1. Introduction
More than two years have passed since the beginning of the COVID-19 pandemic, occurring in late December 2019 in Wuhan, Hubei Province (China). When SARS-CoV-2 was identified as the causative agent of COVID-19, no one expected that the outbreak at the Huanan Seafood Market would grow into such a prolonged global pandemic. The origins of SARS-CoV-2 remain unknown, as does the exact date of the initial outbreak [
1]. As of 6 April 2022, 492,189,439 COVID-19 cases have been identified, with 6,159,474 fatalities [
2]. Despite a number of successes in the fight against the pandemic, including the creation of vaccines, specific antiviral drugs, and the optimization of treatment/prevention regimens, there is no hope for an end to the pandemic this year. Russia, as part of the global community, was also affected by the pandemic. The first COVID-19 patient in Russia was registered on 2 March 2020 [
3]. As of 6 April 2022, 17,940,765 COVID-19 cases have been identified in Russia, with 370,602 fatalities [
4].
Regional differences in the epidemic process occur due to Russia’s expansive geography and the uneven population of its different federal districts (FDs). In two of them, the Central FD and Northwestern FD, the majority of new COVID-19 cases are registered. In addition, the spread of emerging SARS-CoV-2 genetic variants usually begins in these regions. This is due to their high population densities, the presence of two megacities (Moscow in the Central FD and Saint Petersburg in the Northwestern FD), and to the fact that megacities are the largest transport hubs carrying the most international traffic.
Here, we present data characterizing the COVID-19 epidemic in Russia’s northwestern region (total population 13,941,919) in 2021. The epidemiological features of COVID-19 in this region are all the more interesting because a variant under monitoring (VUM), AT.1, was initially identified here [
3]. In our study, we used statistical data from Rospotrebnadzor as well as data from the genetic monitoring of SARS-CoV-2 implemented by the Saint Petersburg Pasteur Institute.
3. Results
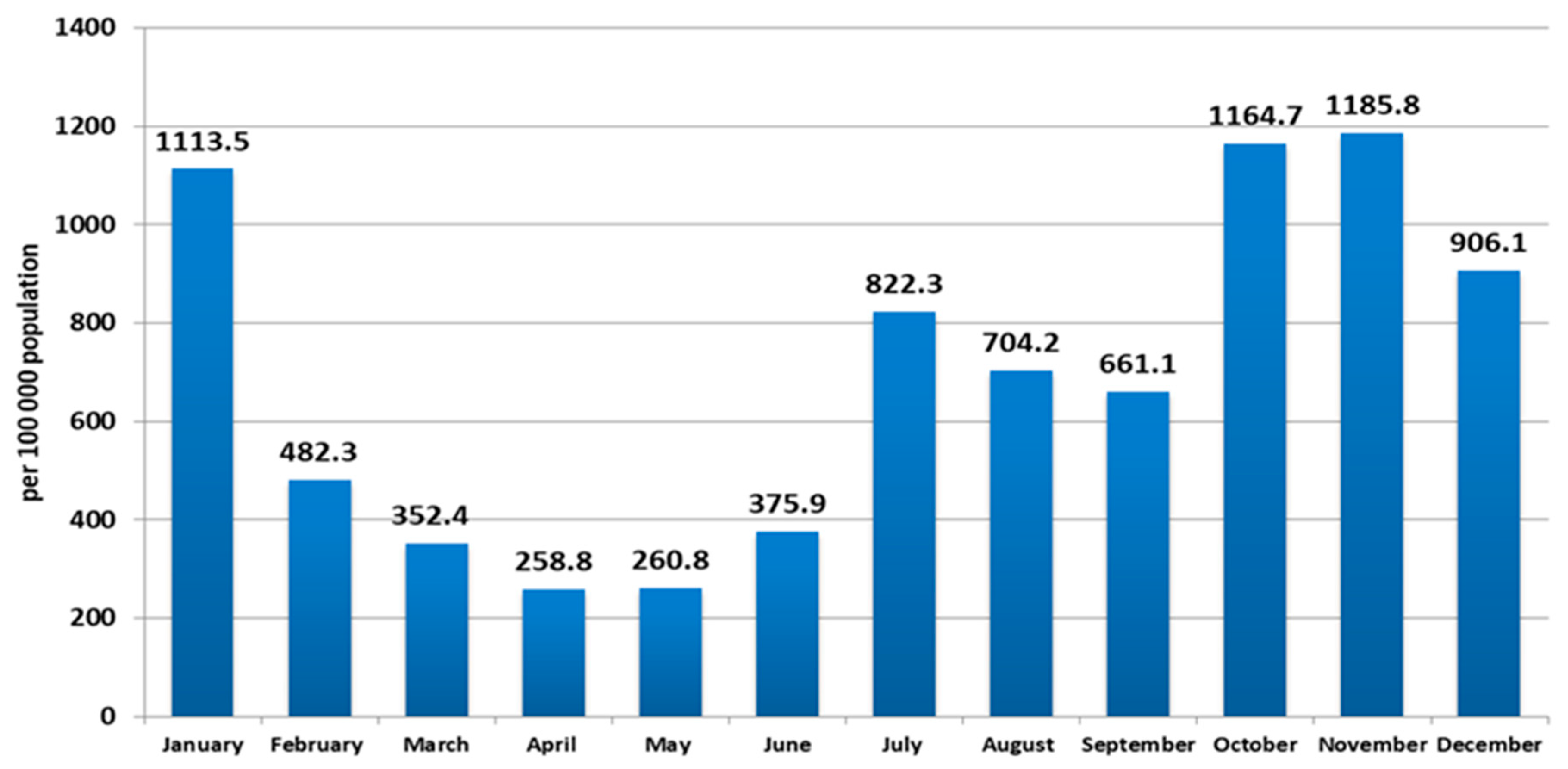
In 2021, COVID-19 incidence in northwest Russia varied in waves, ranging from 258.8 (per 100 K/month) in April to 1185.8 in November. At the same time, increases in incidence with the achievement of local maxima were recorded in January (1113.5 per 100 K/month), June (822.3 per 100 K/month), and November (1185.8 per 100 K/month) (
Figure 1).
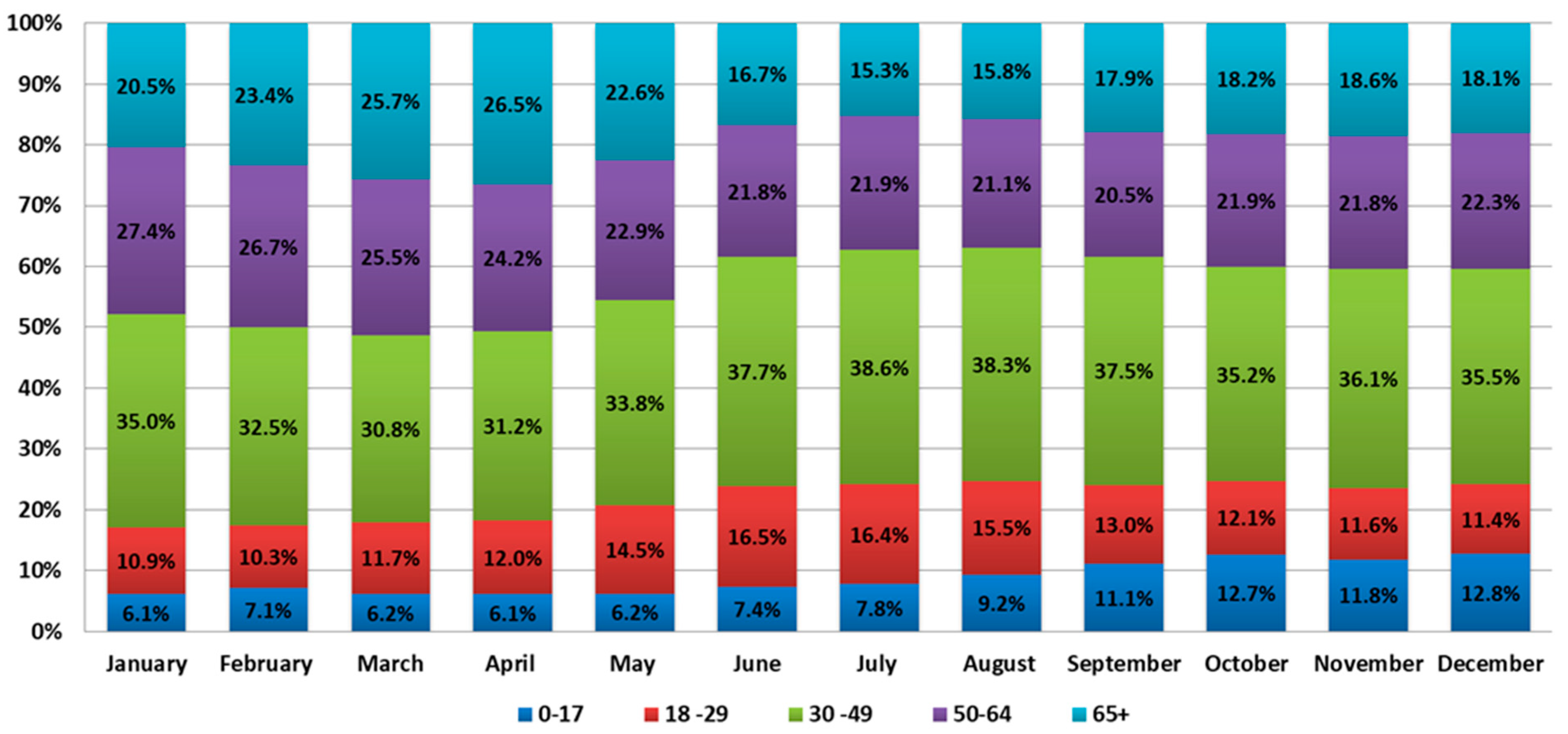
An analysis of the age structure of COVID-19 incidence showed the greatest fluctuations in the age group from 0 to 17 years. In January, COVID-19 incidence in this group was 6.1% of cases, while in December, the incidence was 12.8% of cases. During 2021, there was a gradual increase in incidence in this age group. In the other age groups at the same time, the fluctuations were not so significant and were as follows: from 10.9% to 16.5% in the age group 18–29 years; from 30.8% to 38.6% in the age group 30–49 years; from 20.5% to 27.4% in the age group 50–64 years; and from 15.3% to 26.5% in the group 65
+ year (
Figure 2).
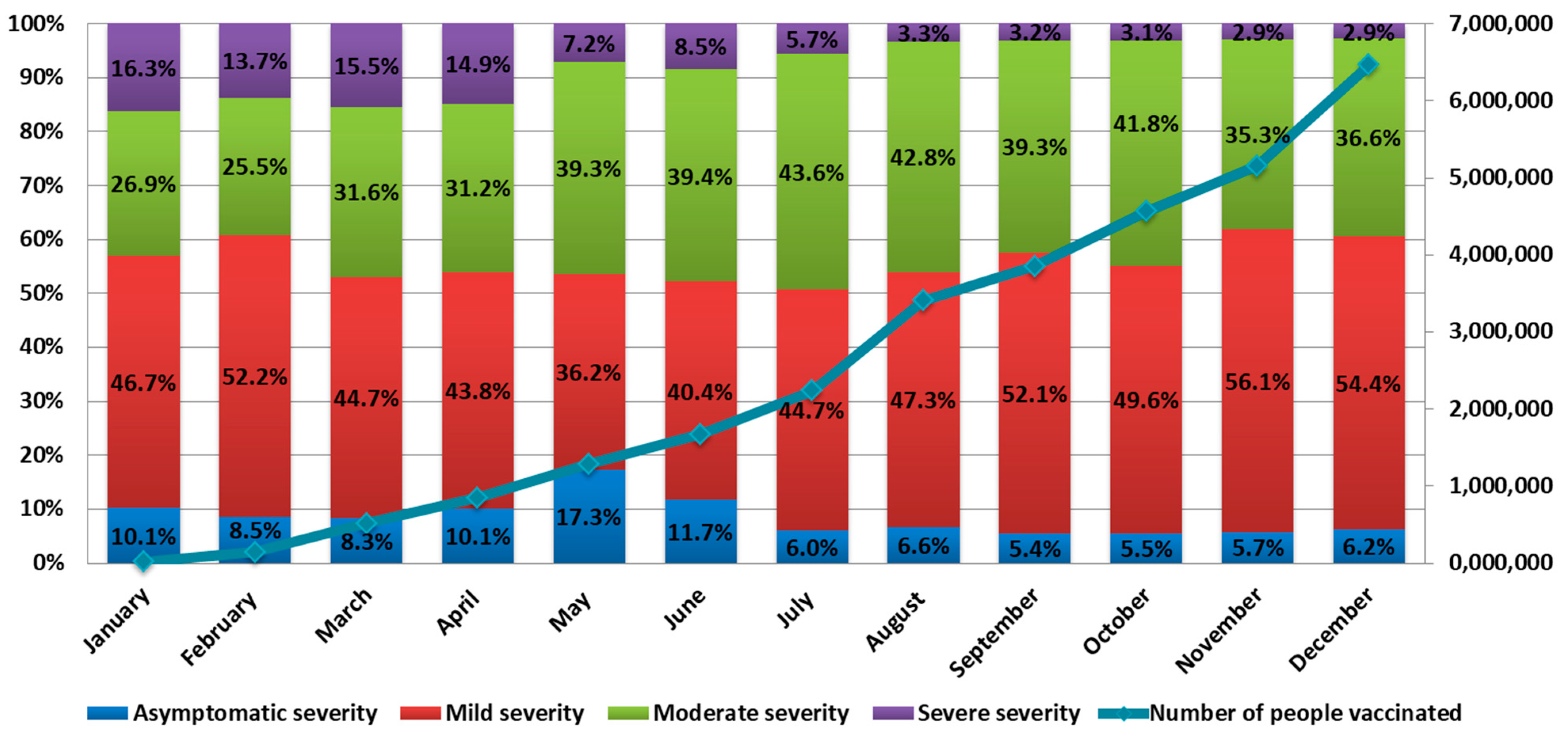
The number of cases of severe COVID-19 in 2021 ranged from 16.3% in January to 2.9% in December. The number of cases of asymptomatic infection ranged from 5.4% in September to 17.3% in May. Mild forms ranged from 36.2% in May to 56.1% in November. The number of COVID-19 cases with moderate symptoms ranged from 26.9% in January to 43.6% in July (
Figure 3).
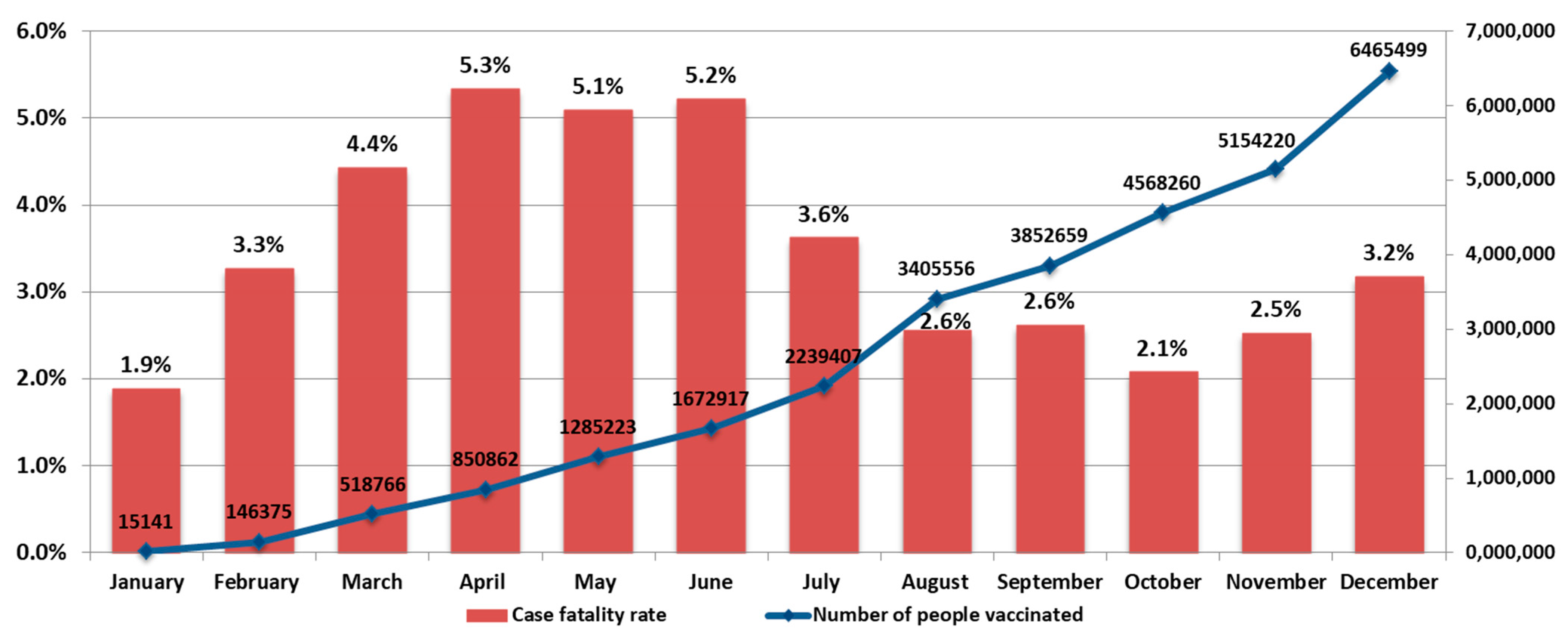
The COVID-19 mortality rate also varied in waves, ranging from 1.9% in January to 5.3% in April. Local maxima in mortality were seen in April (5.3%), September (2.6%), and December (3.2%) (
Figure 4).
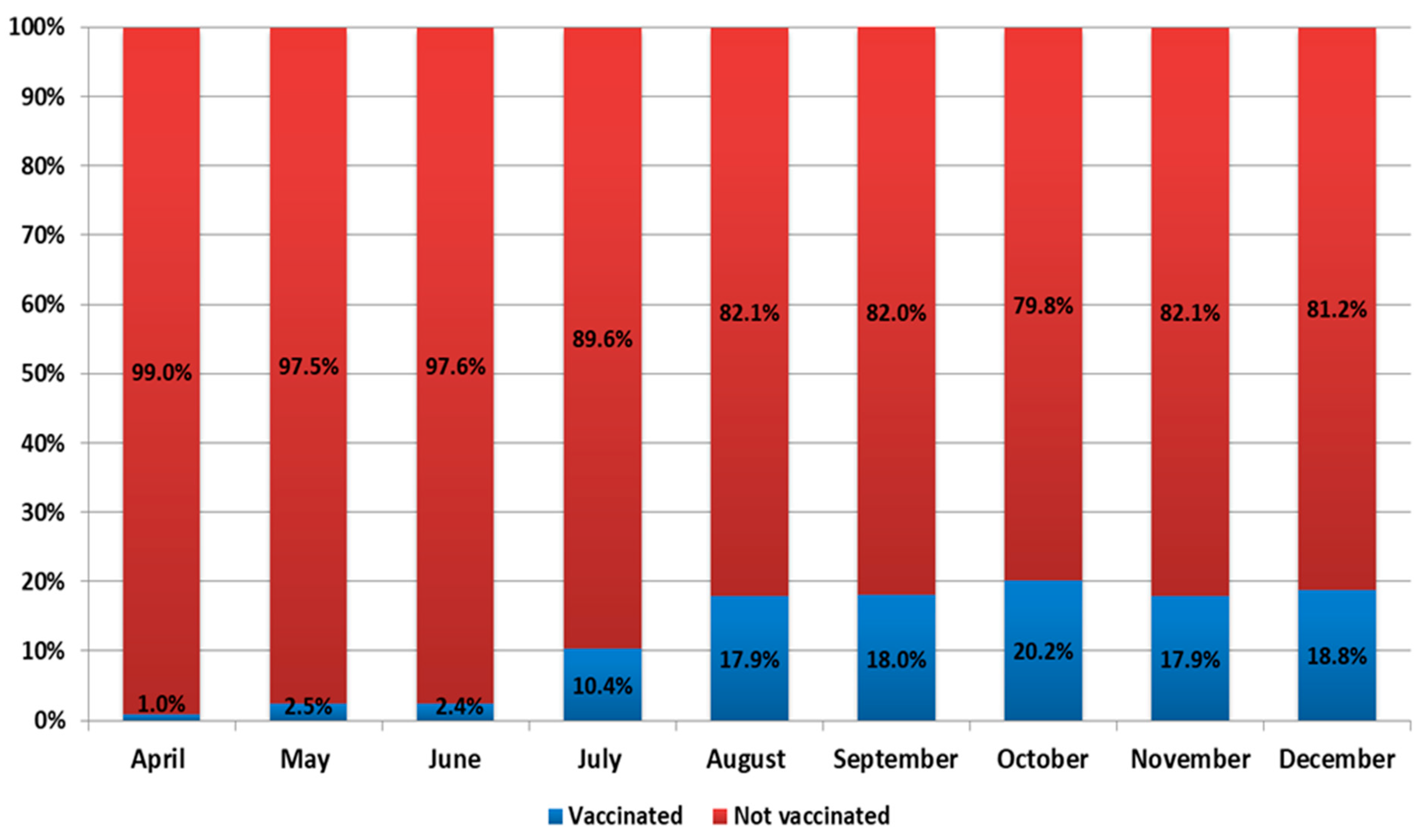
It should be noted that, during 2021, events developed against the background of a vaccination campaign. The number of vaccinated individuals increased from 15,141 in January to 6,465,499 in December (
Figure 4 and
Figure 5). In 2021, there was an increase in the share of samples from vaccinated individuals in the total volume of positive samples detected. For instance, the share of vaccinated persons among positive samples ranged from 1% in January to 18.8% in December (
Figure 5).
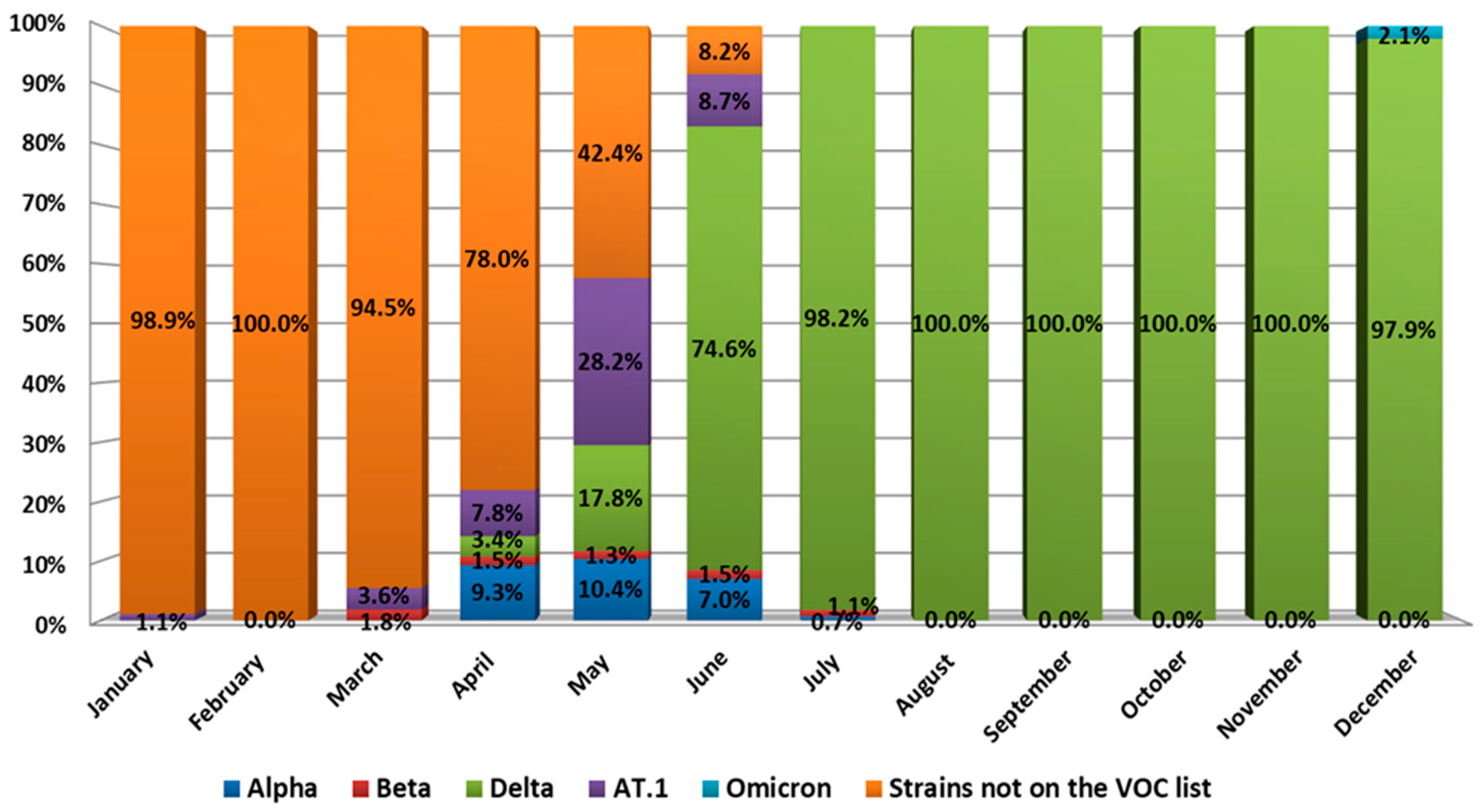
SARS-CoV-2 genetic diversity in northwest Russia also varied and was characterized by the presence of variants of concern (VOC) (
Figure 6).
During the first quarter of 2021, the 20B lineage SARS-CoV-2 variant dominated in northwest Russia. However, along with wild type, we detected the Beta SARS-CoV-2 variant, which is one of the so-called variants of concern (VOCs) [
13]. In addition, we detected the AT.1 SARS-CoV-2 variant in January 2021, which was admitted as a variant under monitoring (VUM) [
3]. Its prevalence in January reached 1.1%. It stayed low during the first ten days of 2021 but reached 3.6% in March.
The genetic landscape of SARS-CoV-2 changed in April. The share of wild type genetic variants dropped to 78%. At the same time, circulation of the Alpha and Delta variants of concern began, with Alpha reaching 9.3%. The share of Beta variants remained low as Delta variants appeared in the population. AT.1 prevalence also reached 7.8%.
A significant change in the genetic landscape began in May 2021. Displacement of the wild-type virus continued, and its prevalence decreased to 42.4%. At the same time, AT.1 prevalence reached 28.2%, while Delta variant prevalence was 17.8%. The Alpha variant constituted 10.4% of all genetic variants in May. Such Delta variant dynamics led to the fact that, by the end of June, there was almost a complete displacement of all other SARS-CoV-2 variants. Thus, Delta variant prevalence in July was 97.1%, while wild type (20B) prevalence was only 2.9%.
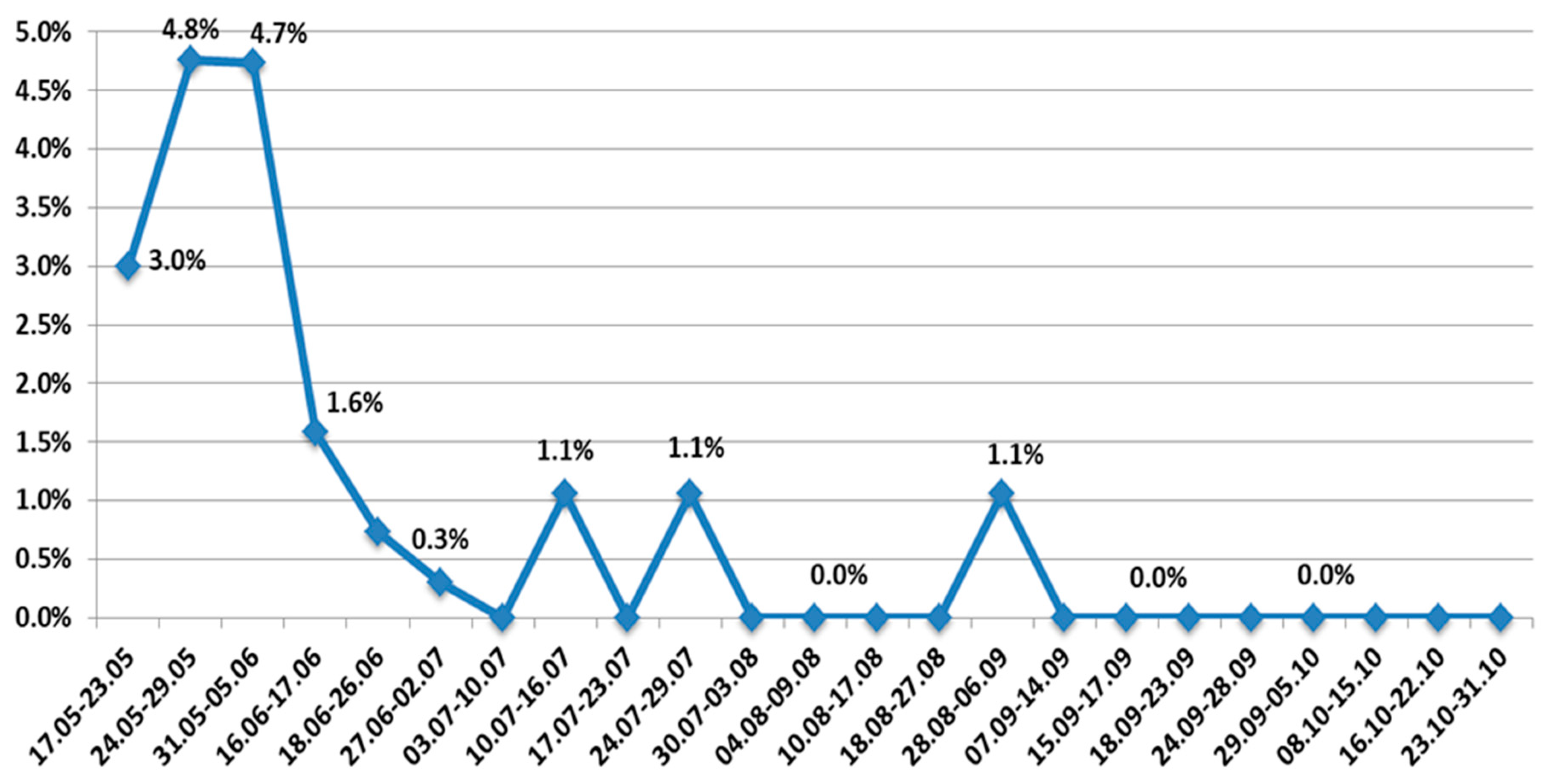
From the beginning of July to the middle of December, a complete dominance of the Delta variant was noted. Other VOCs, as well as wild type SARS-CoV-2, were eliminated. At the same time, the AT.1 SARS-CoV-2 variant continued to circulate sporadically until early September 2021 (
Figure 7).
The variant ceased to circulate widely with the advent of the Delta variant, although AT.1 continued to be sporadically detected until September 2021. In mid-December, an Omicron SARS-CoV-2 variant was identified in northwest Russia. By the end of 2021, Omicron prevalence was 2.1%, while Delta prevalence was 97.9%.
3.1. SARS-CoV-2 Genetic Diversity in Northwest Russia
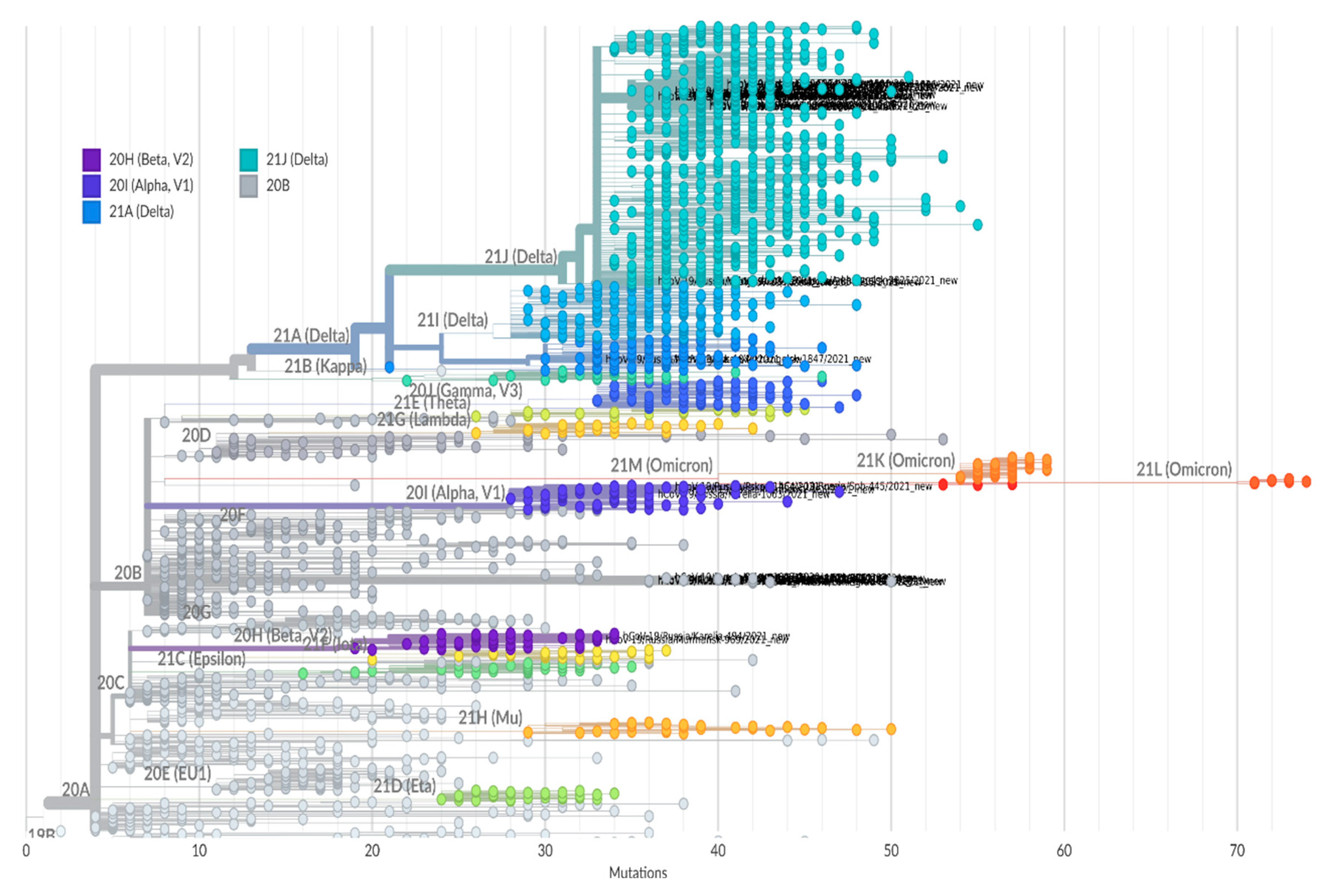
Genomic analysis based on Nextclade SARS-CoV-2 Clade Assigner showed seven lineages (
Figure 8) circulating in the region in 2021: 20A, 20B, 20H (Beta); 20I (Alpha); 21A (Delta); 21J (Delta); and 21K (Omicron). Since July, the majority of isolates belonged to 21J (Delta).
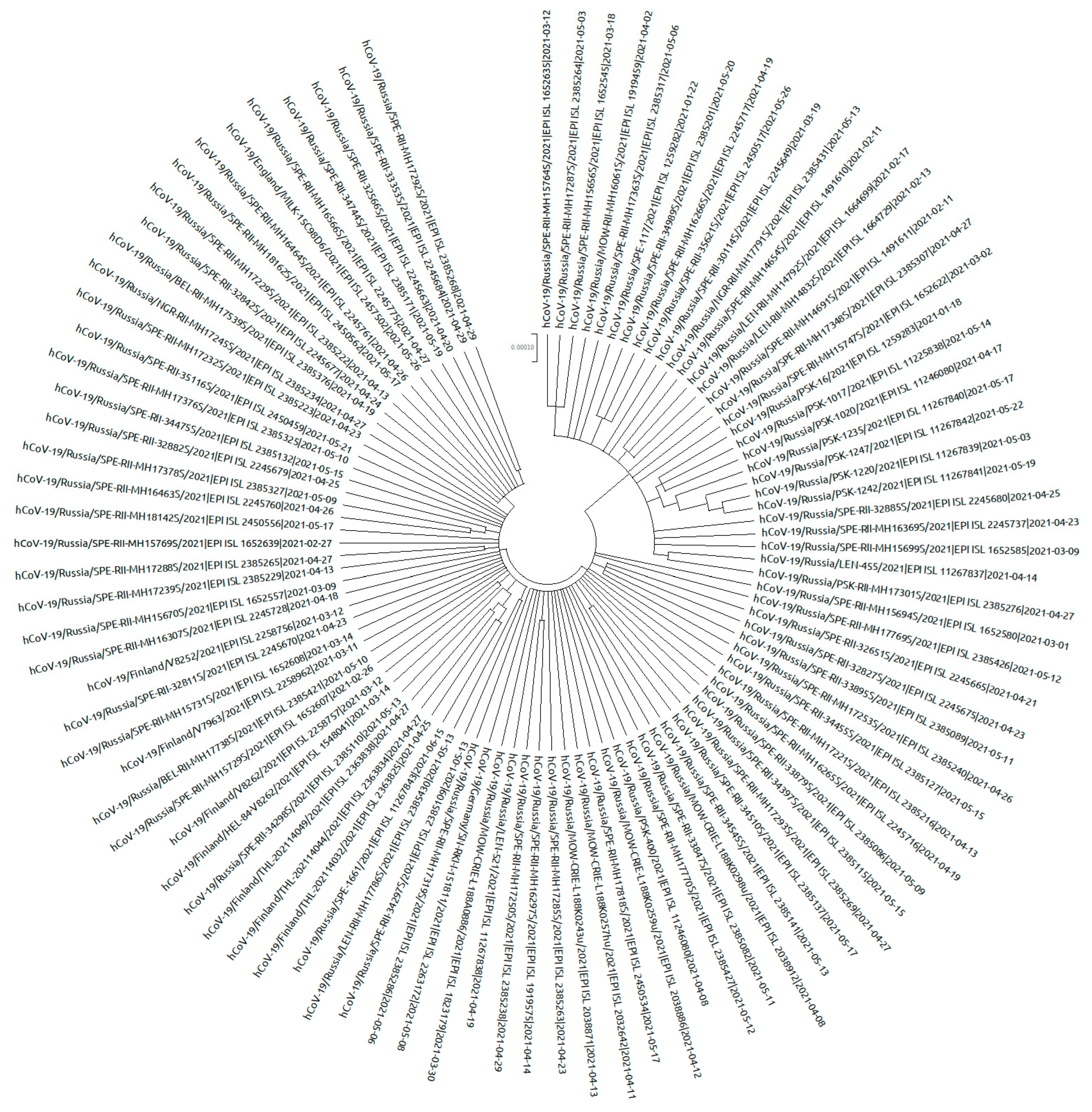
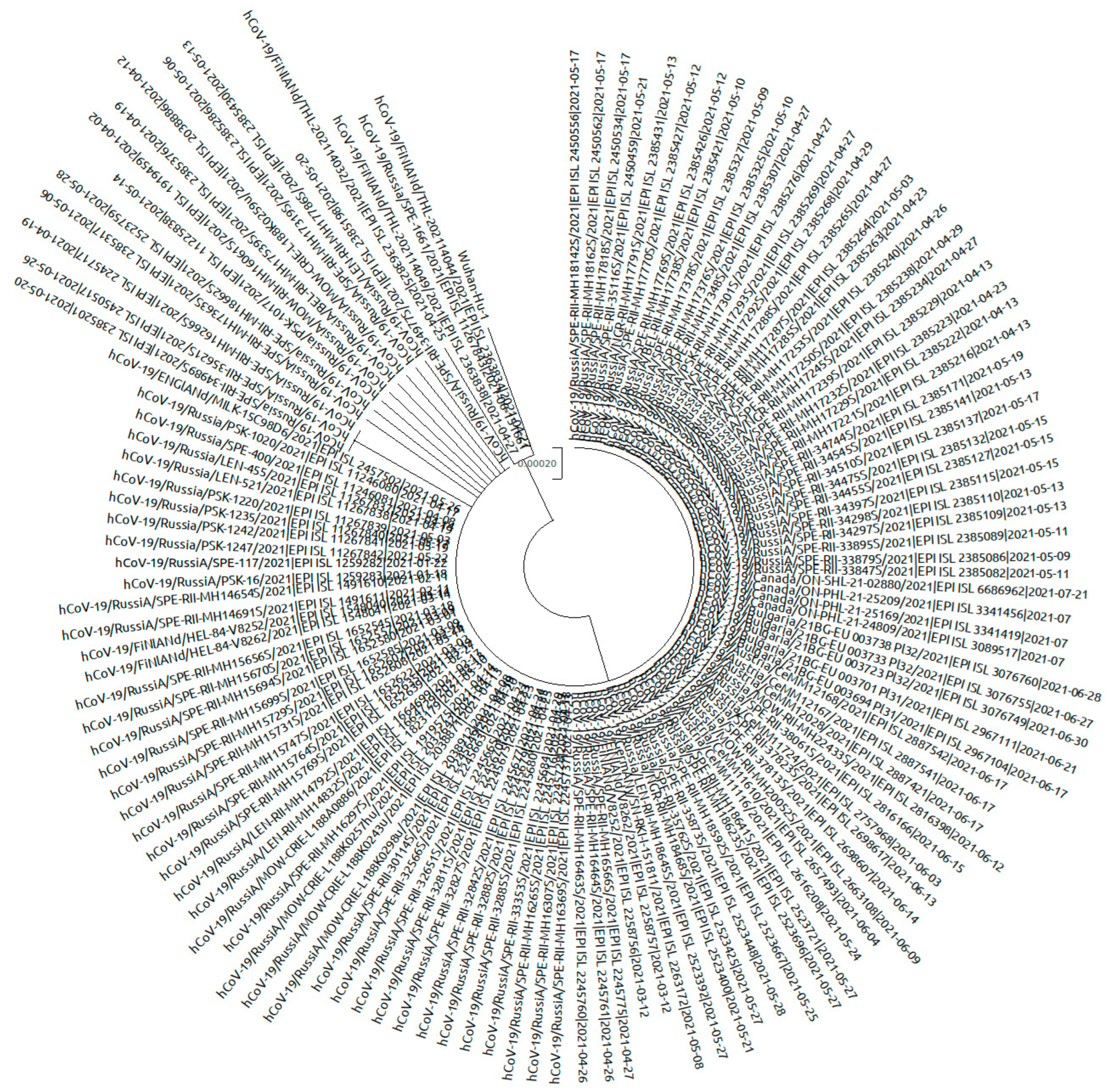
3.2. Phylogeny of the AT.1 Lineage
Phylogenetic analysis of AT.1 strains was performed with all of the sequences available in the GISAID database. According to the Pangoline database [
14], AT.1 isolates were identified in Russia, Finland, Estonia, Lithuania, Bulgaria, Austria, Germany, and Belgium. The AT.1 lineage does not form a separate cluster on the global tree of SARS-CoV-2 strains (
Figure 8) but forms a long lineage within clade 20B.
The AT.1 genetic variant is characterized by the presence of five non-synonymous substitutions (P9L, D215G, H245P, E484K, E780K) as well as a long deletion (136–144) and an insertion (679) in the S gene. Analysis of 150 AT.1 sequences from GISAID has shown that not all of them share these mutations or characteristic insertion and deletion. For this reason, only sequences with characteristic insertion and deletion (127 sequences) were chosen for phylogenetic analysis. In total, 25 variable sites were identified, 7 of which were non-singleton. Translated S gene sequence had 16 substitutions, 3 of which were non-singleton. Mutations D215G and H245P were not universal: 5.5% of sequences contained D at position 215 like the reference genome S gene; and 3.9% of sequences contained H at position 245 like the reference genome. Position 614 featured G or D variability.
The phylogenetic tree was constructed with sequences from Russia, Finland, Germany, and England. Intra-lineage diversity represented: up to 48 nucleotides in difference between each pair of sequences per genome alignment; and up to 5 nucleotides in difference between each pair of sequences within S gene alignment. On the phylogenetic tree, sequences from northwest Russia (Saint Petersburg, Leningrad region, Novgorod region, Pskov region) were mixed together with sequences from other regions (Moscow, Belgorod region, Yaroslavl region) and sequences from outside of Russia (
Figure 9 and
Figure 10).
3.3. Variant of Concern Genetic Diversity in the Northwest Region
For 20I (Alpha) variants, 254 sequences from Russia were analyzed in total. Pairwise distance was up to 26 nucleotides per genome and up to 6 nucleotides for the S gene. Within S gene sequences from Russia, 114 variable sites were identified, of which 29 were non-singletons. For amino acid alignment, 62 variable sites were identified, of which 16 were non-singletons. For sequences from northwest Russia (75 sequences), 34 nucleotide sites were variable, 8 of which were non-singletons. Regarding amino acid sequence, there were 18 amino acid substitutions 5 of which were singletons. Non-singleton substitutions were at S gene positions 67 (A/V), 502 (Y/N), 571 (A/D), 615 (G/D), and 846 (A/S).
On the global phylogenetic tree, two sequences from northwest Russia obtained within this work (CoV-19/Russia/SPE-445/2021, hCoV-19/Russia/PSK-1464/2021) were clustered with sequences from Germany (Germany/NW-RKI-I-093558/2021). Sequence hCoV-19/Russia/KR-1063/2021 clustered with sequences from England and Denmark. Sequence hCoV-19/Russia/MUR-466/2021 clustered with mixed sequences from Europe, and sequence hCoV-19/Russia/VLG-1249/2021 was located more closely to an isolate from England (England/PORT-2E5324/2021).
For 20H (Beta) variants, 33 sequences from Russia were analyzed. Pairwise distance was up to 22 substitutions per genome and up to 6 substitutions in the S gene. Within S gene sequences from Russia, 16 variable sites were identified, 4 of which were non-singletons. For amino acid alignment, 10 variable sites were seen, of which 3 were non-singletons. For sequences from northwest Russia (8 sequences), 4 nucleotide sites were variable, 2 of which were non-singletons. Regarding amino acid sequences, there were 2 amino acid substitutions, one of which was singleton. The non-singleton substitution was at position 18 (F/L) in the S gene.
On the global phylogenetic tree, one sequence from northwest Russia obtained within this work (hCoV-19/Russia/MUR-969/2021) was located in the cluster with sequences from the USA, South Africa, and Germany. Another (hCoV-19/Russia/KR-494/2021) was located in the cluster with sequences from Wales and England.
The first Delta strains began to appear in the northwest region in April, belonging to two clades: 21A and 21J. The within-genome pairwise difference for 21A was 11 nucleotides and up to 2 nucleotides for the S gene. For sequences from northwest Russia (6 sequences), 21 nucleotide sites were variable within the S gene, 1 of which was non-singleton. Regarding the amino acid sequence, there were 2 amino acid substitutions, 1 of which was non-singleton. The non-singleton substitution was at position 950 (N/D) of the S gene.
For 21J, pairwise differences were up to 15 nucleotides for the complete genome and up to 4 nucleotides for the S gene. For sequences from northwest Russia (56 sequences), 23 nucleotide sites were variable within the S gene, 3 of which were non-singletons. Regarding amino acid sequences, there were 20 amino acid substitutions, 3 of which were singletons. Non-singleton substitutions were at S gene positions 19 (T/R), 614 (D/G), and 950 (N/D).
Sequences from clade 21A were closely related to the Canada/MB-NML-86772/2021 sequence. Sequences from 21J were more abundant. On the SARS-CoV-2 global tree, they formed two groups: one located in a basal position of 21J (closest relative England/NORT-1BD1AFD/2021); and the other forming a subcluster (closest relative Finland/THL-202122712/2021) on the tree.
Figure 9.
Phylogenetic tree of AT.1 SARS-CoV-2 variants. Sequences (98) from Russia and Europe were included in the analysis. The TN93 substitution model was chosen as optimal for 19,205 bp without gaps/missing data alignment.
Figure 9.
Phylogenetic tree of AT.1 SARS-CoV-2 variants. Sequences (98) from Russia and Europe were included in the analysis. The TN93 substitution model was chosen as optimal for 19,205 bp without gaps/missing data alignment.
Figure 10.
Phylogenetic tree of AT.1 SARS-CoV-2 variants based on S gene sequence alignment. Sequences (127) from Russia and Europe were included in the analysis. The TN92 substitution model was chosen as optimal. Wuhan-Hu-1 sequence used as outgroup (GenBank: NC_045512).
Figure 10.
Phylogenetic tree of AT.1 SARS-CoV-2 variants based on S gene sequence alignment. Sequences (127) from Russia and Europe were included in the analysis. The TN92 substitution model was chosen as optimal. Wuhan-Hu-1 sequence used as outgroup (GenBank: NC_045512).
4. Discussion
Systematic monitoring of key epidemiological indicators is an integral part of anti-epidemic measures, both at the regional and national levels. The modern development of molecular technologies has made it possible to study the genetic variability of SARS-CoV-2 and link it with epidemiological indicators since the beginning of the COVID-19 pandemic. All viruses, including SARS-CoV-2, change their genetic structure during circulation. Most events associated with a change in genetic sequence do not lead to changes in the biological properties of the pathogen. However, some mutations can cause changes in a number of properties, such as infectious activity, pathogenicity, the ability to escape post-vaccination immunity, etc.
As a result of intense and widespread circulation, SARS-CoV-2 genetic variants, which pose an increased risk to global health, began to appear in the second half of 2020. This circumstance prompted the WHO to introduce warning classifications for such SARS-CoV-2 genetic variants. It is in this context that the VOC, VOI, and VUM definitions appeared. Such classification made it possible to create guidelines for evaluating SARS-CoV-2 genetic monitoring data. It also contributed to the adoption of optimal management solutions related to anti-epidemic measures.
The appearance of new genetic variants is a natural result of SARS-CoV-2 evolution. At the same time, some variants, being more aggressive, are able to displace others. Such features were first noted with the British genetic variant, which appeared in England in the autumn of 2020. From the beginning of 2021, it began to rapidly spread around the world and displace genetic variants circulating at that time [
15]. Being the first VOC, the British (Alpha) genetic variant was defined by the presence of 23 nucleotide mutations across the genome that map to a single branch of the phylogenetic tree [
16]. The presence of a wide range of mutations, including in the RBD (the site responsible for cellular ACE2 receptor binding), contributed to its increased infectious activity and a change in the clinical manifestation of COVID-19 [
15,
17]. The Alpha SARS-CoV-2 variant was obtained in northwest Russian at the end of 2020. This VOC spread throughout the region and accounted for up to 10% of sequenced samples by May 2021.
The first variant capable of totally displacing others was the Delta genetic variant. It was first discovered in October 2020 in India and demonstrated the highest transmissibility rate seen during the study period. Despite the fact that the previous variants of concern (Alpha, Beta, Gamma) had spike protein mutations, the presence of a special mutation signature (L452R, T478K, P681R) in the S protein made it very highly transmissible. The transmissivity of this strain is 97% higher than that of the Wuhan variant [
18]. By September 2021, the dominance of the Delta genetic variant was widespread. In the northwestern region, the Delta variant totally displaced the others by July 2021 and totally dominated for six months. The Omicron genetic variant was detected in the northwestern region in December 2021 and is rapidly spreading around the world at the present time.
Other variants are less aggressive than VOCs, but their occurrence indicates local signs of viral evolution. Hence, after appearing in the northwestern region, a variant belonging to the AT.1 lineage reached 28.2% of the viral population in the region and actively competed with other genetic variants circulating at that time. Only the appearance of the Delta genetic variant, which totally displaced all others, reduced AT.1 and led to elimination in the region, although sporadic cases were noted until September 2021.
This paper presents a comprehensive analysis of the COVID-19 situation in northwest Russia in 2021, including not only epidemiological indicators but also data on genetic variants. Epidemiological data were taken from a non-public Rospotrebnadzor database that is updated daily, which allows for the objective assessment of the epidemiological situation at specific time points. The genetic structure of the viral population, on the contrary, cannot be studied in its entirety due to the limited selection of strains suitable for sequencing. This, to some degree, limits the identification of rare variants in the viral population. The appearance of the AT.1 variant in the region makes clear that unnoticed sporadic appearance or disappearance of other rare genetic variants cannot be ruled out.
The epidemic process in northwest Russia was characterized by periodic rises and falls in incidence. These could be associated with, among other things, the predominance of certain SARS-CoV-2 genetic variants. The epidemic course continued alongside increasing vaccination, which has been actively conducted since December 2020, when vaccines were developed in Russia [
19,
20,
21]. By the end of 2021, 45.7% of the Russian population (66,540,512 people) were vaccinated. At the same time, 46.4% of the population (6,465,494 people) were vaccinated in Russia’s Northwestern Federal District [
22]. It should be noted that the majority of people were vaccinated with
Gam-COVID-Vac, better known as Sputnik V [
19]. In this context, there were decreases in the mortality rate and in the severity of COVID-19 clinical manifestation. Thus, while vaccination could not completely stop the epidemic process, it nevertheless had a positive effect on it. This effect persists, to one degree or another, despite a decrease in the strength of post-vaccination immunity against newly emerging SARS-CoV-2 variants. The reduction in the risk of SARS-CoV-2 infection in vaccinated persons is indirectly indicated by the fact that, during 2021, the share of children among COVID-19 cases (who did not participate in the immunization program) increased by twofold.

 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}