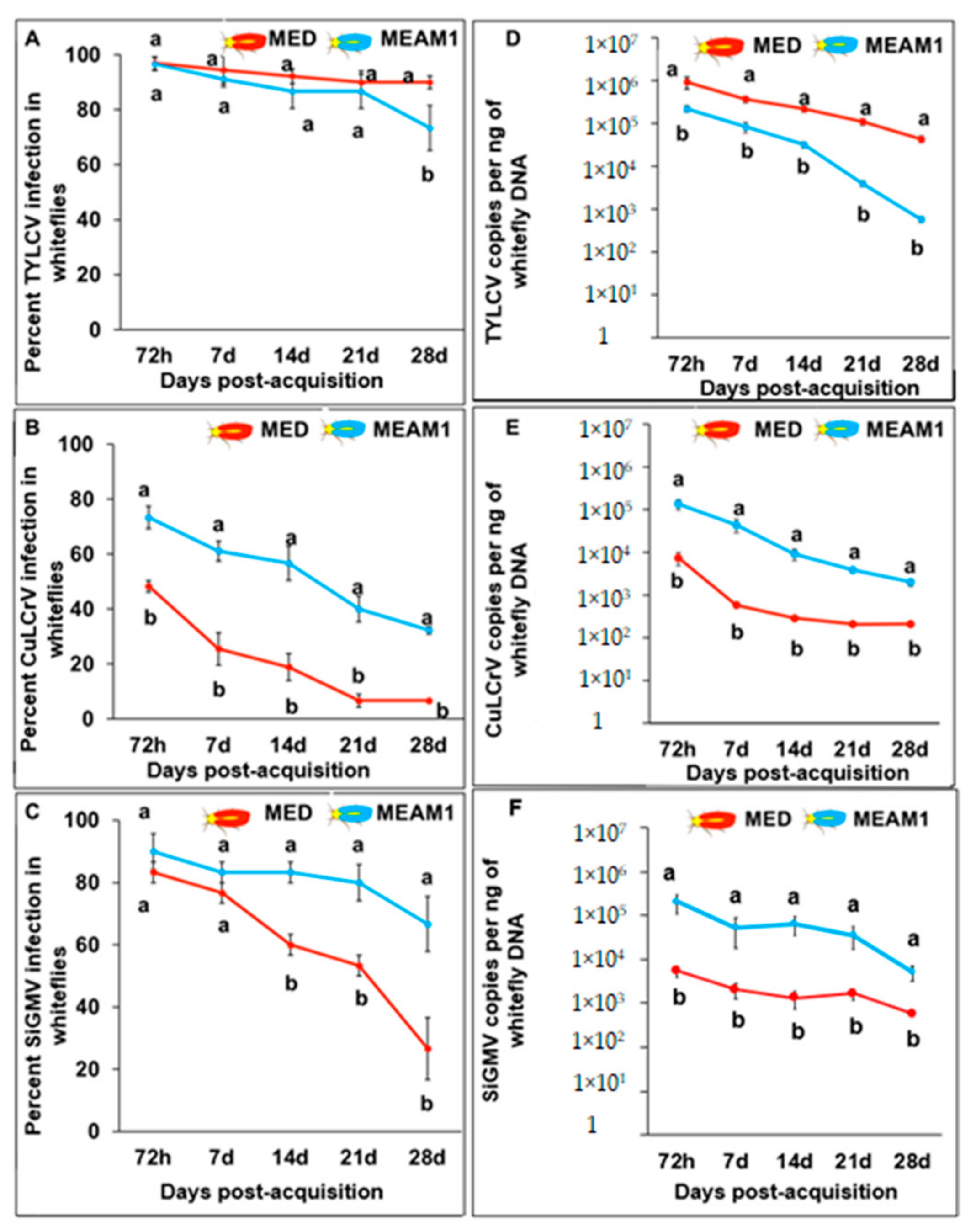
3.3. Retention of TYLCV, CuLCrV, or SiGMV by B. tabaci MEAM1 and B. tabaci MED
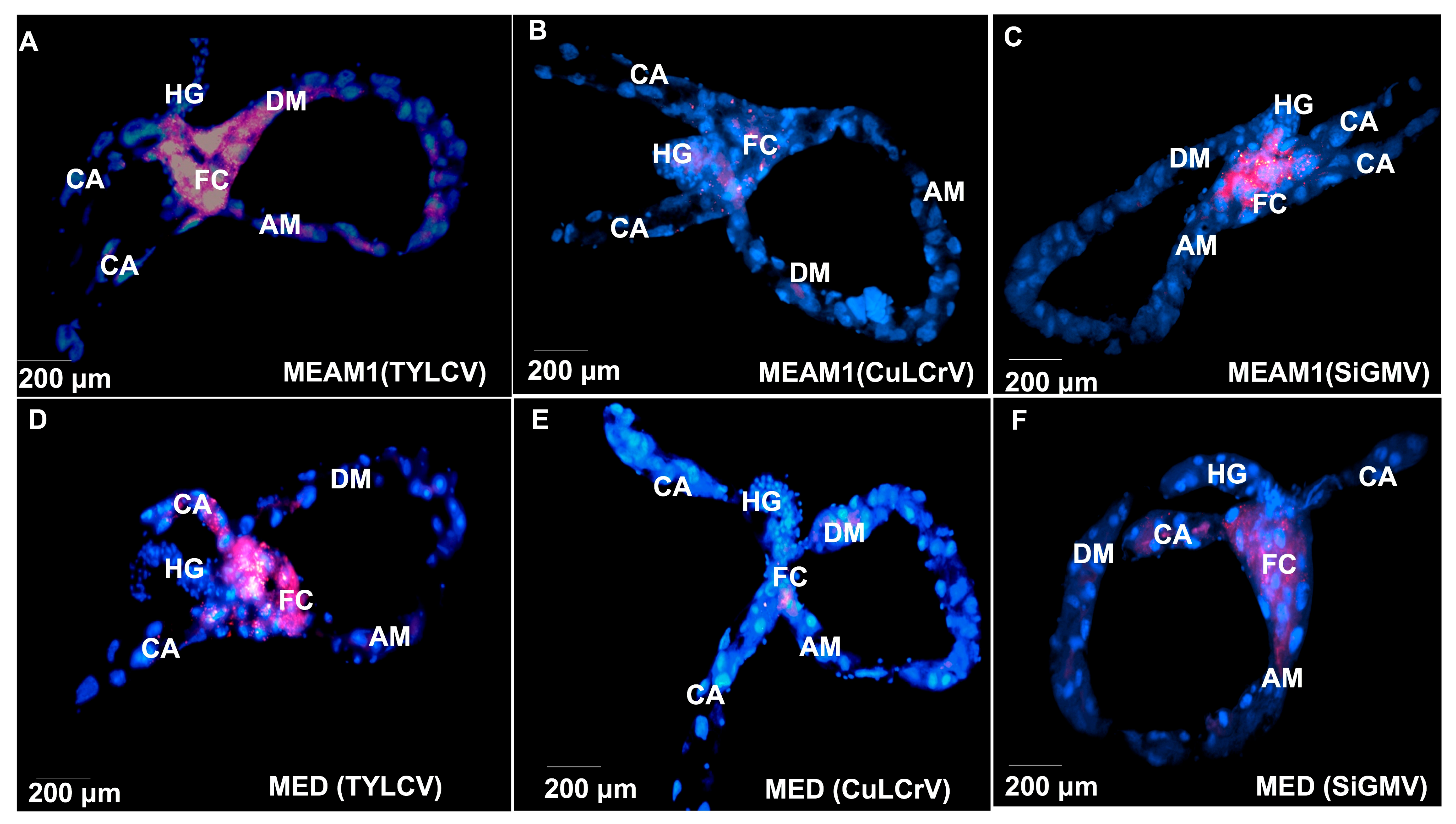
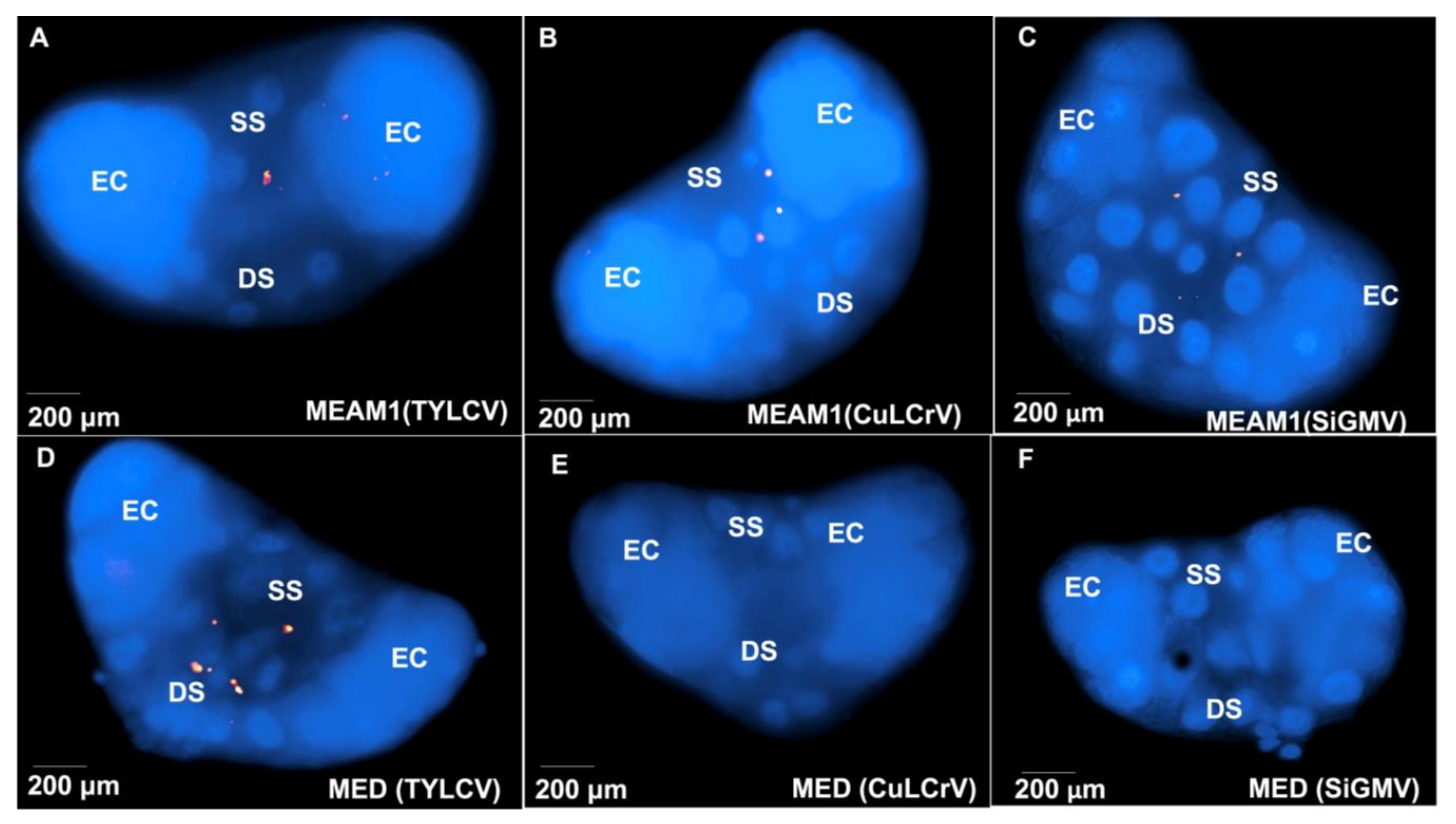
3.4. TYLCV, CuLCrV, or SiGMV Localization and Accumulation in B. tabaci MEAM1 and B. tabaci MED Tissues
3.5. Comparison of Capsid Protein Sequences of B. tabaci MED Transmissible and Non-Transmissible Begomoviruses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aukema, J.E.; Leung, B.; Kovacs, K.; Chivers, C.; Britton, K.O.; Englin, J.; Frankel, S.J.; Haight, R.G.; Holmes, T.P.; Liebhold, A.M.; et al. Economic Impacts of Non-Native Forest Insects in the Continental United States. PLoS ONE 2011, 6, e24587. [Google Scholar] [CrossRef] [PubMed]
- Liebhold, A.M.; Brockerhoff, E.G.; Garrett, L.J.; Parke, J.L.; Britton, K.O. Live plant imports: The major pathway for forest insect and pathogen invasions of the US. Front. Ecol. Environ. 2012, 10, 135–143. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-S.; De Barro, P.J.; Xu, J.; Luan, J.-B.; Zang, L.-S.; Ruan, Y.-M.; Wan, F.-H. Asymmetric Mating Interactions Drive Widespread Invasion and Displacement in a Whitefly. Science 2007, 318, 1769–1772. [Google Scholar] [CrossRef]
- Liansheng, Z.; Shusheng, L.; Yinquan, L.; Yongming, R.; Fanghao, W. Competition between the B biotype and a non-B biotype of the whitefly, Bemisia tabaci (Homoptera: Aleyrodidae) in Zhejiang, China. Chin. Biodivers. 2005, 13, 181–187. [Google Scholar]
- Pan, H.; Chu, D.; Yan, W.; Su, Q.; Liu, B.; Wang, S.; Wu, Q.; Xie, W.; Jiao, X.; Li, R.; et al. Rapid Spread of Tomato Yellow Leaf Curl Virus in China Is Aided Differentially by Two Invasive Whiteflies. PLoS ONE 2012, 7, e34817. [Google Scholar] [CrossRef] [Green Version]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef]
- Bar-Joseph, M.; Marcus, R.; Lee, R.F. The Continuous Challenge of Citrus Tristeza Virus Control. Annu. Rev. Phytopathol. 1989, 27, 291–316. [Google Scholar] [CrossRef]
- McClean, A.P. Triesteza virus complex: Its transmission by the aphid, Toxoptera citricidus. Phytophylactica 1975, 7, 109–114. [Google Scholar]
- Dennehy, T.; Degain, B.; Harpold, V.; Brown, J.; Morin, S.; Fabrick, J.; Byrne, F.; Nichols, R.L. New challenges to management of whitefly resistance to insecticides in Arizona. Univ. Ariz.—Coop. Ext. Veg. Rep. 2005, 10150, 144. [Google Scholar]
- McKenzie, C.L.; Bethke, J.A.; Byrne, F.J.; Chamberlin, J.R.; Dennehy, T.J.; Dickey, A.M.; Gilrein, D.; Hall, P.M.; Ludwig, S.; Oetting, R.D.; et al. Distribution of Bemisia tabaci (Hemiptera: Aleyrodidae) Biotypes in North America After the Q Invasion. J. Econ. Entomol. 2012, 105, 753–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenzie, C.L.; Osborne, L.S. Bemisia tabaci MED (Q biotype) (Hemiptera: Aleyrodidae) in Florida is on the Move to Residential Landscapes and May Impact Open-Field Agriculture. Fla. Entomol. 2017, 100, 481–484. [Google Scholar] [CrossRef] [Green Version]
- Gautam, S.; Crossley, M.S.; Dutta, B.; Coolong, T.; Simmons, A.M.; da Silva, A.; Snyder, W.E.; Srinivasan, R. Low Genetic Variability in Bemisia tabaci MEAM1 Populations within Farmscapes of Georgia, USA. Insects 2020, 11, 834. [Google Scholar] [CrossRef]
- Boykin, L.M.; Shatters, R.G., Jr.; Rosell, R.C.; McKenzie, C.L.; Bagnall, R.A.; De Barro, P.; Frohlich, D.R. Global relationships of Bemisia tabaci (Hemiptera: Aleyrodidae) revealed using Bayesian analysis of mitochondrial COI DNA sequences. Mol. Phylogenet. Evol. 2007, 44, 1306–1319. [Google Scholar] [CrossRef] [PubMed]
- De Barro, P.J.; Liu, S.-S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A Statement of Species Status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef]
- Firdaus, S.; Vosman, B.; Hidayati, N.; Jaya Supena, E.D.; Visser, R.G.F.; van Heusden, A.W. The Bemisia tabaci species complex: Additions from different parts of the world. Insect Sci. 2013, 20, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Marubayashi, J.M.; Yuki, V.A.; Rocha, K.C.G.; Mituti, T.; Pelegrinotti, F.M.; Ferreira, F.Z.; Moura, M.F.; Navas-Castillo, J.; Moriones, E.; Pavan, M.A.; et al. At least two indigenous species of the Bemisia tabaci complex are present in Brazil. J. Appl. Entomol. 2013, 137, 113–121. [Google Scholar] [CrossRef]
- Roopa, H.K.; Asokan, R.; Rebijith, K.B.; Hande, R.H.; Mahmood, R.; Kumar, N.K.K. Prevalence of a New Genetic Group, MEAM-K, of the Whitefly Bemisia tabaci (Hemiptera: Aleyrodidae) in Karnataka, India, as Evident from mtCOI Sequences. Fla. Entomol. 2015, 98, 1062–1071. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Zhang, X.; Jiang, Z.; Zhang, F.; Liu, Y.; Li, Z.; Zhang, Z. New putative cryptic species detection and genetic network analysis of Bemisia tabaci (Hempitera: Aleyrodidae) in China based on mitochondrial COI sequences. Mitochondrial DNA Part A 2018, 29, 474–484. [Google Scholar] [CrossRef]
- Mugerwa, H.; Colvin, J.; Alicai, T.; Omongo, C.A.; Kabaalu, R.; Visendi, P.; Sseruwagi, P.; Seal, S.E. Genetic diversity of whitefly (Bemisia spp.) on crop and uncultivated plants in Uganda: Implications for the control of this devastating pest species complex in Africa. J. Pest. Sci. 2021, 94, 1307–1330. [Google Scholar] [CrossRef]
- Schuster, D.J.; Price, J.F.; Kring, J.B.; Everett, P.H. Integrated Management of the Sweetpotato Whitefly on Commercial Tomato; University of Florida: Gainesville, FL, USA, 1989. [Google Scholar]
- Hoelmer, K.A.; Osborne, L.S.; Yokomi, R.K. Foliage Disorders in Florida Associated with Feeding by Sweetpotato Whitefly, Bemisia tabaci. Florida Entomol. 1991, 74, 162. [Google Scholar] [CrossRef]
- Perring, T.M.; Cooper, A.; Kazmer, D.J. Identification of the Poinsettia Strain of Bemisia tabaci (Homoptera: Aleyrodidae) on Broccoli by Electrophoresis. J. Econ. Entomol. 1992, 85, 1278–1284. [Google Scholar] [CrossRef]
- Perring, T.M.; Cooper, A.; Kazmer, D.J.; Shields, C.; Shields, J. New strain of sweetpotato whitefly invades California vegetables. Calif. Agric. 1991, 45, 10–12. [Google Scholar] [CrossRef] [Green Version]
- Gonzales, R.A.; Godman, G.E.; Natwick, E.T.; Rosenberg, H.; Groeshop, J.I.; Sutter, S.R.; Funakoshi, T.; Davilagarcia, S. Whitefly invasion in Imperial Valley costs growers, workers millions in losses. Calif. Agric. 1992, 46, 7–8. [Google Scholar] [CrossRef]
- Brown, J.K.; Frohlich, D.R.; Rosell, R.C. The Sweetpotato or Silverleaf Whiteflies: Biotypes of Bemisia tabaci or a Species Complex? Annu. Rev. Entomol. 1995, 40, 511–534. [Google Scholar] [CrossRef]
- Morales, F.J.; Anderson, P.K. The emergence and dissemination of whitefly-transmitted geminiviruses in Latin America. Arch. Virol. 2001, 146, 415–441. [Google Scholar] [CrossRef]
- Simone, G.W.; Brown, J.K.; Hiebert, E.; Cullen, R. Geminiviruses associated with epidemics in Florida tomatoes and pepper. Phytopathology 1990, 80, 1063–1070. [Google Scholar]
- Isakeit, T. First Report of Squash Leaf Curl Virus on Watermelon in Texas. Plant Dis. 1994, 78, 1010D. [Google Scholar] [CrossRef]
- Chu, D.; Zhang, Y.-J.; Brown, J.K.; Cong, B.; Xu, B.-Y.; Wu, Q.-J.; Zhu, G.-R. The Introduction of the Exotic Q Biotype of Bemisia tabaci from the Mediterranean Region into China on Ornamental Crops. Florida Entomol. 2006, 89, 168–174. [Google Scholar] [CrossRef]
- Teng, X.; Wan, F.-H.; Chu, D. Bemisia tabaci Biotype Q Dominates Other Biotypes Across China. Florida Entomol. 2010, 93, 363–368. [Google Scholar] [CrossRef]
- Tang, X.-T.; Cai, L.; Shen, Y.; Xu, L.-L.; Du, Y.-Z. Competitive Displacement between Bemisia tabaci MEAM1 and MED and Evidence for Multiple Invasions of MED. Insects 2019, 11, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, M.R.; Hagen, C.; Lucas, W.J.; Gilbertson, R.L. Exploiting Chinks in the Plant’s Armor: Evolution and Emergence of Geminiviruses. Annu. Rev. Phytopathol. 2005, 43, 361–394. [Google Scholar] [CrossRef] [PubMed]
- Bedford, I.D.; Briddon, R.W.; Brown, J.K.; Rosell, R.C.; Markham, P.G. Geminivirus transmission and biological characterisation of Bemisia tabaci (Gennadius) biotypes from different geographic regions. Ann. Appl. Biol. 1994, 125, 311–325. [Google Scholar] [CrossRef]
- Inoue-Nagata, A.K.; Lima, M.F.; Gilbertson, R.L. A review of geminivirus diseases in vegetables and other crops in Brazil: Current status and approaches for management. Hortic. Bras. 2016, 34, 8–18. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.R. Plant Viruses Transmitted by Whiteflies. Eur. J. Plant Pathol. 2003, 109, 196–219. [Google Scholar] [CrossRef]
- Ellsworth, P.C.; Martinez-Carrillo, J.L. IPM for Bemisia tabaci: A case study from North America. Crop. Prot. 2001, 20, 853–869. [Google Scholar] [CrossRef]
- Palumbo, J.; Horowitz, A.; Prabhaker, N. Insecticidal control and resistance management for Bemisia tabaci. Crop. Prot. 2001, 20, 739–765. [Google Scholar] [CrossRef]
- Elbert, A.; Nauen, R. Resistance of Bemisia tabaci (Homoptera: Aleyrodidae) to insecticides in southern Spain with special reference to neonicotinoids. Pest. Manag. Sci. 2000, 56, 60–64. [Google Scholar] [CrossRef]
- Horowitz, A.R.; Denholm, I.; Gorman, K.; Cenis, J.; Kontsedalov, S.; Ishaaya, I. Biotype Q of Bemisia tabaci identified in Israel. Phytoparasitica 2003, 31, 94–98. [Google Scholar] [CrossRef]
- Pascual, S.; Callejas, C. Intra- and interspecific competition between biotypes B and Q of Bemisia tabaci (Hemiptera: Aleyrodidae) from Spain. Bull. Entomol. Res. 2004, 94, 369–375. [Google Scholar] [CrossRef]
- Stansly, P.A.; McKenzie, C.L. Fourth International Bemisia Workshop International Whitefly Genomics Workshop December 3–8, 2006, Duck Key, FL, USA. J. Insect Sci. 2008, 8, 1–54. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Preisser, E.L.; Chu, D.; Wang, S.; Wu, Q.; Carrière, Y.; Zhou, X.; Zhang, Y. Insecticides promote viral outbreaks by altering herbivore competition. Ecol. Appl. 2015, 25, 1585–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Preisser, E.L.; Chu, D.; Pan, H.; Xie, W.; Wang, S.; Wu, Q.; Zhou, X.; Zhang, Y. Multiple Forms of Vector Manipulation by a Plant-Infecting Virus: Bemisia tabaci and Tomato Yellow Leaf Curl Virus. J. Virol. 2013, 87, 4929–4937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, W.; Shi, X.; Liu, B.; Pan, H.; Wei, W.; Zeng, Y.; Sun, X.; Xie, W.; Wang, S.; Wu, Q.; et al. Transmission of Tomato Yellow Leaf Curl Virus by Bemisia tabaci as Affected by Whitefly Sex and Biotype. Sci. Rep. 2015, 5, 10744. [Google Scholar] [CrossRef] [PubMed]
- Rosen, R.; Kanakala, S.; Kliot, A.; Cathrin Pakkianathan, B.; Farich, B.A.; Santana-Magal, N.; Elimelech, M.; Kontsedalov, S.; Lebedev, G.; Cilia, M.; et al. Persistent, circulative transmission of begomoviruses by whitefly vectors. Curr. Opin. Virol. 2015, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging Virus Diseases Transmitted by Whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef]
- Czosnek, H.; Hariton-Shalev, A.; Sobol, I.; Gorovits, R.; Ghanim, M. The Incredible Journey of Begomoviruses in Their Whitefly Vector. Viruses 2017, 9, 273. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Pan, L.-L.; Liu, S.-S.; Navas-Castillo, J. Transmission of Begomoviruses and Other Whitefly-Borne Viruses: Dependence on the Vector Species. Phytopathology 2020, 110, 10–17. [Google Scholar] [CrossRef]
- Harrison, B.D.; Swanson, M.M.; Fargette, D. Begomovirus coat protein: Serology, variation and functions. Physiol. Mol. Plant Pathol. 2002, 60, 257–271. [Google Scholar] [CrossRef]
- Guo, T.; Zhao, J.; Pan, L.-L.; Geng, L.; Lei, T.; Wang, X.-W.; Liu, S.-S. The level of midgut penetration of two begomoviruses affects their acquisition and transmission by two species of Bemisia tabaci. Virology 2018, 515, 66–73. [Google Scholar] [CrossRef]
- Pan, L.-L.; Cui, X.-Y.; Chen, Q.-F.; Wang, X.-W.; Liu, S.-S. Cotton Leaf Curl Disease: Which Whitefly Is the Vector? Phytopathology 2018, 108, 1172–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Zhao, J.-J.; Zhang, T.; Li, F.-F.; Ghanim, M.; Zhou, X.-P.; Ye, G.-Y.; Liu, S.-S.; Wang, X.-W. Specific Cells in the Primary Salivary Glands of the Whitefly Bemisia tabaci Control Retention and Transmission of Begomoviruses. J. Virol. 2014, 88, 13460–13468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnishi, J.; Kitamura, T.; Terami, F.; Honda, K. A selective barrier in the midgut epithelial cell membrane of the nonvector whitefly Trialeurodes vaporariorum to Tomato yellow leaf curl virus uptake. J. Gen. Plant Pathol. 2009, 75, 131–139. [Google Scholar] [CrossRef]
- Caciagli, P.; Medina Piles, V.; Marian, D.; Vecchiati, M.; Masenga, V.; Mason, G.; Falcioni, T.; Noris, E. Virion Stability Is Important for the Circulative Transmission of Tomato Yellow Leaf Curl Sardinia Virus by Bemisia tabaci, but Virion Access to Salivary Glands Does Not Guarantee Transmissibility. J. Virol. 2009, 83, 5784–5795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Kontsedalov, S.; Skaljac, M.; Brumin, M.; Sobol, I.; Czosnek, H.; Vavre, F.; Fleury, F.; et al. The Transmission Efficiency of Tomato Yellow Leaf Curl Virus by the Whitefly Bemisia tabaci Is Correlated with the Presence of a Specific Symbiotic Bacterium Species. J. Virol. 2010, 84, 9310–9317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polston, J.E.; McGovern, R.J.; Brown, L.G. Introduction of Tomato Yellow Leaf Curl Virus in Florida and Implications for the Spread of This and Other Geminiviruses of Tomato. Plant Dis. 1999, 83, 984–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagen, C.; Rojas, M.R.; Sudarshana, M.R.; Xoconostle-Cazares, B.; Natwick, E.T.; Turini, T.A.; Gilbertson, R.L. Biology and Molecular Characterization of Cucurbit leaf crumple virus, an Emergent Cucurbit-Infecting Begomovirus in the Imperial Valley of California. Plant Dis. 2008, 92, 781–793. [Google Scholar] [CrossRef] [Green Version]
- Gautam, S. The Role of Bemisia tabaci in the Transmission of Vegetable Viruses in the Farmscape of Georgia. Ph.D. Thesis, University of Georgia, Athens, GA, USA, 2019. [Google Scholar]
- Frohlich, D.R.; Torres-Jerez, I.; Bedford, I.D.; Markham, P.G.; Brown, J.K. A phylogeographical analysis of the Bemisia tabaci species complex based on mitochondrial DNA markers. Mol. Ecol. 1999, 8, 1683–1691. [Google Scholar] [CrossRef]
- Mugerwa, H.; Seal, S.; Wang, H.-L.; Patel, M.V.; Kabaalu, R.; Omongo, C.A.; Alicai, T.; Tairo, F.; Ndunguru, J.; Sseruwagi, P.; et al. African ancestry of New World, Bemisia tabaci-whitefly species. Sci. Rep. 2018, 8, 2734. [Google Scholar] [CrossRef] [Green Version]
- Legarrea, S.; Barman, A.; Marchant, W.; Diffie, S.; Srinivasan, R. Temporal Effects of a Begomovirus Infection and Host Plant Resistance on the Preference and Development of an Insect Vector, Bemisia tabaci, and Implications for Epidemics. PLoS ONE 2015, 10, e0142114. [Google Scholar] [CrossRef] [Green Version]
- Gautam, S.; Gadhave, K.R.; Buck, J.W.; Dutta, B.; Coolong, T.; Adkins, S.; Srinivasan, R. Virus-virus interactions in a plant host and in a hemipteran vector: Implications for vector fitness and virus epidemics. Virus Res. 2020, 286, 198069. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Brooks, W.M. Initial Handling and Diagnosis of Diseased Insects; Lacey, L., Ed.; Academic Press: Cambridge, MA, USA, 1997; ISBN 9780080535777. [Google Scholar]
- Rosell, R.C.; Torres-Jerez, I.; Brown, J.K. Tracing the Geminivirus-Whitefly Transmission Pathway by Polymerase Chain Reaction in Whitefly Extracts, Saliva, Hemolymph, and Honeydew. Phytopathology 1999, 89, 239–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kliot, A.; Kontsedalov, S.; Lebedev, G.; Brumin, M.; Cathrin, P.B.; Marubayashi, J.M.; Skaljac, M.; Belausov, E.; Czosnek, H.; Ghanim, M. Fluorescence in situ Hybridizations (FISH) for the Localization of Viruses and Endosymbiotic Bacteria in Plant and Insect Tissues. J. Vis. Exp. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- He, Y.-Z.; Wang, Y.-M.; Yin, T.-Y.; Fiallo-Olivé, E.; Liu, Y.-Q.; Hanley-Bowdoin, L.; Wang, X.-W. A plant DNA virus replicates in the salivary glands of its insect vector via recruitment of host DNA synthesis machinery. Proc. Natl. Acad. Sci. USA 2020, 117, 16928–16937. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Ashton, B.; Buxton, S.; Cheung, M.; Cooper, A.; Duran, C.; Field, M.; Heled, J.; Kearse, M.; Markowitz, S.; et al. Geneious, version Pro v. 8.1.9, Auckland, New Zealand. 2014. Available online: https://www.geneious.com/ (accessed on 24 March 2022).
- Fiallo-Olivé, E.; Tovar, R.; Navas-Castillo, J. Deciphering the biology of deltasatellites from the New World: Maintenance by New World begomoviruses and whitefly transmission. New Phytol. 2016, 212, 680–692. [Google Scholar] [CrossRef] [Green Version]
- Bello, V.H.; Watanabe, L.F.M.; Santos, B.R.; Marubayashi, J.M.; Yuki, V.A.; De Marchi, B.R.; Pavan, M.A.; Krause-Sakate, R. Evidence for increased efficiency of virus transmission by populations of Mediterranean species of Bemisia tabaci with high Hamiltonella prevalence. Phytoparasitica 2019, 47, 293–300. [Google Scholar] [CrossRef]
- Xavier, D.C.A. Begomoviruses: Variability, Evolution, and Interactions with Hosts and Vectors; Universidade Federal de Viçosa: Viçosa, Brazil, 2019. [Google Scholar]
- Pan, L.; Chen, Q.; Guo, T.; Wang, X.; Li, P.; Wang, X.; Liu, S. Differential efficiency of a begomovirus to cross the midgut of different species of whiteflies results in variation of virus transmission by the vectors. Sci. China Life Sci. 2018, 61, 1254–1265. [Google Scholar] [CrossRef]
- Jiang, Y.X.; De Blas, C.; Bedford, I.D.; Nombela, G.; Muñiz, M. Short communication. Effect of Bemisia tabaci biotype in the transmission of Tomato Yellow Leaf Curl Sardinia Virus (TYLCSV-ES) between tomato and common weeds. Spanish J. Agric. Res. 2004, 2, 115. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Saeed, Q.; He, Z.; Lu, L. Transmission efficiency of Cotton leaf curl Multan virus by three cryptic species of Bemisia tabaci complex in cotton cultivars. PeerJ 2019, 7, e7788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, T.; Guo, Q.; Cui, X.-Y.; Liu, Y.-Q.; Hu, J.; Liu, S.-S. Comparison of transmission of Papaya leaf curl China virus among four cryptic species of the whitefly Bemisia tabaci complex. Sci. Rep. 2015, 5, 15432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, S.H.; Tsai, W.S.; Kenyon, L.; Tsai, C.W. Different transmission efficiencies may drive displacement of tomato begomoviruses in the fields in Taiwan. Ann. Appl. Biol. 2015, 166, 321–330. [Google Scholar] [CrossRef]
- Fortes, I.; Sánchez-Campos, S.; Fiallo-Olivé, E.; Díaz-Pendón, J.; Navas-Castillo, J.; Moriones, E. A Novel Strain of Tomato Leaf Curl New Delhi Virus Has Spread to the Mediterranean Basin. Viruses 2016, 8, 307. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Pan, L.-L.; Bouvaine, S.; Fan, Y.-Y.; Liu, Y.-Q.; Liu, S.-S.; Seal, S.; Wang, X.-W. Differential transmission of Sri Lankan cassava mosaic virus by three cryptic species of the whitefly Bemisia tabaci complex. Virology 2020, 540, 141–149. [Google Scholar] [CrossRef]
- Peng, J.; Xie, G.; Zhang, S.; Zheng, L.; Gao, Y.; Zhang, Z.; Luo, L.; Su, P.; Wang, D.; Liu, Y.; et al. Higher Ramie mosaic virus transmission efficiency by females than by males of Bemisia tabaci MED. Sci. Rep. 2020, 10, 525. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed.; SAGE Publications, Inc.: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Idris, A.M.; Smith, S.E.; Brown, J.K. Ingestion, transmission, and persistence of Chino del tomate virus (CdTV), a New World begomovirus, by Old and New World biotypes of the whitefly vector Bemisia tabaci. Ann. Appl. Biol. 2001, 139, 145–154. [Google Scholar] [CrossRef]
- Caciagli, P.; Bosco, D. Quantitation Over Time of Tomato Yellow Leaf Curl Geminivirus DNA in Its Whitefly Vector. Phytopathology 1997, 87, 610–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Q.; Pan, H.; Liu, B.; Chu, D.; Xie, W.; Wu, Q.; Wang, S.; Xu, B.; Zhang, Y. Insect symbiont facilitates vector acquisition, retention and transmission of plant virus. Sci. Rep. 2013, 3, 1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapidot, M.; Friedmann, M.; Pilowsky, M.; Ben-Joseph, R.; Cohen, S. Effect of Host Plant Resistance to Tomato yellow leaf curl virus (TYLCV) on Virus Acquisition and Transmission by Its Whitefly Vector. Phytopathology 2001, 91, 1209–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legarrea, S.; Barman, A.; Diffie, S.; Srinivasan, R. Virus Accumulation and Whitefly Performance Modulate the Role of Alternate Host Species as Inoculum Sources of Tomato Yellow Leaf Curl Virus. Plant Dis. 2020, 104, 2958–2966. [Google Scholar] [CrossRef]
- Kollenberg, M.; Winter, S.; Götz, M. Quantification and Localization of Watermelon Chlorotic Stunt Virus and Tomato Yellow Leaf Curl Virus (Geminiviridae) in Populations of Bemisia tabaci (Hemiptera, Aleyrodidae) with Differential Virus Transmission Characteristics. PLoS ONE 2014, 9, e111968. [Google Scholar] [CrossRef]
- Czosnek, H.; Ghanim, M.; Ghanim, M. The circulative pathway of begomoviruses in the whitefly vector Bemisia tabaci—insights from studies with Tomato yellow leaf curl virus. Ann. Appl. Biol. 2002, 140, 215–231. [Google Scholar] [CrossRef]
- Ban, F.-X.; Yin, T.-Y.; Guo, Q.; Pan, L.-L.; Liu, Y.-Q.; Wang, X.-W. Localization and Quantification of Begomoviruses in Whitefly Tissues by Immunofluorescence and Quantitative PCR. J. Vis. Exp. 2020. [Google Scholar] [CrossRef]
- Pan, L.-L.; Chi, Y.; Liu, C.; Fan, Y.-Y.; Liu, S.-S. Mutations in the coat protein of a begomovirus result in altered transmission by different species of whitefly vectors. Virus Evol. 2020, 6, veaa014. [Google Scholar] [CrossRef]
- Höhnle, M.; Höfer, P.; Bedford, I.D.; Briddon, R.W.; Markham, P.G.; Frischmuth, T. Exchange of Three Amino Acids in the Coat Protein Results in Efficient Whitefly Transmission of a Nontransmissible Abutilon Mosaic Virus Isolate. Virology 2001, 290, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Noris, E.; Vaira, A.M.; Caciagli, P.; Masenga, V.; Gronenborn, B.; Accotto, G.P. Amino Acids in the Capsid Protein of Tomato Yellow Leaf Curl Virus That Are Crucial for Systemic Infection, Particle Formation, and Insect Transmission. J. Virol. 1998, 72, 10050–10057. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Ghanim, M. Factors Determining Transmission of Persistent Viruses by Bemisia tabaci and Emergence of New Virus–Vector Relationships. Viruses 2021, 13, 1808. [Google Scholar] [CrossRef] [PubMed]
- Catto, M.A.; Mugerwa, H.; Myers, B.K.; Pandey, S.; Dutta, B.; Srinivasan, R. A Review on Transcriptional Responses of Interactions between Insect Vectors and Plant Viruses. Cells 2022, 11, 693. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Bouvaine, S.; Maruthi, M.N. Prevalence and Genetic Diversity of Endosymbiotic Bacteria Infecting Cassava Whiteflies in Africa. BMC Microbiol. 2015, 15, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Lei, T.; Zhang, X.-J.; Yin, T.-Y.; Wang, X.-W.; Liu, S.-S. A vector whitefly endocytic receptor facilitates the entry of begomoviruses into its midgut cells via binding to virion capsid proteins. PLoS Pathog. 2020, 16, e1009053. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | GenBank Accession Numbers | Name | Sequence (5′–3′) | Product Size (bp) |
|---|---|---|---|---|
| TYLCV | KY971372 | TYLC-C2-For | GCAGTGATGAGTTCCCCTGT | 102 |
| TYLC-C2-Rev | CCAATAAGGCGTAAGCGTGT | |||
| C2-1159 | CTAGATCCACTGCTCTGATTACA | 694 | ||
| C2-1853 | TCATTGATGACCTAGCAAAG | |||
| CuLCrV | AF224760 | CuLCrV-QF | CCTCAAAGGTTTCCCGCTCT | 110 |
| CuLCrV-QR | CCGATAGATCCTGGGCTTCC | |||
| 3FAC3 | TTTATATCATGATTTTCGAGTACA | 525 | ||
| 5RAC1 | AAAATGAAAGCCTAAGAGAGTGGA | |||
| SiGMV | MK387701 | SiGMV-QF | CTCAAAGGTTAGCCGCAACG | 114 |
| SiGMV-QR | CGGTAGATCCTGGGCTTCCT | |||
| SiGMVF | TTCTCCTCGTGCAGGTAGTG | 574 | ||
| SiGMVR | ACTTGCCAGCCTCTTGATGA |
| Sr. No. | Virus | Transmission Efficiency (No. of Viruliferous Whitefly Used Plant) | Accession Number | Type a | Reference |
|---|---|---|---|---|---|
| 1 | Cucurbit leaf crumple virus (CuLCrV) | 0% (100) | AF256200 | NW (bipartite) | Current study |
| 2 | Sida golden mosaic virus (SiGMV) | 0% (100) | MK387701 | NW (bipartite) | Current study |
| 3 | Sida golden yellow vein virus (SiGYVV) | 0% (50) | HQ822123 | NW (bipartite) | [71] |
| 4 | Bean golden mosaic virus (BGMV) | 100% (10) | NC_004042 | NW (bipartite) | [72] |
| 5 | Tomato severe rugose virus (ToSRV) | 10% (15) | MG837738 | NW (bipartite) | [73] |
| 6 | Tomato yellow spot virus (ToYSV) | 0% (15) | DQ336350 | NW (bipartite) | [73] |
| 7 | Tomato leaf deformation virus (ToYLDV) | 0% (50) | NC_014510 | NW (monopartite) | [71] |
| 8 | Tomato yellow leaf curl China virus (TYLCCNV) | 0% (10) | AJ319675 | OW (monopartite) | [53] |
| 9 | Tobacco curly shoot virus (TbCSV) | 75% (5) | AJ420318 | OW (monopartite) | [74] |
| 10 | Tomato yellow leaf curl Sardinia virus (TYLCSV-ES) | 40% (2) | Z25751 | OW (monopartite) | [75] |
| 11 | Tomato yellow leaf curl virus Israel strain (TYLCV) | 50% (2) | KY965923 | OW (monopartite) | [45] |
| 12 | Cotton leaf curl Multan virus (CLCuMuV) | 0% (10) | KP762786 | OW (monopartite) | [76] |
| 13 | Papaya leaf curl China virus (PaLCuCNV) | ~60% (10) | AM691554 | OW (monopartite) | [77] |
| 14 | Tomato leaf curl Taiwan virus (ToLCTWV) | 45% (5) | DQ866122 | OW (monopartite) | [78] |
| 17 | Tomato leaf curl New Delhi virus (ToLCNDV)-ES | 100% (25–30) | KF749224 | OW (bipartite) | [79] |
| 18 | Sri Lankan cassava mosaic virus (SLCMV) | 63.3% (10) | MH891840 | OW (bipartite) | [80] |
| 19 | Tomato yellow leaf curl Thailand virus (TYLCTHV) | 50% (5) | GU723742 | OW (bipartite) | [78] |
| 20 | Ramie mosaic virus (RaMoV) | ~100% (2) | KU522485 | OW (bipartite) | [81] |
| Plant Type | Whitefly Type | N a | Percent Egg Survival | Egg-Adult b | N c | Fecundity | |||
|---|---|---|---|---|---|---|---|---|---|
| Tomato | MEAM1 | 45 | 80 ± 3.80 73 ± 10.20 | χ21,88 = 2.26; p = 0.21 | 21 (18–29) | χ21,58 = 4.35; p = 0.23 | 30 | 72 ± 7.20 | F1,58 = 3.59 p = 0.16 |
| MED | 45 | 21 (17–30) | 30 | 82 ± 5.30 | |||||
| Squash | MEAM1 | 45 | 78 ± 8.00 | χ21,88 = 3.31; p = 0.32 | 21 (19–30) | χ21,59 = 3.41; p = 0.52 | 30 | 68 ± 13.50 | F1,58 = 1.09 p = 0.41 |
| MED | 45 | 69 ± 9.70 | 22 (18–28) | 30 | 53 ± 5.30 | ||||
| Prickly sida | MEAM1 | 45 | 56 ± 4.40 | χ21,88 = 7.26; p = 0.08 | 23 (19–32) | χ21,42 = 6.23; p = 0.32 | 30 | 34 ± 9.20 | F1,58 = 4.76 p = 0.09 |
| MED | 45 | 53 ± 10.20 | 24 (19–33) | 30 | 36 ± 2.90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gautam, S.; Mugerwa, H.; Buck, J.W.; Dutta, B.; Coolong, T.; Adkins, S.; Srinivasan, R. Differential Transmission of Old and New World Begomoviruses by Middle East-Asia Minor 1 (MEAM1) and Mediterranean (MED) Cryptic Species of Bemisia tabaci. Viruses 2022, 14, 1104. https://doi.org/10.3390/v14051104
Gautam S, Mugerwa H, Buck JW, Dutta B, Coolong T, Adkins S, Srinivasan R. Differential Transmission of Old and New World Begomoviruses by Middle East-Asia Minor 1 (MEAM1) and Mediterranean (MED) Cryptic Species of Bemisia tabaci. Viruses. 2022; 14(5):1104. https://doi.org/10.3390/v14051104
Chicago/Turabian StyleGautam, Saurabh, Habibu Mugerwa, James W. Buck, Bhabesh Dutta, Tim Coolong, Scott Adkins, and Rajagopalbabu Srinivasan. 2022. "Differential Transmission of Old and New World Begomoviruses by Middle East-Asia Minor 1 (MEAM1) and Mediterranean (MED) Cryptic Species of Bemisia tabaci" Viruses 14, no. 5: 1104. https://doi.org/10.3390/v14051104
APA StyleGautam, S., Mugerwa, H., Buck, J. W., Dutta, B., Coolong, T., Adkins, S., & Srinivasan, R. (2022). Differential Transmission of Old and New World Begomoviruses by Middle East-Asia Minor 1 (MEAM1) and Mediterranean (MED) Cryptic Species of Bemisia tabaci. Viruses, 14(5), 1104. https://doi.org/10.3390/v14051104