
Advances in Bovine Coronavirus Epidemiology



Abstract
:1. Introduction
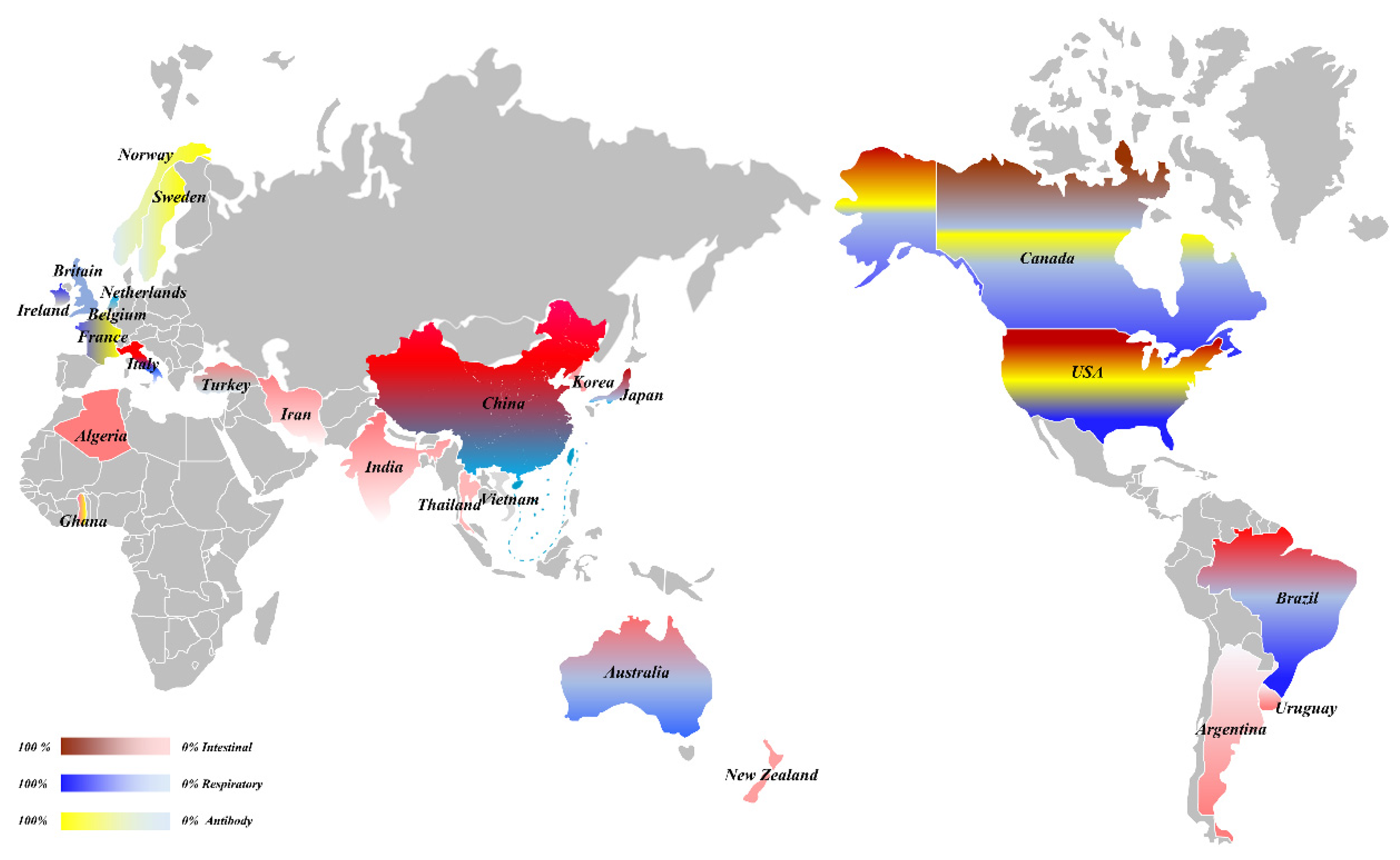
2. The Prevalence of BCoV
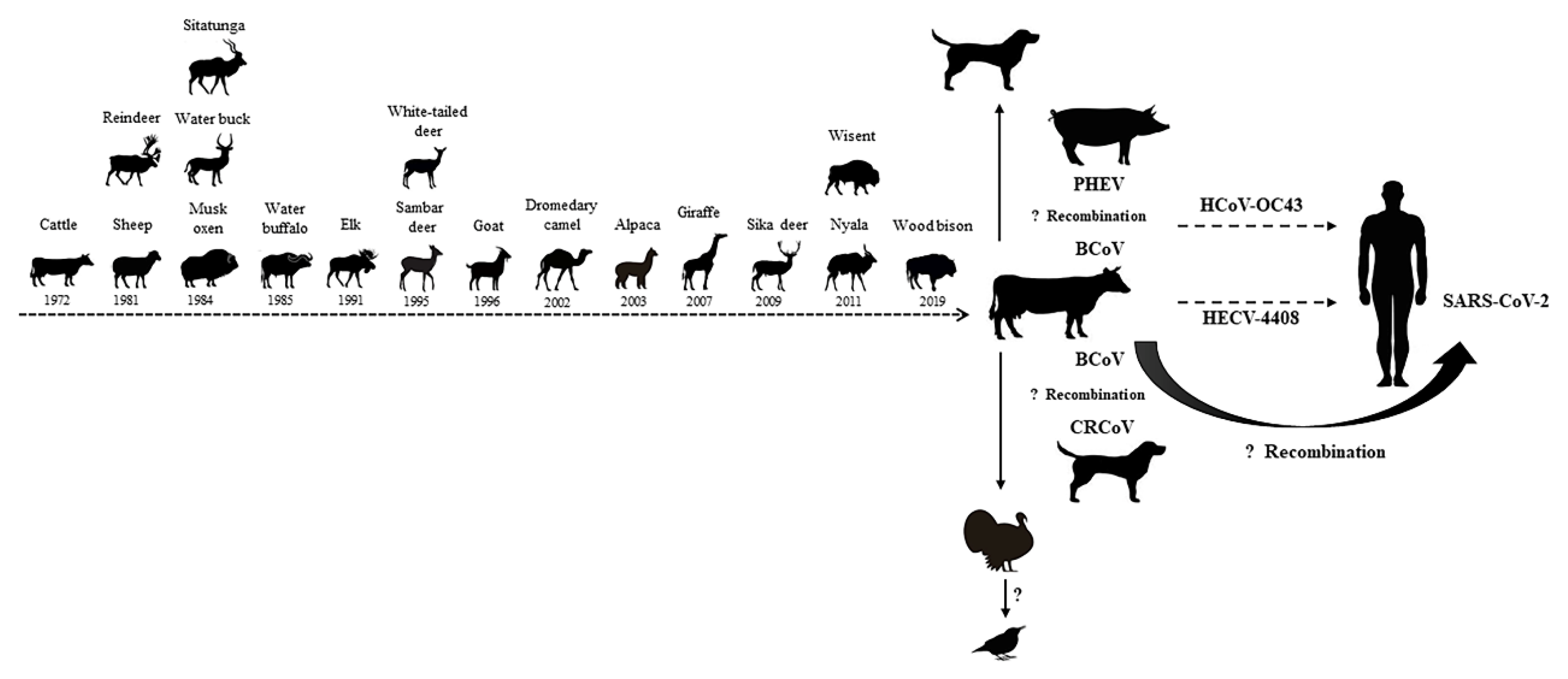
2.1. The Origin of BCoV
2.2. The Transmission of BCoV
2.2.1. The Transmission of Enteric BCoV
2.2.2. The Transmission of Respiratory BCoV
2.2.3. The Serological Surveys of BCoV
2.3. The BCoV Infection-Associated Factors
2.4. Co-infection of BCoV with Typical Bovine Enteric and Respiratory Pathogens
2.4.1. Co-infection of BCoV with Typical Bovine Enteric Pathogens
2.4.2. Co-infection of BCoV with Typical Bovine Respiratory Pathogens
2.5. Pathogenicity of BCoV
3. Genetic Evolution of BCoV
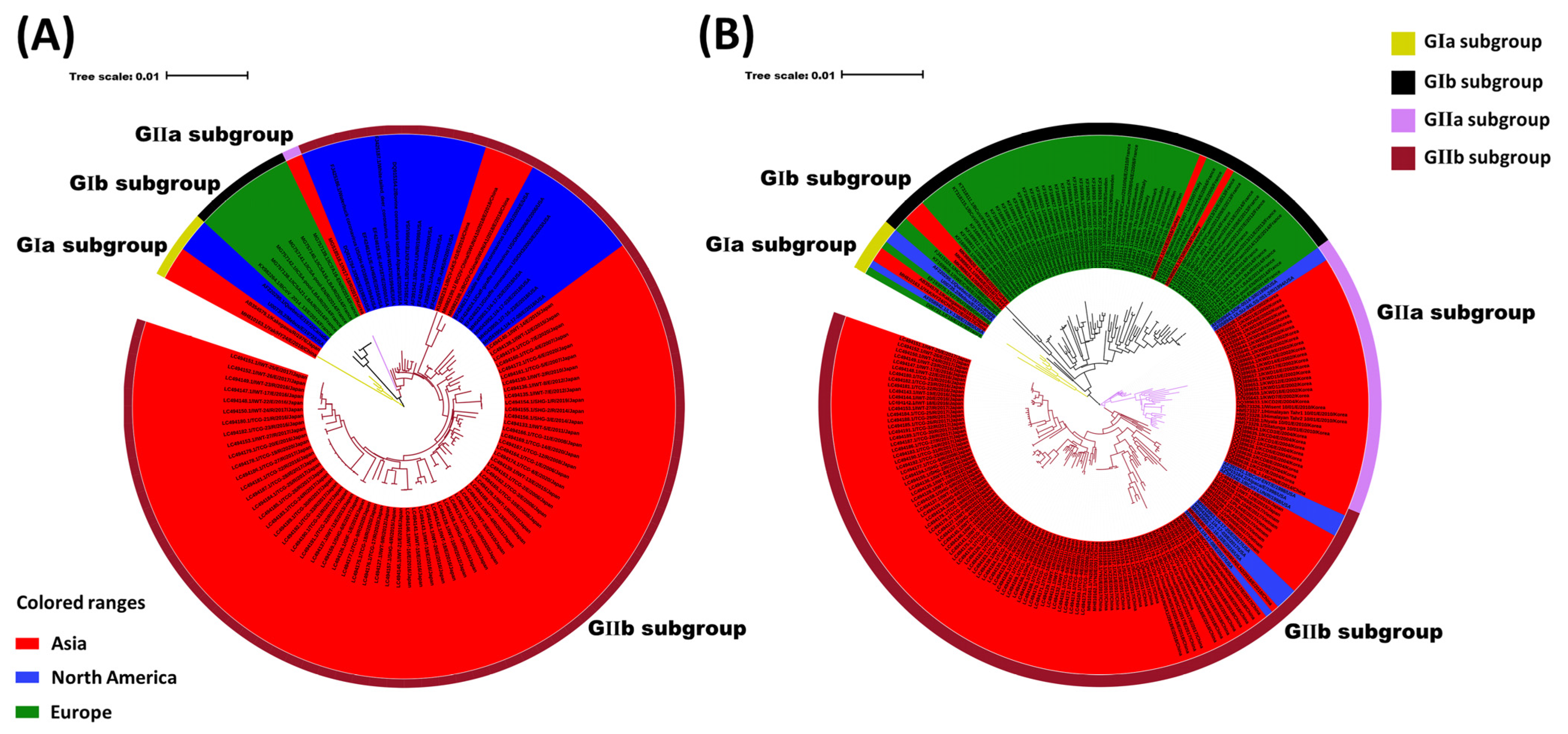
3.1. Phylogenetic Analyses of BCoV Strains
3.2. Comparison Analysis of BCoV Strains between the Enteric and Respiratory Tracts
4. Potential Cross-Species Transmission of BCoV
5. Conclusions and Future Perspective
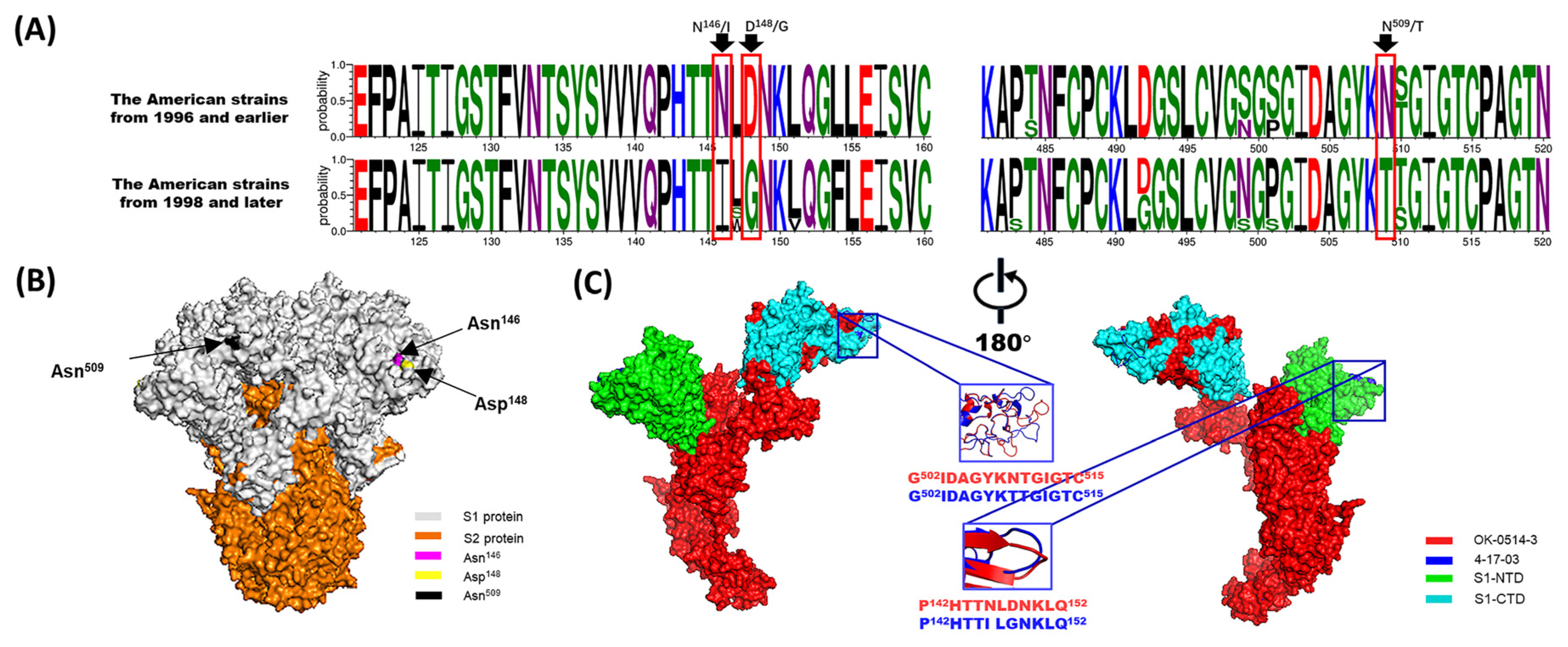
5.1. Potential Two-Receptor Binding Motif System for BCoV Entry
5.2. BCoV and SARS-CoV-2
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saif, L.J.; Jung, K. Comparative Pathogenesis of Bovine and Porcine Respiratory Coronaviruses in the Animal Host Species and SARS-CoV-2 in Humans. J. Clin. Microbiol. 2020, 58, e01355-20. [Google Scholar] [CrossRef] [PubMed]
- Saif, L.J. Bovine respiratory coronavirus. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Herbst, W.; Kousoulas, K.G.; Storz, J. Biological and genetic characterization of a hemagglutinating coronavirus isolated from a diarrhoeic child. J. Med. Virol. 1994, 44, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Amer, H.M. Bovine-like coronaviruses in domestic and wild ruminants. Anim. Health Res. Rev. 2018, 19, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Mebus, C.A.; Stair, E.L.; Rhodes, M.B.; Twiehaus, M.J. Pathology of neonatal calf diarrhea induced by a coronavirus-like agent. Vet. Pathol. 1973, 10, 45–64. [Google Scholar] [CrossRef] [Green Version]
- McNulty, M.S.; Bryson, D.G.; Allan, G.M.; Logan, E.F. Coronavirus infection of the bovine respiratory tract. Vet. Microbiol. 1984, 9, 425–434. [Google Scholar] [CrossRef]
- Bartels, C.J.M.; Holzhauer, M.; Jorritsma, R.; Swart, W.A.J.M.; Lam, T.J.G.M. Prevalence, prediction and risk factors of enteropathogens in normal and non-normal faeces of young Dutch dairy calves. Prev. Vet. Med. 2010, 93, 162–169. [Google Scholar] [CrossRef]
- Burimuah, V.; Sylverken, A.; Owusu, M.; El-Duah, P.; Yeboah, R.; Lamptey, J.; Frimpong, Y.O.; Agbenyega, O.; Folitse, R.; Emikpe, B.; et al. Molecular-based cross-species evaluation of bovine coronavirus infection in cattle, sheep and goats in Ghana. BMC Vet. Res. 2020, 16, 405. [Google Scholar] [CrossRef]
- Gagea, M.I.; Bateman, K.G.; van Dreumel, T.; McEwen, B.J.; Carman, S.; Archambault, M.; Shanahan, R.A.; Caswell, J.L. Diseases and pathogens associated with mortality in Ontario beef feedlots. J. Vet. Diagn. Investig. 2006, 18, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Kanno, T.; Hatama, S.; Ishihara, R.; Uchida, I. Molecular analysis of the S glycoprotein gene of bovine coronaviruses isolated in Japan from 1999 to 2006. J. Gen. Virol. 2007, 88 Pt 4, 1218–1224. [Google Scholar] [CrossRef]
- Keha, A.; Xue, L.; Yan, S.; Yue, H.; Tang, C. Prevalence of a novel bovine coronavirus strain with a recombinant hemagglutinin/esterase gene in dairy calves in China. Transbound Emerg. Dis. 2019, 66, 1971–1981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Li, J.; Deng, Z.; Xiong, W.; Wang, Q.; Hu, Y.Q. Comprehensive detection and identification of seven animal coronaviruses and human respiratory coronavirus 229E with a microarray hybridization assay. Intervirology 2010, 53, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Campolo, M.; Mari, V.; Desario, C.; Colaianni, M.L.; Di Trani, L.; Cordioli, P.; Buonavoglia, C. A candidate modified-live bovine coronavirus vaccine: Safety and immunogenicity evaluation. New Microbiol. 2009, 32, 109–113. [Google Scholar]
- Kin, N.; Miszczak, F.; Diancourt, L.; Caro, V.; Moutou, F.; Vabret, A.; Ar Gouilh, M. Comparative molecular epidemiology of two closely related coronaviruses, bovine coronavirus (BCoV) and human coronavirus OC43 (HCoV-OC43), reveals a different evolutionary pattern. Infect. Genet. Evol. 2016, 40, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Hagglund, S.; Hakhverdyan, M.; Alenius, S.; Larsen, L.E.; Belak, S. Molecular epidemiology of bovine coronavirus on the basis of comparative analyses of the S gene. J. Clin. Microbiol. 2006, 44, 957–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohlson, A.; Blanco-Penedo, I.; Fall, N. Comparison of Bovine coronavirus-specific and Bovine respiratory syncytial virus-specific antibodies in serum versus milk samples detected by enzyme-linked immunosorbent assay. J. Vet. Diagn. Investig. 2014, 26, 113–116. [Google Scholar] [CrossRef] [Green Version]
- Plummer, P.J.; Rohrbach, B.W.; Daugherty, R.A.; Daugherty, R.A.; Thomas, K.V.; Wilkes, R.P.; Duggan, F.E.; Kennedy, M.A. Effect of intranasal vaccination against bovine enteric coronavirus on the occurrence of respiratory tract disease in a commercial backgrounding feedlot. J. Am. Vet. Med. Assoc. 2004, 225, 726–731. [Google Scholar] [CrossRef]
- Takiuchi, E.; Stipp, D.T.; Alfieri, A.F.; Alfieri, A.A. Improved detection of bovine coronavirus N gene in faeces of calves infected naturally by a semi-nested PCR assay and an internal control. J. Virol. Methods 2006, 131, 148–154. [Google Scholar] [CrossRef]
- Duckmanton, L.; Carman, S.; Nagy, E.; Petric, M. Detection of bovine torovirus in fecal specimens of calves with diarrhea from Ontario farms. J. Clin. Microbiol. 1998, 36, 1266–1270. [Google Scholar] [CrossRef] [Green Version]
- Bok, M.; Mino, S.; Rodriguez, D.; Badaracco, A.; Nunes, I.; Souza, S.P.; Bilbao, G.; Louge Uriarte, E.; Galarza, R.; Vega, C.; et al. Molecular and antigenic characterization of bovine Coronavirus circulating in Argentinean cattle during 1994–2010. Vet. Microbiol. 2015, 181, 221–229. [Google Scholar] [CrossRef]
- Heckert, R.A.; Saif, L.J.; Hoblet, K.H.; Agnes, A.G. A longitudinal study of bovine coronavirus enteric and respiratory infections in dairy calves in two herds in Ohio. Vet. Microbiol. 1990, 22, 187–201. [Google Scholar] [CrossRef]
- Hasoksuz, M.; Hoet, A.E.; Loerch, S.C.; Wittum, T.E.; Nielsen, P.R.; Saif, L.J. Detection of respiratory and enteric shedding of bovine coronaviruses in cattle in an Ohio feedlot. J. Vet. Diagn. Investig. 2002, 14, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Hill, J.E.; Fernando, C.; Alexander, T.W.; Timsit, E.; van der Meer, F.; Huang, Y. Respiratory viruses identified in western Canadian beef cattle by metagenomic sequencing and their association with bovine respiratory disease. Transbound Emerg. Dis. 2019, 66, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.J.; Morgan, J.H.; Chanter, N.; Jones, P.W.; Bridger, J.C.; Debney, T.G.; Bunch, K.J. Microbiology of calf diarrhoea in southern Britain. Vet. Rec. 1986, 119, 34–39. [Google Scholar] [CrossRef] [PubMed]
- de Graaf, D.C.; Vanopdenbosch, E.; Ortega-Mora, L.M.; Abbassi, H.; Peeters, J.E. A review of the importance of cryptosporidiosis in farm animals. Int. J. Parasitol. 1999, 29, 1269–1287. [Google Scholar] [CrossRef]
- Mawatari, T.; Hirano, K.; Ikeda, H.; Tsunemitsu, H.; Suzuki, T. Surveillance of diarrhea-causing pathogens in dairy and beef cows in Yamagata Prefecture, Japan from 2002 to 2011. Microbiol. Immunol. 2014, 58, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.H.; Kim, G.Y.; Yoon, S.S.; Park, S.J.; Kim, Y.J.; Sung, C.M.; Shin, S.S.; Lee, B.J.; Kang, M.I.; Park, N.Y.; et al. Molecular analysis of S gene of spike glycoprotein of winter dysentery bovine coronavirus circulated in Korea during 2002–2003. Virus Res. 2005, 108, 207–212. [Google Scholar] [CrossRef]
- Alkan, F.; Ozkul, A.; Bilge-Dagalp, S.; Karaoglu, T.; Oguzoglu, T.C.; Caliskan, E.; Burgu, I. The detection and genetic characterization based on the S1 gene region of BCoVs from respiratory and enteric infections in Turkey. Transbound Emerg. Dis. 2011, 58, 179–185. [Google Scholar] [CrossRef]
- Hasoksuz, M.; Kayar, A.; Dodurka, T.; Ilgaz, A. Detection of respiratory and enteric shedding of bovine coronaviruses in cattle in Northwestern Turkey. Acta Vet. Hung. 2005, 53, 137–146. [Google Scholar] [CrossRef]
- Jeong, J.H.; Kim, G.Y.; Yoon, S.S.; Park, S.J.; Kim, Y.J.; Sung, C.M.; Jang, O.J.; Shin, S.S.; Koh, H.B.; Lee, B.J.; et al. Detection and isolation of winter dysentery bovine coronavirus circulated in Korea during 2002–2004. J. Vet. Med. Sci. 2005, 67, 187–189. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Kim, H.Y.; Choi, E.W.; Kim, D. Causative agents and epidemiology of diarrhea in Korean native calves. J. Vet. Sci. 2019, 20, e64. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Lim, G.K.; Park, S.I.; Kim, H.H.; Koh, H.B.; Cho, K.O. Detection and molecular characterization of calf diarrhoea bovine coronaviruses circulating in South Korea during 2004–2005. Zoonoses Public Health 2007, 54, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Mari, V.; Desario, C.; Campolo, M.; Elia, G.; Martella, V.; Greco, G.; Cirone, F.; Colaianni, M.L.; Cordioli, P.; et al. Severe outbreak of bovine coronavirus infection in dairy cattle during the warmer season. Vet. Microbiol. 2008, 126, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Brandao, P.E.; Gregori, F.; Richtzenhain, L.J.; Rosales, C.A.; Villarreal, L.Y.; Jerez, J.A. Molecular analysis of Brazilian strains of bovine coronavirus (BCoV) reveals a deletion within the hypervariable region of the S1 subunit of the spike glycoprotein also found in human coronavirus OC43. Arch. Virol. 2006, 151, 1735–1748. [Google Scholar] [CrossRef] [Green Version]
- Beuttemmuller, E.A.; Alfieri, A.F.; Headley, S.A.; Alfieri, A.A. Brazilian strain of bovine respiratory coronavirus is derived from dual enteric and respiratory tropism. Genet. Mol. Res. 2017, 16, 28387879. [Google Scholar] [CrossRef]
- Coura, F.M.; Freitas, M.D.; Ribeiro, J.; de Leme, R.A.; de Souza, C.; Alfieri, A.A.; Facury Filho, E.J.; de Carvalho, A.U.; Silva, M.X.; Lage, A.P.; et al. Longitudinal study of Salmonella spp., diarrheagenic Escherichia coli, Rotavirus, and Coronavirus isolated from healthy and diarrheic calves in a Brazilian dairy herd. Trop. Anim. Health Prod. 2015, 47, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Headley, S.A.; Okano, W.; Balbo, L.C.; Marcasso, R.A.; Oliveira, T.E.; Alfieri, A.F.; Negri Filho, L.C.; Michelazzo, M.Z.; Rodrigues, S.C.; Baptista, A.L.; et al. Molecular survey of infectious agents associated with bovine respiratory disease in a beef cattle feedlot in southern Brazil. J. Vet. Diagn. Investig. 2018, 30, 249–251. [Google Scholar] [CrossRef] [Green Version]
- Al Mawly, J.; Grinberg, A.; Prattley, D.; Moffat, J.; French, N. Prevalence of endemic enteropathogens of calves in New Zealand dairy farms. N. Z. Vet. J. 2015, 63, 147–152. [Google Scholar] [CrossRef]
- Izzo, M.M.; Kirkland, P.D.; Mohler, V.L.; Perkins, N.R.; Gunn, A.A.; House, J.K. Prevalence of major enteric pathogens in Australian dairy calves with diarrhoea. Aust. Vet. J. 2011, 89, 167–173. [Google Scholar] [CrossRef]
- Ammar, S.S.; Mokhtaria, K.; Tahar, B.B.; Amar, A.A.; Redha, B.A.; Yuva, B.; Mohamed, H.S.; Abdellatif, N.; Laid, B. Prevalence of rotavirus (GARV) and coronavirus (BCoV) associated with neonatal diarrhea in calves in western Algeria. Asian Pac. J. Trop. Biomed. 2014, 4 (Suppl. 1), S318–S322. [Google Scholar] [CrossRef] [Green Version]
- Lotfollahzadeh, S.; Madadgar, O.; Reza Mohebbi, M.; Reza Mokhber Dezfouli, M.; George Watson, D. Bovine coronavirus in neonatal calf diarrhoea in Iran. Vet. Med. Sci. 2020, 6, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Singasa, K.; Songserm, T.; Lertwatcharasarakul, P.; Arunvipas, P. Molecular and phylogenetic characterization of bovine coronavirus virus isolated from dairy cattle in Central Region, Thailand. Trop. Anim. Health Prod. 2017, 49, 1523–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brar, A.P.S.; Sood, N.K.; Kaur, P.; Singla, L.D.; Sandhu, B.S.; Gupta, K.; Narang, D.; Singh, C.K.; Chandra, M. Periurban outbreaks of bovine calf scours in Northern India caused by Cryptosporidium in association with other enteropathogens. Epidemiol. Infect. 2017, 145, 2717–2726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.; Tark, D.; Le, V.P.; Choe, S.; Cha, R.M.; Park, G.N.; Cho, I.S.; Nga, B.T.T.; Lan, N.T.; An, D.J. Genetic characterization of bovine coronavirus in Vietnam. Virus Genes 2019, 55, 415–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.; Su, M.; Li, Z.; Wang, X.; Qi, S.; Zhao, F.; Li, L.; Guo, D.; Feng, L.; Li, B.; et al. Epidemiological survey and genetic diversity of bovine coronavirus in Northeast China. Virus Res. 2021, 308, 198632. [Google Scholar] [CrossRef] [PubMed]
- Bertoni, E.; Aduriz, M.; Bok, M.; Vega, C.; Saif, L.; Aguirre, D.; Cimino, R.O.; Mino, S.; Parreno, V. First report of group A rotavirus and bovine coronavirus associated with neonatal calf diarrhea in the northwest of Argentina. Trop. Anim. Health Prod. 2020, 52, 2761–2768. [Google Scholar] [CrossRef]
- Castells, M.; Giannitti, F.; Caffarena, R.D.; Casaux, M.L.; Schild, C.; Castells, D.; Riet-Correa, F.; Victoria, M.; Parreno, V.; Colina, R. Bovine coronavirus in Uruguay: Genetic diversity, risk factors and transboundary introductions from neighboring countries. Arch. Virol. 2019, 164, 2715–2724. [Google Scholar] [CrossRef] [Green Version]
- Fulton, R.W.; Blood, K.S.; Panciera, R.J.; Payton, M.E.; Ridpath, J.F.; Confer, A.W.; Saliki, J.T.; Burge, L.T.; Welsh, R.D.; Johnson, B.J.; et al. Lung pathology and infectious agents in fatal feedlot pneumonias and relationship with mortality, disease onset, and treatments. J. Vet. Diagn. Investig. 2009, 21, 464–477. [Google Scholar] [CrossRef] [Green Version]
- Francoz, D.; Buczinski, S.; Belanger, A.M.; Forte, G.; Labrecque, O.; Tremblay, D.; Wellemans, V.; Dubuc, J. Respiratory pathogens in Quebec dairy calves and their relationship with clinical status, lung consolidation, and average daily gain. J. Vet. Intern. Med. 2015, 29, 381–387. [Google Scholar] [CrossRef]
- Decaro, N.; Campolo, M.; Desario, C.; Cirone, F.; D’Abramo, M.; Lorusso, E.; Greco, G.; Mari, V.; Colaianni, M.L.; Elia, G.; et al. Respiratory disease associated with bovine coronavirus infection in cattle herds in Southern Italy. J. Vet. Diagn. Investig. 2008, 20, 28–32. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, R.; Mooney, J.; Connaghan, E.; Furphy, C.; Graham, D.A. Patterns of detection of respiratory viruses in nasal swabs from calves in Ireland: A retrospective study. Vet. Rec. 2014, 175, 351. [Google Scholar] [CrossRef] [PubMed]
- Salem, E.; Dhanasekaran, V.; Cassard, H.; Hause, B.; Maman, S.; Meyer, G.; Ducatez, M.F. Global Transmission, Spatial Segregation, and Recombination Determine the Long-Term Evolution and Epidemiology of Bovine Coronaviruses. Viruses 2020, 12, 534. [Google Scholar] [CrossRef] [PubMed]
- Pratelli, A.; Cirone, F.; Capozza, P.; Trotta, A.; Corrente, M.; Balestrieri, A.; Buonavoglia, C. Bovine respiratory disease in beef calves supported long transport stress: An epidemiological study and strategies for control and prevention. Res. Vet. Sci. 2021, 135, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Mekata, H.; Hamabe, S.; Sudaryatma, P.E.; Kobayashi, I.; Kanno, T.; Okabayashi, T. Molecular epidemiological survey and phylogenetic analysis of bovine respiratory coronavirus in Japan from 2016 to 2018. J. Vet. Med. Sci. 2020, 82, 726–730. [Google Scholar] [CrossRef] [Green Version]
- Hick, P.M.; Read, A.J.; Lugton, I.; Busfield, F.; Dawood, K.E.; Gabor, L.; Hornitzky, M.; Kirkland, P.D. Coronavirus infection in intensively managed cattle with respiratory disease. Aust. Vet. J. 2012, 90, 381–386. [Google Scholar] [CrossRef]
- Elazhary, M.A.; Frechette, J.L.; Silim, A.; Roy, R.S. Serological evidence of some bovine viruses in the caribou (Rangifer tarandus caribou) in Quebec. J. Wildl. Dis. 1981, 17, 609–612. [Google Scholar] [CrossRef]
- Harms, N.J.; Jung, T.S.; Andrew, C.L.; Surujballi, O.P.; VanderKop, M.; Savic, M.; Powell, T. Health Status of Reintroduced Wood Bison (Bison Bison Athabascae): Assessing the Conservation Value of an Isolated Population in Northwestern Canada. J. Wildl. Dis. 2019, 55, 44–53. [Google Scholar] [CrossRef]
- Beaudeau, F.; Bjorkman, C.; Alenius, S.; Frossling, J. Spatial patterns of bovine corona virus and bovine respiratory syncytial virus in the Swedish beef cattle population. Acta Vet. Scand. 2010, 52, 33. [Google Scholar] [CrossRef] [Green Version]
- Gulliksen, S.M.; Jor, E.; Lie, K.I.; Hamnes, I.S.; Loken, T.; Akerstedt, J.; Osteras, O. Enteropathogens and risk factors for diarrhea in Norwegian dairy calves. J. Dairy Sci. 2009, 92, 5057–5066. [Google Scholar] [CrossRef]
- Klem, T.B.; Rimstad, E.; Stokstad, M. Occurrence and phylogenetic analysis of bovine respiratory syncytial virus in outbreaks of respiratory disease in Norway. BMC Vet. Res. 2014, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Toftaker, I.; Agren, E.; Stokstad, M.; Nodtvedt, A.; Frossling, J. Herd level estimation of probability of disease freedom applied on the Norwegian control program for bovine respiratory syncytial virus and bovine coronavirus. Prev. Vet. Med. 2020, 181, 104494. [Google Scholar] [CrossRef] [PubMed]
- Bendali, F.; Bichet, H.; Schelcher, F.; Sanaa, M. Pattern of diarrhoea in newborn beef calves in south-west France. Vet. Res. 1999, 30, 61–74. [Google Scholar] [PubMed]
- Pardon, B.; De Bleecker, K.; Dewulf, J.; Callens, J.; Boyen, F.; Catry, B.; Deprez, P. Prevalence of respiratory pathogens in diseased, non-vaccinated, routinely medicated veal calves. Vet. Rec. 2011, 169, 278. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Elia, G.; Campolo, M.; Desario, C.; Mari, V.; Radogna, A.; Colaianni, M.L.; Cirone, F.; Tempesta, M.; Buonavoglia, C. Detection of bovine coronavirus using a TaqMan-based real-time RT-PCR assay. J. Virol. Methods 2008, 151, 167–171. [Google Scholar] [CrossRef]
- Gomez, D.E.; Arroyo, L.G.; Poljak, Z.; Viel, L.; Weese, J.S. Detection of Bovine Coronavirus in Healthy and Diarrheic Dairy Calves. J. Vet. Intern. Med. 2017, 31, 1884–1891. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Kim, G.Y.; Choy, H.E.; Hong, Y.J.; Saif, L.J.; Jeong, J.H.; Park, S.I.; Kim, H.H.; Kim, S.K.; Shin, S.S.; et al. Dual enteric and respiratory tropisms of winter dysentery bovine coronavirus in calves. Arch. Virol. 2007, 152, 1885–1900. [Google Scholar] [CrossRef] [Green Version]
- Fulton, R.W.; Step, D.L.; Wahrmund, J.; Burge, L.J.; Payton, M.E.; Cook, B.J.; Burken, D.; Richards, C.J.; Confer, A.W. Bovine coronavirus (BCV) infections in transported commingled beef cattle and sole-source ranch calves. Can. J. Vet. Res. 2011, 75, 191–199. [Google Scholar]
- Workman, A.M.; Kuehn, L.A.; McDaneld, T.G.; Clawson, M.L.; Loy, J.D. Longitudinal study of humoral immunity to bovine coronavirus, virus shedding, and treatment for bovine respiratory disease in pre-weaned beef calves. BMC Vet. Res. 2019, 15, 161. [Google Scholar] [CrossRef]
- Lin, X.; O’Reilly, K.L.; Burrell, M.L.; Storz, J. Infectivity-neutralizing and hemagglutinin-inhibiting antibody responses to respiratory coronavirus infections of cattle in pathogenesis of shipping fever pneumonia. Clin. Diagn. Lab. Immunol. 2001, 8, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Storz, J.; Lin, X.; Purdy, C.W.; Chouljenko, V.N.; Kousoulas, K.G.; Enright, F.M.; Gilmore, W.C.; Briggs, R.E.; Loan, R.W. Coronavirus and Pasteurella infections in bovine shipping fever pneumonia and Evans’ criteria for causation. J. Clin. Microbiol. 2000, 38, 3291–3298. [Google Scholar] [CrossRef]
- Uhde, F.L.; Kaufmann, T.; Sager, H.; Albini, S.; Zanoni, R.; Schelling, E.; Meylan, M. Prevalence of four enteropathogens in the faeces of young diarrhoeic dairy calves in Switzerland. Vet. Rec. 2008, 163, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Mitra, N.; Cernicchiaro, N.; Torres, S.; Li, F.; Hause, B.M. Metagenomic characterization of the virome associated with bovine respiratory disease in feedlot cattle identified novel viruses and suggests an etiologic role for influenza D virus. J. Gen. Virol. 2016, 97, 1771–1784. [Google Scholar] [CrossRef] [PubMed]
- Ridpath, J.F.; Fulton, R.W.; Bauermann, F.V.; Falkenberg, S.M.; Welch, J.; Confer, A.W. Sequential exposure to bovine viral diarrhea virus and bovine coronavirus results in increased respiratory disease lesions: Clinical, immunologic, pathologic, and immunohistochemical findings. J. Vet. Diagn. Investig. 2020, 32, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Beaudeau, F.; Ohlson, A.; Emanuelson, U. Associations between bovine coronavirus and bovine respiratory syncytial virus infections and animal performance in Swedish dairy herds. J. Dairy Sci. 2010, 93, 1523–1533. [Google Scholar] [CrossRef] [Green Version]
- Wolff, C.; Emanuelson, U.; Ohlson, A.; Alenius, S.; Fall, N. Bovine respiratory syncytial virus and bovine coronavirus in Swedish organic and conventional dairy herds. Acta Vet. Scand. 2015, 57, 2. [Google Scholar] [CrossRef] [Green Version]
- Storz, J.; Purdy, C.W.; Lin, X.; Burrell, M.; Truax, R.E.; Briggs, R.E.; Frank, G.H.; Loan, R.W. Isolation of respiratory bovine coronavirus, other cytocidal viruses, and Pasteurella spp. from cattle involved in two natural outbreaks of shipping fever. J. Am. Vet. Med. Assoc. 2000, 216, 1599–1604. [Google Scholar] [CrossRef]
- Fahkrajang, W.; Sudaryatma, P.E.; Mekata, H.; Hamabe, S.; Saito, A.; Okabayashi, T. Bovine respiratory coronavirus enhances bacterial adherence by upregulating expression of cellular receptors on bovine respiratory epithelial cells. Vet. Microbiol. 2021, 255, 109017. [Google Scholar] [CrossRef]
- Kapil, S.; Pomeroy, K.A.; Goyal, S.M.; Trent, A.M. Experimental infection with a virulent pneumoenteric isolate of bovine coronavirus. J. Vet. Diagn. Investig. 1991, 3, 88–89. [Google Scholar] [CrossRef] [Green Version]
- Ismail, M.M.; Cho, K.O.; Ward, L.A.; Saif, L.J.; Saif, Y.M. Experimental bovine coronavirus in turkey poults and young chickens. Avian Dis. 2001, 45, 157–163. [Google Scholar] [CrossRef]
- Kaneshima, T.; Hohdatsu, T.; Hagino, R.; Hosoya, S.; Nojiri, Y.; Murata, M.; Takano, T.; Tanabe, M.; Tsunemitsu, H.; Koyama, H. The infectivity and pathogenicity of a group 2 bovine coronavirus in pups. J. Vet. Med. Sci. 2007, 69, 301–303. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.R. HLA class I antigen serves as a receptor for human coronavirus OC43. Immunol. Investig. 1993, 22, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Zeng, L.P.; Yang, X.L.; Ge, X.Y.; Zhang, W.; Li, B.; Xie, J.Z.; Shen, X.R.; Zhang, Y.Z.; Wang, N.; et al. Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus. PLoS Pathog. 2017, 13, e1006698. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.; Lee, P.; Tsang, A.K.; Yip, C.C.; Tse, H.; Lee, R.A.; So, L.Y.; Lau, Y.L.; Chan, K.H.; Woo, P.C.; et al. Molecular epidemiology of human coronavirus OC43 reveals evolution of different genotypes over time and recent emergence of a novel genotype due to natural recombination. J. Virol. 2011, 85, 11325–11337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzo, G.; Drigo, M.; Legnardi, M.; Grassi, L.; Pasotto, D.; Menandro, M.L.; Cecchinato, M.; Tucciarone, C.M. Bovine Coronavirus: Variability, Evolution, and Dispersal Patterns of a No Longer Neglected Betacoronavirus. Viruses 2020, 12, 1285. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.; Xu, L.; Lin, Y.L.; Chen, L.; Pasquarella, J.R.; Holmes, K.V.; Li, F. Crystal structure of bovine coronavirus spike protein lectin domain. J. Biol. Chem. 2012, 287, 41931–41938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasoksuz, M.; Sreevatsan, S.; Cho, K.O.; Hoet, A.E.; Saif, L.J. Molecular analysis of the S1 subunit of the spike glycoprotein of respiratory and enteric bovine coronavirus isolates. Virus Res. 2002, 84, 101–109. [Google Scholar] [CrossRef]
- Munir, K.; Ashraf, S.; Munir, I.; Khalid, H.; Muneer, M.A.; Mukhtar, N.; Amin, S.; Ashraf, S.; Imran, M.A.; Chaudhry, U.; et al. Zoonotic and reverse zoonotic events of SARS-CoV-2 and their impact on global health. Emerg. Microbes Infect. 2020, 9, 2222–2235. [Google Scholar] [CrossRef]
- Vijgen, L.; Keyaerts, E.; Moes, E.; Thoelen, I.; Wollants, E.; Lemey, P.; Vandamme, A.M.; Van Ranst, M. Complete genomic sequence of human coronavirus OC43: Molecular clock analysis suggests a relatively recent zoonotic coronavirus transmission event. J. Virol. 2005, 79, 1595–1604. [Google Scholar] [CrossRef] [Green Version]
- Patrono, L.V.; Samuni, L.; Corman, V.M.; Nourifar, L.; Rothemeier, C.; Wittig, R.M.; Drosten, C.; Calvignac-Spencer, S.; Leendertz, F.H. Human coronavirus OC43 outbreak in wild chimpanzees, Côte d´Ivoire, 2016. Emerg. Microbes Infect. 2018, 7, 118. [Google Scholar] [CrossRef] [Green Version]
- Bidokhti, M.R.M.; Traven, M.; Krishna, N.K.; Munir, M.; Belak, S.; Alenius, S.; Cortey, M. Evolutionary dynamics of bovine coronaviruses: Natural selection pattern of the spike gene implies adaptive evolution of the strains. J. Gen. Virol. 2013, 94 Pt 9, 2036–2049. [Google Scholar] [CrossRef]
- Lu, S.; Wang, Y.; Chen, Y.; Wu, B.; Qin, K.; Zhao, J.; Lou, Y.; Tan, W. Discovery of a novel canine respiratory coronavirus support genetic recombination among betacoronavirus1. Virus Res. 2017, 237, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Hulswit, R.J.; de Haan, C.A.; Bosch, B.J. Coronavirus Spike Protein and Tropism Changes. Adv. Virus Res. 2016, 96, 29–57. [Google Scholar] [PubMed]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar] [PubMed] [Green Version]
- Guan, H.; Wang, Y.; Perculija, V.; Saeed, A.; Liu, Y.; Li, J.; Jan, S.S.; Li, Y.; Zhu, P.; Ouyang, S. Cryo-electron Microscopy Structure of the Swine Acute Diarrhea Syndrome Coronavirus Spike Glycoprotein Provides Insights into Evolution of Unique Coronavirus Spike Proteins. J. Virol. 2020, 94, e01301-20. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Qiao, S.; Guo, R.; Wang, X. Cryo-EM structures of HKU2 and SADS-CoV spike glycoproteins provide insights into coronavirus evolution. Nat. Commun. 2020, 11, 3070. [Google Scholar] [CrossRef]
- Szczepanski, A.; Owczarek, K.; Bzowska, M.; Gula, K.; Drebot, I.; Ochman, M.; Maksym, B.; Rajfur, Z.; Mitchell, J.A.; Pyrc, K. Canine Respiratory Coronavirus, Bovine Coronavirus, and Human Coronavirus OC43: Receptors and Attachment Factors. Viruses 2019, 11, 328. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.M.; Lau, S.K.; Woo, P.C.; Tse, H.; Zheng, B.J.; Chen, L.; Huang, J.D.; Yuen, K.Y. Identification of major histocompatibility complex class I C molecule as an attachment factor that facilitates coronavirus HKU1 spike-mediated infection. J. Virol. 2009, 83, 1026–1035. [Google Scholar] [CrossRef] [Green Version]
- Qian, Z.; Ou, X.; Goes, L.G.; Osborne, C.; Castano, A.; Holmes, K.V.; Dominguez, S.R. Identification of the Receptor-Binding Domain of the Spike Glycoprotein of Human Betacoronavirus HKU1. J. Virol. 2015, 89, 8816–8827. [Google Scholar] [CrossRef] [Green Version]
- Millet, J.K.; Jaimes, J.A.; Whittaker, G.R. Molecular diversity of coronavirus host cell entry receptors. FEMS Microbiol. Rev. 2021, 45, fuaa057. [Google Scholar] [CrossRef]
- Aboubakr, H.A.; Sharafeldin, T.A.; Goyal, S.M. Stability of SARS-CoV-2 and other coronaviruses in the environment and on common touch surfaces and the influence of climatic conditions: A review. Transbound Emerg. Dis. 2021, 68, 296–312. [Google Scholar] [CrossRef]
- Hein, W.R.; Griebel, P.J. A road less travelled: Large animal models in immunological research. Nat. Rev. Immunol. 2003, 3, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Guzman, E.; Montoya, M. Contributions of Farm Animals to Immunology. Front. Vet. Sci. 2018, 5, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenas, A.; Borge, C.; Carbonero, A.; Garcia-Bocanegra, I.; Cano-Terriza, D.; Caballero, J.; Arenas-Montes, A. Bovine Coronavirus Immune Milk Against COVID-19. Front. Immunol. 2021, 12, 637152. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, L.; Wernike, K.; Hoffmann, D.; Mettenleiter, T.C.; Beer, M. Experimental Infection of Cattle with SARS-CoV-2. Emerg. Infect. Dis. 2020, 26, 2979–2981. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Source | Time Sequence | Continent | Country | Positive Rate | References |
|---|---|---|---|---|---|
| Intestinal tract | Before 2000 | America | USA | 16.4–84% | [21,22] |
| Canada | 6.5–70% | [9,19,23] | |||
| Argentina | 2.41–10.52% | [20,46] | |||
| Europe | Britain | 14% | [24] | ||
| Belgium | 8% | [25] | |||
| Asia | Japan | 14.5–62.5% | [10,26] | ||
| 2000–2009 | America | Brazil | 14.91–68.6% | [35,36] | |
| Uruguay | 1.5–11.8% | [47] | |||
| Europe | Netherlands | 2.8% | [7] | ||
| Italy | 46.74% | [33] | |||
| Asia | Korea | 5.6–58.2% | [27,30,31,32] | ||
| Turkey | 10.8–28.1% | [28,29] | |||
| 2010–2019 | Oceania | Australia | 21.6% | [39] | |
| New Zealand | 14% | [38] | |||
| Africa | Algeria | 20.73% | [40] | ||
| Ghana | 0.3% | [8] | |||
| Asia | Iran | 7.2% | [41] | ||
| China | 12.20–69.05% | [11,45] | |||
| Thailand | 12% | [42] | |||
| India | 8.88–16% | [43] | |||
| Vietnam | 6.9% | [44] | |||
| Respiratory tract | Before 2000 | America | USA | 8.1–96% | [6,48] |
| Canada | 57.89–66.67% | [23,49] | |||
| 2000–2009 | Europe | Italy | 9.6–65.85% | [50] | |
| 2010–2019 | America | Brazil | 22–67% | [35,37] | |
| Europe | Ireland | 22.9–60.7% | [51] | ||
| France | 17–70.1% | [52,53] | |||
| Asia | Japan | 21.2% | [54] | ||
| Turkey | 1% | [28] | |||
| Oceania | Australia | 13–33.33% | [55] | ||
| After 2020 | Asia | China | 21.53% | [45] | |
| Serum antibody | America | USA | 11–91% | [22] | |
| Canada | 7–100% | [56,57] | |||
| Europe | Sweden | 23.8–100.0% | [58] | ||
| Norway | 16–72.2% | [59,60,61] | |||
| France | 16.5% | [62] | |||
| Belgium | 30% | [63] | |||
| Africa | Ghana | 55.8% | [8] |
| Codon | MEME p-Value | SLAC p-Value | FUBAR p-Value |
|---|---|---|---|
| 113 | 0.020 | 0.015 | 0.000 |
| 499 | 0.01 | 0.007 | 0.000 |
| 501 | 0.000 | 0.000 | 0.000 |
| 509 | 0.000 | 0.003 | 0.000 |
| 1238 | 0.000 | 0.028 | 0.041 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Q.; Li, B.; Sun, D. Advances in Bovine Coronavirus Epidemiology. Viruses 2022, 14, 1109. https://doi.org/10.3390/v14051109
Zhu Q, Li B, Sun D. Advances in Bovine Coronavirus Epidemiology. Viruses. 2022; 14(5):1109. https://doi.org/10.3390/v14051109
Chicago/Turabian StyleZhu, Qinghe, Bin Li, and Dongbo Sun. 2022. "Advances in Bovine Coronavirus Epidemiology" Viruses 14, no. 5: 1109. https://doi.org/10.3390/v14051109
APA StyleZhu, Q., Li, B., & Sun, D. (2022). Advances in Bovine Coronavirus Epidemiology. Viruses, 14(5), 1109. https://doi.org/10.3390/v14051109