
Henipavirus Immune Evasion and Pathogenesis Mechanisms: Lessons Learnt from Natural Infection and Animal Models



Abstract
:1. Generalities
1.1. Genomic Features
1.2. Biological Features
1.3. Bats: Natural Reservoir of Henipaviruses
2. Pathogenesis
2.1. Transmission
2.2. Pathogenesis of Henipaviruses
Henipavirus Immune Responses in Humans
2.3. Pathogenesis in Accidental Hosts
2.3.1. Pathogenesis in Horses
2.3.2. Pathogenesis in Pigs
2.3.3. Pathogenesis in Humans
3. Immune Responses in Bats as Reservoir Hosts for Henipaviruses
3.1. Biological Factors
3.2. Immune Factors
3.2.1. Immune Cell Populations
3.2.2. The Innate Immune Response
Pattern Recognition Receptors
Interferon and Soluble Immune Mediators
NK Cells
3.2.3. Adaptive Immune Response
Antibodies
T-Cell Responses
3.3. Conclusions
4. Animal Models
4.1. Henipavirus Infection in Cats
Henipavirus Immune Responses in Cats
4.2. Henipavirus Infection in Dogs
4.3. Experimental Henipavirus Infection in Ferrets
Henipavirus Immune Responses in Ferrets
4.4. Henipavirus Infection in Horses
Henipavirus Immune Responses in Horses
4.5. Experimental Henipavirus Infection in Monkey Models
Henipavirus Immune Responses in Monkey Models
4.6. Henipavirus Infection in Pigs
4.6.1. NiV Infection in Pigs
4.6.2. HeV Infection in Pigs
Henipavirus Immune Responses in Pigs
4.7. Experimental Henipavirus Infection in Mice
Henipavirus Immune Responses in Mice
4.8. Experimental Henipavirus Infection in Hamsters
Henipavirus Immune Responses in Hamsters
4.9. Experimental Henipavirus Infection in Guinea Pigs
Henipavirus Immune Responses in Guinea Pigs
4.10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eaton, B.T.; Broder, C.C.; Middleton, D.; Wang, L.F. Hendra and Nipah viruses: Different and dangerous. Nat. Rev. Microbiol. 2006, 4, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Halpin, K.; Rota, P. A Review of Hendra Virus and Nipah Virus Infections in Man and Other Animals. Zoonoses Infect. Affect. Hum. Anim. Focus Public Health Asp. 2014, 997–1012. [Google Scholar] [CrossRef]
- Aditi; Shariff, M. Nipah virus infection: A review. Epidemiol. Infect. 2019, 147, e95. [Google Scholar] [CrossRef] [PubMed]
- Jack, P.J.; Boyle, D.B.; Eaton, B.T.; Wang, L.F. The complete genome sequence of J virus reveals a unique genome structure in the family Paramyxoviridae. J. Virol. 2005, 79, 10690–10700. [Google Scholar] [CrossRef] [Green Version]
- Harcourt, B.H.; Tamin, A.; Halpin, K.; Ksiazek, T.G.; Rollin, P.E.; Bellini, W.J.; Rota, P.A. Molecular characterization of the polymerase gene and genomic termini of Nipah virus. Virology 2001, 287, 192–201. [Google Scholar] [CrossRef] [Green Version]
- Harcourt, B.H.; Tamin, A.; Ksiazek, T.G.; Rollin, P.E.; Anderson, L.J.; Bellini, W.J.; Rota, P.A. Molecular characterization of Nipah virus, a newly emergent paramyxovirus. Virology 2000, 271, 334–349. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.F.; Michalski, W.P.; Yu, M.; Pritchard, L.I.; Crameri, G.; Shiell, B.; Eaton, B.T. A novel P/V/C gene in a new member of the Paramyxoviridae family, which causes lethal infection in humans, horses, and other animals. J. Virol. 1998, 72, 1482–1490. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Harcourt, B.H.; Yu, M.; Tamin, A.; Rota, P.A.; Bellini, W.J.; Eaton, B.T. Molecular biology of Hendra and Nipah viruses. Microbes Infect. 2001, 3, 279–287. [Google Scholar] [CrossRef]
- Drexler, J.F.; Corman, V.M.; Muller, M.A.; Maganga, G.D.; Vallo, P.; Binger, T.; Gloza-Rausch, F.; Cottontail, V.M.; Rasche, A.; Yordanov, S.; et al. Bats host major mammalian paramyxoviruses. Nat. Commun. 2012, 3, 796. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.F.; Yu, M.; Hansson, E.; Pritchard, L.I.; Shiell, B.; Michalski, W.P.; Eaton, B.T. The exceptionally large genome of Hendra virus: Support for creation of a new genus within the family Paramyxoviridae. J. Virol. 2000, 74, 9972–9979. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, M.; Setiyono, A.; Handharyani, E.; Rahmadani, I.; Taha, S.; Adiani, S.; Subangkit, M.; Sawa, H.; Nakamura, I.; Kimura, T. Molecular detection of a novel paramyxovirus in fruit bats from Indonesia. Virol. J. 2012, 9, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yob, J.M.; Field, H.; Rashdi, A.M.; Morrissy, C.; van der Heide, B.; Rota, P.; bin Adzhar, A.; White, J.; Daniels, P.; Jamaluddin, A.; et al. Nipah virus infection in bats (order Chiroptera) in peninsular Malaysia. Emerg. Infect. Dis. 2001, 7, 439–441. [Google Scholar] [CrossRef] [PubMed]
- Reynes, J.M.; Counor, D.; Ong, S.; Faure, C.; Seng, V.; Molia, S.; Walston, J.; Georges-Courbot, M.C.; Deubel, V.; Sarthou, J.L. Nipah virus in Lyle’s flying foxes, Cambodia. Emerg. Infect. Dis. 2005, 11, 1042–1047. [Google Scholar] [CrossRef] [PubMed]
- Mohd Nor, M.N.; Gan, C.H.; Ong, B.L. Nipah virus infection of pigs in peninsular Malaysia. Rev. Sci. Tech. 2000, 19, 160–165. [Google Scholar] [CrossRef]
- Wong, K.T.; Shieh, W.J.; Zaki, S.R.; Tan, C.T. Nipah virus infection, an emerging paramyxoviral zoonosis. Springer Semin. Immunopathol. 2002, 24, 215–228. [Google Scholar] [CrossRef]
- Chowdhury, S.; Khan, S.U.; Crameri, G.; Epstein, J.H.; Broder, C.C.; Islam, A.; Peel, A.J.; Barr, J.; Daszak, P.; Wang, L.F.; et al. Serological evidence of henipavirus exposure in cattle, goats and pigs in Bangladesh. PLoS Negl. Trop. Dis. 2014, 8, e3302. [Google Scholar] [CrossRef] [Green Version]
- Middleton, D.J.; Westbury, H.A.; Morrissy, C.J.; van der Heide, B.M.; Russell, G.M.; Braun, M.A.; Hyatt, A.D. Experimental Nipah virus infection in pigs and cats. J. Comp. Pathol. 2002, 126, 124–136. [Google Scholar] [CrossRef]
- Middleton, D. Hendra virus. Vet. Clin. N. Am. Equine Pract. 2014, 30, 579–589. [Google Scholar] [CrossRef]
- Luby, S.P.; Gurley, E.S.; Hossain, M.J. Transmission of human infection with Nipah virus. Clin. Infect. Dis. 2009, 49, 1743–1748. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.T.; Robertson, T.; Ong, B.B.; Chong, J.W.; Yaiw, K.C.; Wang, L.F.; Ansford, A.J.; Tannenberg, A. Human Hendra virus infection causes acute and relapsing encephalitis. Neuropathol. Appl. Neurobiol. 2009, 35, 296–305. [Google Scholar] [CrossRef]
- Williamson, M.M.; Hooper, P.T.; Selleck, P.W.; Westbury, H.A.; Slocombe, R.F. A guinea-pig model of Hendra virus encephalitis. J. Comp. Pathol. 2001, 124, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.T.; Grosjean, I.; Brisson, C.; Blanquier, B.; Fevre-Montange, M.; Bernard, A.; Loth, P.; Georges-Courbot, M.C.; Chevallier, M.; Akaoka, H.; et al. A golden hamster model for human acute Nipah virus infection. Am. J. Pathol. 2003, 163, 2127–2137. [Google Scholar] [CrossRef] [Green Version]
- Dhondt, K.P.; Mathieu, C.; Chalons, M.; Reynaud, J.M.; Vallve, A.; Raoul, H.; Horvat, B. Type I interferon signaling protects mice from lethal henipavirus infection. J. Infect. Dis. 2013, 207, 142–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisbert, T.W.; Daddario-DiCaprio, K.M.; Hickey, A.C.; Smith, M.A.; Chan, Y.P.; Wang, L.F.; Mattapallil, J.J.; Geisbert, J.B.; Bossart, K.N.; Broder, C.C. Development of an acute and highly pathogenic nonhuman primate model of Nipah virus infection. PLoS ONE 2010, 5, e10690. [Google Scholar] [CrossRef] [PubMed]
- Pernet, O.; Wang, Y.E.; Lee, B. Henipavirus receptor usage and tropism. Curr. Top. Microbiol. Immunol. 2012, 359, 59–78. [Google Scholar] [CrossRef] [Green Version]
- Bossart, K.N.; Tachedjian, M.; McEachern, J.A.; Crameri, G.; Zhu, Z.; Dimitrov, D.S.; Broder, C.C.; Wang, L.F. Functional studies of host-specific ephrin-B ligands as Henipavirus receptors. Virology 2008, 372, 357–371. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.T.; Shieh, W.J.; Kumar, S.; Norain, K.; Abdullah, W.; Guarner, J.; Goldsmith, C.S.; Chua, K.B.; Lam, S.K.; Tan, C.T.; et al. Nipah virus infection: Pathology and pathogenesis of an emerging paramyxoviral zoonosis. Am. J. Pathol. 2002, 161, 2153–2167. [Google Scholar] [CrossRef]
- Reperant, L.A.; Brown, I.H.; Haenen, O.L.; de Jong, M.D.; Osterhaus, A.D.; Papa, A.; Rimstad, E.; Valarcher, J.F.; Kuiken, T. Companion Animals as a Source of Viruses for Human Beings and Food Production Animals. J. Comp. Pathol. 2016, 155 (Suppl. 1), S41–S53. [Google Scholar] [CrossRef] [Green Version]
- Cleaveland, S.; Laurenson, M.K.; Taylor, L.H. Diseases of humans and their domestic mammals: Pathogen characteristics, host range and the risk of emergence. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 991–999. [Google Scholar] [CrossRef]
- McKee, C.D.; Islam, A.; Luby, S.P.; Salje, H.; Hudson, P.J.; Plowright, R.K.; Gurley, E.S. The Ecology of Nipah Virus in Bangladesh: A Nexus of Land-Use Change and Opportunistic Feeding Behavior in Bats. Viruses 2021, 13, 169. [Google Scholar] [CrossRef]
- Drexler, J.F.; Geipel, A.; Konig, A.; Corman, V.M.; van Riel, D.; Leijten, L.M.; Bremer, C.M.; Rasche, A.; Cottontail, V.M.; Maganga, G.D.; et al. Bats carry pathogenic hepadnaviruses antigenically related to hepatitis B virus and capable of infecting human hepatocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 16151–16156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, J.H.; Quan, P.L.; Briese, T.; Street, C.; Jabado, O.; Conlan, S.; Ali Khan, S.; Verdugo, D.; Hossain, M.J.; Hutchison, S.K.; et al. Identification of GBV-D, a novel GB-like flavivirus from old world frugivorous bats (Pteropus giganteus) in Bangladesh. PLoS Pathog. 2010, 6, e1000972. [Google Scholar] [CrossRef] [PubMed]
- Quan, P.L.; Firth, C.; Conte, J.M.; Williams, S.H.; Zambrana-Torrelio, C.M.; Anthony, S.J.; Ellison, J.A.; Gilbert, A.T.; Kuzmin, I.V.; Niezgoda, M.; et al. Bats are a major natural reservoir for hepaciviruses and pegiviruses. Proc. Natl. Acad. Sci. USA 2013, 110, 8194–8199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teeling, E.C.; Springer, M.S.; Madsen, O.; Bates, P.; O’Brien, S.J.; Murphy, W.J. A molecular phylogeny for bats illuminates biogeography and the fossil record. Science 2005, 307, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Pfefferle, S.; Oppong, S.; Drexler, J.F.; Gloza-Rausch, F.; Ipsen, A.; Seebens, A.; Muller, M.A.; Annan, A.; Vallo, P.; Adu-Sarkodie, Y.; et al. Distant relatives of severe acute respiratory syndrome coronavirus and close relatives of human coronavirus 229E in bats, Ghana. Emerg. Infect. Dis. 2009, 15, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.; Woo, P.C.; Li, K.S.; Huang, Y.; Tsoi, H.W.; Wong, B.H.; Wong, S.S.; Leung, S.Y.; Chan, K.H.; Yuen, K.Y. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc. Natl. Acad. Sci. USA 2005, 102, 14040–14045. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Shi, Z.; Yu, M.; Ren, W.; Smith, C.; Epstein, J.H.; Wang, H.; Crameri, G.; Hu, Z.; Zhang, H.; et al. Bats are natural reservoirs of SARS-like coronaviruses. Science 2005, 310, 676–679. [Google Scholar] [CrossRef]
- Witkowski, P.T.; Klempa, B.; Ithete, N.L.; Auste, B.; Mfune, J.K.; Hoveka, J.; Matthee, S.; Preiser, W.; Kruger, D.H. Hantaviruses in Africa. Virus Res. 2014, 187, 34–42. [Google Scholar] [CrossRef]
- Chua, K.B.; Koh, C.L.; Hooi, P.S.; Wee, K.F.; Khong, J.H.; Chua, B.H.; Chan, Y.P.; Lim, M.E.; Lam, S.K. Isolation of Nipah virus from Malaysian Island flying-foxes. Microbes Infect. 2002, 4, 145–151. [Google Scholar] [CrossRef]
- Drexler, J.F.; Corman, V.M.; Gloza-Rausch, F.; Seebens, A.; Annan, A.; Ipsen, A.; Kruppa, T.; Muller, M.A.; Kalko, E.K.; Adu-Sarkodie, Y.; et al. Henipavirus RNA in African bats. PLoS ONE 2009, 4, e6367. [Google Scholar] [CrossRef] [Green Version]
- Leroy, E.M.; Kumulungui, B.; Pourrut, X.; Rouquet, P.; Hassanin, A.; Yaba, P.; Delicat, A.; Paweska, J.T.; Gonzalez, J.P.; Swanepoel, R. Fruit bats as reservoirs of Ebola virus. Nature 2005, 438, 575–576. [Google Scholar] [CrossRef] [PubMed]
- Clayton, B.A.; Wang, L.F.; Marsh, G.A. Henipaviruses: An updated review focusing on the pteropid reservoir and features of transmission. Zoonoses Public Health 2013, 60, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Wacharapluesadee, S.; Lumlertdacha, B.; Boongird, K.; Wanghongsa, S.; Chanhome, L.; Rollin, P.; Stockton, P.; Rupprecht, C.E.; Ksiazek, T.G.; Hemachudha, T. Bat Nipah virus, Thailand. Emerg. Infect. Dis. 2005, 11, 1949–1951. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Yang, L.; Yang, F.; Ren, X.; Jiang, J.; Dong, J.; Sun, L.; Zhu, Y.; Zhou, H.; Jin, Q. Novel Henipa-like virus, Mojiang Paramyxovirus, in rats, China, 2012. Emerg. Infect. Dis. 2014, 20, 1064–1066. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kim, K.; Kim, J.; No, J.S.; Park, K.; Budhathoki, S.; Lee, S.H.; Lee, J.; Cho, S.H.; Cho, S.; et al. Discovery and Genetic Characterization of Novel Paramyxoviruses Related to the Genus Henipavirus in Crocidura Species in the Republic of Korea. Viruses 2021, 13, 2020. [Google Scholar] [CrossRef]
- Field, H.; Young, P.; Yob, J.M.; Mills, J.; Hall, L.; Mackenzie, J. The natural history of Hendra and Nipah viruses. Microbes Infect. 2001, 3, 307–314. [Google Scholar] [CrossRef]
- Hyatt, A.D.; Daszak, P.; Cunningham, A.A.; Field, H.; Gould, A.R. Henipaviruses: Gaps in the Knowledge of Emergence. EcoHealth 2004, 1, 25–38. [Google Scholar] [CrossRef]
- Field, H.; Schaaf, K.; Kung, N.; Simon, C.; Waltisbuhl, D.; Hobert, H.; Moore, F.; Middleton, D.; Crook, A.; Smith, G.; et al. Hendra virus outbreak with novel clinical features, Australia. Emerg. Infect. Dis. 2010, 16, 338–340. [Google Scholar] [CrossRef]
- Murray, K.; Selleck, P.; Hooper, P.; Hyatt, A.; Gould, A.; Gleeson, L.; Westbury, H.; Hiley, L.; Selvey, L.; Rodwell, B.; et al. A morbillivirus that caused fatal disease in horses and humans. Science 1995, 268, 94–97. [Google Scholar] [CrossRef]
- Douglas, I. Equine Morbillivirus: The Search for the Agent; Australian Association of Cattle Veterinarians: Sydney, Australia, 1998. [Google Scholar]
- Young, P.; Halpin, K.; Field, H.; Mackenzie, J.; Asche, V. Natural hendra virus infection in flying-foxes. Recent Adv. Microbiol. 1997. [Google Scholar]
- Westbury, H.A.; Hooper, P.T.; Selleck, P.W.; Murray, P.K. Equine morbillivirus pneumonia: Susceptibility of laboratory animals to the virus. Aust. Vet. J. 1995, 72, 278–279. [Google Scholar] [CrossRef] [PubMed]
- Pavri, K.M.; Singh, K.R.; Hollinger, F.B. Isolation of a new parainfluenza virus from a frugivorous bat, Rousettus leschenaulti, collected at Poona, India. Am. J. Trop. Med. Hyg. 1971, 20, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Zeller, H.G.; Karabatsos, N.; Calisher, C.H.; Digoutte, J.P.; Murphy, F.A.; Shope, R.E. Electron microscopy and antigenic studies of uncharacterized viruses. I. Evidence suggesting the placement of viruses in families Arenaviridae, Paramyxoviridae, or Poxviridae. Arch. Virol. 1989, 108, 191–209. [Google Scholar] [CrossRef] [PubMed]
- Philbey, A.W.; Kirkland, P.D.; Ross, A.D.; Davis, R.J.; Gleeson, A.B.; Love, R.J.; Daniels, P.W.; Gould, A.R.; Hyatt, A.D. An apparently new virus (family Paramyxoviridae) infectious for pigs, humans, and fruit bats. Emerg. Infect. Dis. 1998, 4, 269–271. [Google Scholar] [CrossRef] [Green Version]
- Field, H. The Ecology of Hendra Virus and Australian Bat Lyssavirus. Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 2004. [Google Scholar]
- Centers for Disease Control and Prevention. Outbreak of Hendra-like virus—Malaysia and Singapore, 1998–1999. Morb. Mortal. Wkly. Rep. 1999, 48, 265–269. [Google Scholar]
- Hsu, V.P.; Hossain, M.J.; Parashar, U.D.; Ali, M.M.; Ksiazek, T.G.; Kuzmin, I.; Niezgoda, M.; Rupprecht, C.; Bresee, J.; Breiman, R.F. Nipah virus encephalitis reemergence, Bangladesh. Emerg. Infect. Dis. 2004, 10, 2082–2087. [Google Scholar] [CrossRef]
- Mire, C.E.; Satterfield, B.A.; Geisbert, J.B.; Agans, K.N.; Borisevich, V.; Yan, L.; Chan, Y.P.; Cross, R.W.; Fenton, K.A.; Broder, C.C.; et al. Pathogenic Differences between Nipah Virus Bangladesh and Malaysia Strains in Primates: Implications for Antibody Therapy. Sci. Rep. 2016, 6, 30916. [Google Scholar] [CrossRef]
- Harcourt, B.H.; Lowe, L.; Tamin, A.; Liu, X.; Bankamp, B.; Bowden, N.; Rollin, P.E.; Comer, J.A.; Ksiazek, T.G.; Hossain, M.J.; et al. Genetic characterization of Nipah virus, Bangladesh, 2004. Emerg. Infect. Dis. 2005, 11, 1594–1597. [Google Scholar] [CrossRef]
- Chan, Y.P.; Chua, K.B.; Koh, C.L.; Lim, M.E.; Lam, S.K. Complete nucleotide sequences of Nipah virus isolates from Malaysia. J. Gen. Virol. 2001, 82, 2151–2155. [Google Scholar] [CrossRef]
- AbuBakar, S.; Chang, L.Y.; Ali, A.R.; Sharifah, S.H.; Yusoff, K.; Zamrod, Z. Isolation and molecular identification of Nipah virus from pigs. Emerg. Infect. Dis. 2004, 10, 2228–2230. [Google Scholar] [CrossRef]
- Chua, K.B.; Bellini, W.J.; Rota, P.A.; Harcourt, B.H.; Tamin, A.; Lam, S.K.; Ksiazek, T.G.; Rollin, P.E.; Zaki, S.R.; Shieh, W.; et al. Nipah virus: A recently emergent deadly paramyxovirus. Science 2000, 288, 1432–1435. [Google Scholar] [CrossRef] [PubMed]
- Plowright, R.K.; Becker, D.J.; Crowley, D.E.; Washburne, A.D.; Huang, T.; Nameer, P.O.; Gurley, E.S.; Han, B.A. Prioritizing surveillance of Nipah virus in India. PLoS Negl. Trop. Dis. 2019, 13, e0007393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowron, K.; Bauza-Kaszewska, J.; Grudlewska-Buda, K.; Wiktorczyk-Kapischke, N.; Zacharski, M.; Bernaciak, Z.; Gospodarek-Komkowska, E. Nipah Virus-Another Threat From the World of Zoonotic Viruses. Front. Microbiol. 2021, 12, 811157. [Google Scholar] [CrossRef] [PubMed]
- Sahay, R.R.; Yadav, P.D.; Gupta, N.; Shete, A.M.; Radhakrishnan, C.; Mohan, G.; Menon, N.; Bhatnagar, T.; Suma, K.; Kadam, A.V.; et al. Experiential learnings from the Nipah virus outbreaks in Kerala towards containment of infectious public health emergencies in India. Epidemiol. Infect. 2020, 148, e90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpin, K.; Young, P.L.; Field, H.; Mackenzie, J. Isolation of Hendra virus from pteropid bats: A natural reservoir of Hendra virus. J. Gen. Virol. 2000, 81, 1927–1932. [Google Scholar] [CrossRef]
- Halpin, K.; Hyatt, A.D.; Fogarty, R.; Middleton, D.; Bingham, J.; Epstein, J.H.; Rahman, S.A.; Hughes, T.; Smith, C.; Field, H.E.; et al. Pteropid bats are confirmed as the reservoir hosts of henipaviruses: A comprehensive experimental study of virus transmission. Am. J. Trop. Med. Hyg. 2011, 85, 946–951. [Google Scholar] [CrossRef] [Green Version]
- Epstein, J.H.; Anthony, S.J.; Islam, A.; Kilpatrick, A.M.; Ali Khan, S.; Balkey, M.D.; Ross, N.; Smith, I.; Zambrana-Torrelio, C.; Tao, Y.; et al. Nipah virus dynamics in bats and implications for spillover to humans. Proc. Natl. Acad. Sci. USA 2020, 117, 29190–29201. [Google Scholar] [CrossRef]
- Bates, P.; Harrison, D. Sub-order MEGACHIROPTERA: Family Pteropodidae: Old World fruit bats. In Bats of the Indian Subcontinent; Harrison Zoological Museum: Kent, UK, 1997; pp. 13–15. [Google Scholar]
- Peel, A.J.; Pulliam, J.R.; Luis, A.D.; Plowright, R.K.; O’Shea, T.J.; Hayman, D.T.; Wood, J.L.; Webb, C.T.; Restif, O. The effect of seasonal birth pulses on pathogen persistence in wild mammal populations. Proc. Biol. Sci. 2014, 281, 20132962. [Google Scholar] [CrossRef] [Green Version]
- Plowright, R.K.; Eby, P.; Hudson, P.J.; Smith, I.L.; Westcott, D.; Bryden, W.L.; Middleton, D.; Reid, P.A.; McFarlane, R.A.; Martin, G.; et al. Ecological dynamics of emerging bat virus spillover. Proc. Biol. Sci. 2015, 282, 20142124. [Google Scholar] [CrossRef] [Green Version]
- Gortazar, C.; Reperant, L.A.; Kuiken, T.; de la Fuente, J.; Boadella, M.; Martinez-Lopez, B.; Ruiz-Fons, F.; Estrada-Pena, A.; Drosten, C.; Medley, G.; et al. Crossing the interspecies barrier: Opening the door to zoonotic pathogens. PLoS Pathog. 2014, 10, e1004129. [Google Scholar] [CrossRef] [Green Version]
- Elena, S.F.; Sanjuan, R. Adaptive value of high mutation rates of RNA viruses: Separating causes from consequences. J. Virol 2005, 79, 11555–11558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, D.F.; Rocha, R.; Lopez-Baucells, A.; Farneda, F.Z.; Carreiras, J.M.B.; Palmeirim, J.M.; Meyer, C.F.J. Season-modulated responses of Neotropical bats to forest fragmentation. Ecol. Evol. 2017, 7, 4059–4071. [Google Scholar] [CrossRef] [PubMed]
- Ancillotto, L.; Santini, L.; Ranc, N.; Maiorano, L.; Russo, D. Extraordinary range expansion in a common bat: The potential roles of climate change and urbanisation. Naturwissenschaften 2016, 103, 15. [Google Scholar] [CrossRef] [PubMed]
- Edenborough, K.M.; Mu, A.; Muhldorfer, K.; Lechner, J.; Lander, A.; Bokelmann, M.; Couacy-Hymann, E.; Radonic, A.; Kurth, A. Microbiomes in the insectivorous bat species Mops condylurus rapidly converge in captivity. PLoS ONE 2020, 15, e0223629. [Google Scholar] [CrossRef] [Green Version]
- Mari Saez, A.; Weiss, S.; Nowak, K.; Lapeyre, V.; Zimmermann, F.; Dux, A.; Kuhl, H.S.; Kaba, M.; Regnaut, S.; Merkel, K.; et al. Investigating the zoonotic origin of the West African Ebola epidemic. EMBO Mol. Med. 2015, 7, 17–23. [Google Scholar] [CrossRef]
- Nahar, N.; Paul, R.C.; Sultana, R.; Gurley, E.S.; Garcia, F.; Abedin, J.; Sumon, S.A.; Banik, K.C.; Asaduzzaman, M.; Rimi, N.A.; et al. Raw Sap Consumption Habits and Its Association with Knowledge of Nipah Virus in Two Endemic Districts in Bangladesh. PLoS ONE 2015, 10, e0142292. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.S.; Hossain, J.; Gurley, E.S.; Nahar, N.; Sultana, R.; Luby, S.P. Use of infrared camera to understand bats’ access to date palm sap: Implications for preventing Nipah virus transmission. Ecohealth 2010, 7, 517–525. [Google Scholar] [CrossRef]
- Fogarty, R.; Halpin, K.; Hyatt, A.D.; Daszak, P.; Mungall, B.A. Henipavirus susceptibility to environmental variables. Virus Res. 2008, 132, 140–144. [Google Scholar] [CrossRef] [Green Version]
- Luby, S.P.; Hossain, M.J.; Gurley, E.S.; Ahmed, B.N.; Banu, S.; Khan, S.U.; Homaira, N.; Rota, P.A.; Rollin, P.E.; Comer, J.A.; et al. Recurrent zoonotic transmission of Nipah virus into humans, Bangladesh, 2001–2007. Emerg. Infect. Dis. 2009, 15, 1229–1235. [Google Scholar] [CrossRef]
- Luby, S.P.; Rahman, M.; Hossain, M.J.; Blum, L.S.; Husain, M.M.; Gurley, E.; Khan, R.; Ahmed, B.N.; Rahman, S.; Nahar, N.; et al. Foodborne transmission of Nipah virus, Bangladesh. Emerg. Infect. Dis. 2006, 12, 1888–1894. [Google Scholar] [CrossRef]
- Islam, M.S.; Sazzad, H.M.; Satter, S.M.; Sultana, S.; Hossain, M.J.; Hasan, M.; Rahman, M.; Campbell, S.; Cannon, D.L.; Ströher, U.; et al. Nipah Virus Transmission from Bats to Humans Associated with Drinking Traditional Liquor Made from Date Palm Sap, Bangladesh, 2011–2014. Emerg. Infect. Dis. 2016, 22, 664–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurley, E.S.; Hegde, S.T.; Hossain, K.; Sazzad, H.M.S.; Hossain, M.J.; Rahman, M.; Sharker, M.A.Y.; Salje, H.; Islam, M.S.; Epstein, J.H.; et al. Convergence of Humans, Bats, Trees, and Culture in Nipah Virus Transmission, Bangladesh. Emerg. Infect. Dis. 2017, 23, 1446–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simons, R.R.L.; Gale, P.; Horigan, V.; Snary, E.L.; Breed, A.C. Potential for introduction of bat-borne zoonotic viruses into the EU: A review. Viruses 2014, 6, 2084–2121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A.; Hossain, M.J.; Sultana, S.; Homaira, N.; Khan, S.U.; Rahman, M.; Gurley, E.S.; Rollin, P.E.; Lo, M.K.; Comer, J.A.; et al. Date palm sap linked to Nipah virus outbreak in Bangladesh, 2008. Vector Borne Zoonotic Dis. 2012, 12, 65–72. [Google Scholar] [CrossRef]
- de Wit, E.; Munster, V.J. Animal models of disease shed light on Nipah virus pathogenesis and transmission. J. Pathol. 2015, 235, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Marsh, G.A.; Haining, J.; Hancock, T.J.; Robinson, R.; Foord, A.J.; Barr, J.A.; Riddell, S.; Heine, H.G.; White, J.R.; Crameri, G.; et al. Experimental infection of horses with Hendra virus/Australia/horse/2008/Redlands. Emerg. Infect. Dis. 2011, 17, 2232–2238. [Google Scholar] [CrossRef]
- Gurley, E.S.; Montgomery, J.M.; Hossain, M.J.; Bell, M.; Azad, A.K.; Islam, M.R.; Molla, M.A.; Carroll, D.S.; Ksiazek, T.G.; Rota, P.A.; et al. Person-to-person transmission of Nipah virus in a Bangladeshi community. Emerg. Infect. Dis. 2007, 13, 1031–1037. [Google Scholar] [CrossRef]
- Hadley, M.B.; Blum, L.S.; Mujaddid, S.; Parveen, S.; Nuremowla, S.; Haque, M.E.; Ullah, M. Why Bangladeshi nurses avoid ‘nursing’: Social and structural factors on hospital wards in Bangladesh. Soc. Sci. Med. 2007, 64, 1166–1177. [Google Scholar] [CrossRef]
- Chakraborty, A.; Sazzad, H.M.; Hossain, M.J.; Islam, M.S.; Parveen, S.; Husain, M.; Banu, S.S.; Podder, G.; Afroj, S.; Rollin, P.E.; et al. Evolving epidemiology of Nipah virus infection in Bangladesh: Evidence from outbreaks during 2010–2011. Epidemiol. Infect. 2016, 144, 371–380. [Google Scholar] [CrossRef] [Green Version]
- de Wit, E.; Bushmaker, T.; Scott, D.; Feldmann, H.; Munster, V.J. Nipah virus transmission in a hamster model. PLoS Negl. Trop. Dis. 2011, 5, e1432. [Google Scholar] [CrossRef] [Green Version]
- Weatherman, S.; Feldmann, H.; de Wit, E. Transmission of henipaviruses. Curr. Opin. Virol. 2018, 28, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Dimkic, I.; Fira, D.; Janakiev, T.; Kabic, J.; Stupar, M.; Nenadic, M.; Unkovic, N.; Grbic, M.L. The microbiome of bat guano: For what is this knowledge important? Appl. Microbiol. Biotechnol. 2021, 105, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Kudagammana, H.; Thevanesam, V.; Chu, D.K.W.; Eriyagama, N.B.; Peiris, J.S.M.; Noordeen, F. Coronaviruses in guano from Pteropus medius bats in Peradeniya, Sri Lanka. Transbound. Emerg. Dis. 2018, 65, 1122–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wacharapluesadee, S.; Sintunawa, C.; Kaewpom, T.; Khongnomnan, K.; Olival, K.J.; Epstein, J.H.; Rodpan, A.; Sangsri, P.; Intarut, N.; Chindamporn, A.; et al. Group C betacoronavirus in bat guano fertilizer, Thailand. Emerg. Infect. Dis. 2013, 19, 1349–1351. [Google Scholar] [CrossRef] [Green Version]
- Bowden, T.A.; Aricescu, A.R.; Gilbert, R.J.; Grimes, J.M.; Jones, E.Y.; Stuart, D.I. Structural basis of Nipah and Hendra virus attachment to their cell-surface receptor ephrin-B2. Nat. Struct. Mol. Biol. 2008, 15, 567–572. [Google Scholar] [CrossRef]
- Xu, K.; Rajashankar, K.R.; Chan, Y.P.; Himanen, J.P.; Broder, C.C.; Nikolov, D.B. Host cell recognition by the henipaviruses: Crystal structures of the Nipah G attachment glycoprotein and its complex with ephrin-B3. Proc. Natl. Acad. Sci. USA 2008, 105, 9953–9958. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.; Ataman, Z.A. Modes of paramyxovirus fusion: A Henipavirus perspective. Trends Microbiol. 2011, 19, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Maar, D.; Harmon, B.; Chu, D.; Schulz, B.; Aguilar, H.C.; Lee, B.; Negrete, O.A. Cysteines in the stalk of the nipah virus G glycoprotein are located in a distinct subdomain critical for fusion activation. J. Virol. 2012, 86, 6632–6642. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Amaya, M.; Addetia, A.; Dang, H.V.; Reggiano, G.; Yan, L.; Hickey, A.C.; DiMaio, F.; Broder, C.C.; Veesler, D. Architecture and antigenicity of the Nipah virus attachment glycoprotein. Science 2022, 375, 1373–1378. [Google Scholar] [CrossRef]
- Bowden, T.A.; Crispin, M.; Jones, E.Y.; Stuart, D.I. Shared paramyxoviral glycoprotein architecture is adapted for diverse attachment strategies. Biochem. Soc. Trans. 2010, 38, 1349–1355. [Google Scholar] [CrossRef] [Green Version]
- Negrete, O.A.; Levroney, E.L.; Aguilar, H.C.; Bertolotti-Ciarlet, A.; Nazarian, R.; Tajyar, S.; Lee, B. EphrinB2 is the entry receptor for Nipah virus, an emergent deadly paramyxovirus. Nature 2005, 436, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Pryce, R.; Azarm, K.; Rissanen, I.; Harlos, K.; Bowden, T.A.; Lee, B. A key region of molecular specificity orchestrates unique ephrin-B1 utilization by Cedar virus. Life Sci. Alliance 2020, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laing, E.D.; Navaratnarajah, C.K.; Da Silva, S.C.; Petzing, S.R.; Xu, Y.; Sterling, S.L.; Marsh, G.A.; Wang, L.F.; Amaya, M.; Nikolov, D.B.; et al. Structural and functional analyses reveal promiscuous and species specific use of ephrin receptors by Cedar virus. Proc. Natl. Acad. Sci. USA 2019, 116, 20707–20715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erbar, S.; Diederich, S.; Maisner, A. Selective receptor expression restricts Nipah virus infection of endothelial cells. Virol. J. 2008, 5, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maisner, A.; Neufeld, J.; Weingartl, H. Organ- and endotheliotropism of Nipah virus infections in vivo and in vitro. Thromb. Haemost. 2009, 102, 1014–1023. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, C.; Pohl, C.; Szecsi, J.; Trajkovic-Bodennec, S.; Devergnas, S.; Raoul, H.; Cosset, F.L.; Gerlier, D.; Wild, T.F.; Horvat, B. Nipah virus uses leukocytes for efficient dissemination within a host. J. Virol. 2011, 85, 7863–7871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gale, N.W.; Baluk, P.; Pan, L.; Kwan, M.; Holash, J.; DeChiara, T.M.; McDonald, D.M.; Yancopoulos, G.D. Ephrin-B2 selectively marks arterial vessels and neovascularization sites in the adult, with expression in both endothelial and smooth-muscle cells. Dev. Biol. 2001, 230, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Baseler, L.; de Wit, E.; Scott, D.P.; Munster, V.J.; Feldmann, H. Syrian hamsters (Mesocricetus auratus) oronasally inoculated with a Nipah virus isolate from Bangladesh or Malaysia develop similar respiratory tract lesions. Vet. Pathol. 2015, 52, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Liebl, D.J.; Morris, C.J.; Henkemeyer, M.; Parada, L.F. mRNA expression of ephrins and Eph receptor tyrosine kinases in the neonatal and adult mouse central nervous system. J. Neurosci. Res. 2003, 71, 7–22. [Google Scholar] [CrossRef]
- Benson, M.D.; Romero, M.I.; Lush, M.E.; Lu, Q.R.; Henkemeyer, M.; Parada, L.F. Ephrin-B3 is a myelin-based inhibitor of neurite outgrowth. Proc. Natl. Acad. Sci. USA 2005, 102, 10694–10699. [Google Scholar] [CrossRef] [Green Version]
- Negrete, O.A.; Chu, D.; Aguilar, H.C.; Lee, B. Single amino acid changes in the Nipah and Hendra virus attachment glycoproteins distinguish ephrinB2 from ephrinB3 usage. J. Virol. 2007, 81, 10804–10814. [Google Scholar] [CrossRef] [Green Version]
- Bossart, K.N.; Geisbert, T.W.; Feldmann, H.; Zhu, Z.; Feldmann, F.; Geisbert, J.B.; Yan, L.; Feng, Y.R.; Brining, D.; Scott, D.; et al. A neutralizing human monoclonal antibody protects african green monkeys from hendra virus challenge. Sci. Transl. Med. 2011, 3, 105ra103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughson, F.M. Enveloped viruses: A common mode of membrane fusion? Curr. Biol. 1997, 7, R565–R569. [Google Scholar] [CrossRef] [Green Version]
- Chua, K.B.; Goh, K.J.; Wong, K.T.; Kamarulzaman, A.; Tan, P.S.; Ksiazek, T.G.; Zaki, S.R.; Paul, G.; Lam, S.K.; Tan, C.T. Fatal encephalitis due to Nipah virus among pig-farmers in Malaysia. Lancet 1999, 354, 1257–1259. [Google Scholar] [CrossRef]
- Hafner, C.; Becker, B.; Landthaler, M.; Vogt, T. Expression profile of Eph receptors and ephrin ligands in human skin and downregulation of EphA1 in nonmelanoma skin cancer. Mod. Pathol. 2006, 19, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Bossart, K.N.; Mungall, B.A.; Crameri, G.; Wang, L.F.; Eaton, B.T.; Broder, C.C. Inhibition of Henipavirus fusion and infection by heptad-derived peptides of the Nipah virus fusion glycoprotein. Virol. J. 2005, 2, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu, C.; Guillaume, V.; Sabine, A.; Ong, K.C.; Wong, K.T.; Legras-Lachuer, C.; Horvat, B. Lethal Nipah virus infection induces rapid overexpression of CXCL10. PLoS ONE 2012, 7, e32157. [Google Scholar] [CrossRef] [Green Version]
- Valbuena, G.; Halliday, H.; Borisevich, V.; Goez, Y.; Rockx, B. A human lung xenograft mouse model of Nipah virus infection. PLoS Pathog. 2014, 10, e1004063. [Google Scholar] [CrossRef]
- Escaffre, O.; Borisevich, V.; Carmical, J.R.; Prusak, D.; Prescott, J.; Feldmann, H.; Rockx, B. Henipavirus pathogenesis in human respiratory epithelial cells. J. Virol. 2013, 87, 3284–3294. [Google Scholar] [CrossRef] [Green Version]
- Basler, C.F. Nipah and hendra virus interactions with the innate immune system. Curr. Top. Microbiol. Immunol. 2012, 359, 123–152. [Google Scholar] [CrossRef]
- Yoneda, M.; Guillaume, V.; Sato, H.; Fujita, K.; Georges-Courbot, M.C.; Ikeda, F.; Omi, M.; Muto-Terao, Y.; Wild, T.F.; Kai, C. The nonstructural proteins of Nipah virus play a key role in pathogenicity in experimentally infected animals. PLoS ONE 2010, 5, e12709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu, C.; Guillaume, V.; Volchkova, V.A.; Pohl, C.; Jacquot, F.; Looi, R.Y.; Wong, K.T.; Legras-Lachuer, C.; Volchkov, V.E.; Lachuer, J.; et al. Nonstructural Nipah virus C protein regulates both the early host proinflammatory response and viral virulence. J. Virol. 2012, 86, 10766–10775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolakofsky, D.; Roux, L.; Garcin, D.; Ruigrok, R.W.H. Paramyxovirus mRNA editing, the “rule of six” and error catastrophe: A hypothesis. J. Gen. Virol. 2005, 86, 1869–1877. [Google Scholar] [CrossRef]
- Lo, M.K.; Harcourt, B.H.; Mungall, B.A.; Tamin, A.; Peeples, M.E.; Bellini, W.J.; Rota, P.A. Determination of the henipavirus phosphoprotein gene mRNA editing frequencies and detection of the C, V and W proteins of Nipah virus in virus-infected cells. J. Gen. Virol. 2009, 90, 398–404. [Google Scholar] [CrossRef]
- Ciancanelli, M.J.; Volchkova, V.A.; Shaw, M.L.; Volchkov, V.E.; Basler, C.F. Nipah virus sequesters inactive STAT1 in the nucleus via a P gene-encoded mechanism. J. Virol. 2009, 83, 7828–7841. [Google Scholar] [CrossRef] [Green Version]
- Shaw, M.L.; Garcia-Sastre, A.; Palese, P.; Basler, C.F. Nipah virus V and W proteins have a common STAT1-binding domain yet inhibit STAT1 activation from the cytoplasmic and nuclear compartments, respectively. J. Virol. 2004, 78, 5633–5641. [Google Scholar] [CrossRef] [Green Version]
- Horie, R.; Yoneda, M.; Uchida, S.; Sato, H.; Kai, C. Region of Nipah virus C protein responsible for shuttling between the cytoplasm and nucleus. Virology 2016, 497, 294–304. [Google Scholar] [CrossRef]
- Lo, M.K.; Peeples, M.E.; Bellini, W.J.; Nichol, S.T.; Rota, P.A.; Spiropoulou, C.F. Distinct and overlapping roles of Nipah virus P gene products in modulating the human endothelial cell antiviral response. PLoS ONE 2012, 7, e47790. [Google Scholar] [CrossRef] [Green Version]
- Lo, M.K.; Sogaard, T.M.; Karlin, D.G. Evolution and structural organization of the C proteins of paramyxovirinae. PLoS ONE 2014, 9, e90003. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Kitagawa, Y.; Zhou, M.; Itoh, M.; Gotoh, B. An anti-interferon activity shared by paramyxovirus C proteins: Inhibition of Toll-like receptor 7/9-dependent alpha interferon induction. FEBS Lett. 2014, 588, 28–34. [Google Scholar] [CrossRef]
- Shaw, M.L.; Cardenas, W.B.; Zamarin, D.; Palese, P.; Basler, C.F. Nuclear localization of the Nipah virus W protein allows for inhibition of both virus- and toll-like receptor 3-triggered signaling pathways. J. Virol. 2005, 79, 6078–6088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, J.J.; Parisien, J.P.; Horvath, C.M. Nipah virus V protein evades alpha and gamma interferons by preventing STAT1 and STAT2 activation and nuclear accumulation. J. Virol. 2002, 76, 11476–11483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Childs, K.; Randall, R.; Goodbourn, S. Paramyxovirus V proteins interact with the RNA Helicase LGP2 to inhibit RIG-I-dependent interferon induction. J. Virol. 2012, 86, 3411–3421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Childs, K.; Stock, N.; Ross, C.; Andrejeva, J.; Hilton, L.; Skinner, M.; Randall, R.; Goodbourn, S. mda-5, but not RIG-I, is a common target for paramyxovirus V proteins. Virology 2007, 359, 190–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, J.J.; Wang, L.F.; Horvath, C.M. Hendra virus V protein inhibits interferon signaling by preventing STAT1 and STAT2 nuclear accumulation. J. Virol. 2003, 77, 11842–11845. [Google Scholar] [CrossRef] [Green Version]
- Lieu, K.G.; Marsh, G.A.; Wang, L.F.; Netter, H.J. The non-pathogenic Henipavirus Cedar paramyxovirus phosphoprotein has a compromised ability to target STAT1 and STAT2. Antivir. Res. 2015, 124, 69–76. [Google Scholar] [CrossRef]
- Schountz, T.; Campbell, C.; Wagner, K.; Rovnak, J.; Martellaro, C.; DeBuysscher, B.L.; Feldmann, H.; Prescott, J. Differential Innate Immune Responses Elicited by Nipah Virus and Cedar Virus Correlate with Disparate In Vivo Pathogenesis in Hamsters. Viruses 2019, 11, 291. [Google Scholar] [CrossRef] [Green Version]
- Bharaj, P.; Wang, Y.E.; Dawes, B.E.; Yun, T.E.; Park, A.; Yen, B.; Basler, C.F.; Freiberg, A.N.; Lee, B.; Rajsbaum, R. The Matrix Protein of Nipah Virus Targets the E3-Ubiquitin Ligase TRIM6 to Inhibit the IKKepsilon Kinase-Mediated Type-I IFN Antiviral Response. PLoS Pathog. 2016, 12, e1005880. [Google Scholar] [CrossRef]
- Sugai, A.; Sato, H.; Takayama, I.; Yoneda, M.; Kai, C. Nipah and Hendra Virus Nucleoproteins Inhibit Nuclear Accumulation of Signal Transducer and Activator of Transcription 1 (STAT1) and STAT2 by Interfering with Their Complex Formation. J. Virol. 2017, 91, e01136-17. [Google Scholar] [CrossRef] [Green Version]
- Ramasundpum, V.; Tan, C.T.; Chua, K.B.; Chong, H.T.; Jin, K.; Chew, N.K.; Tan, S.K.; Tahyaparan, T.; Kunjapan, S.R.; Petharunam, V.; et al. Kinetics of IgM and IgCî seroconversion in Nipah virus infection. Neurol. J. Southeast Asia 2000, 5, 23–28. [Google Scholar]
- Arunkumar, G.; Devadiga, S.; McElroy, A.K.; Prabhu, S.; Sheik, S.; Abdulmajeed, J.; Robin, S.; Sushama, A.; Jayaram, A.; Nittur, S.; et al. Adaptive Immune Responses in Humans During Nipah Virus Acute and Convalescent Phases of Infection. Clin. Infect. Dis. 2019, 69, 1752–1756. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.; Playford, E.G.; McBride, W.J.; McMahon, J.; Warrilow, D. No evidence of prolonged Hendra virus shedding by 2 patients, Australia. Emerg Infect Dis. 2012, 18, 2025–2027. [Google Scholar] [CrossRef] [PubMed]
- Weingartl, H.; Czub, S.; Copps, J.; Berhane, Y.; Middleton, D.; Marszal, P.; Gren, J.; Smith, G.; Ganske, S.; Manning, L.; et al. Invasion of the central nervous system in a porcine host by nipah virus. J. Virol. 2005, 79, 7528–7534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munster, V.J.; Prescott, J.B.; Bushmaker, T.; Long, D.; Rosenke, R.; Thomas, T.; Scott, D.; Fischer, E.R.; Feldmann, H.; de Wit, E. Rapid Nipah virus entry into the central nervous system of hamsters via the olfactory route. Sci Rep. 2012, 2, 736. [Google Scholar] [CrossRef] [PubMed]
- Field, H.E. Hendra virus ecology and transmission. Curr. Opin. Virol. 2016, 16, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Williamson, M.M.; Hooper, P.T.; Selleck, P.W.; Gleeson, L.J.; Daniels, P.W.; Westbury, H.A.; Murray, P.K. Transmission studies of Hendra virus (equine morbillivirus) in fruit bats, horses and cats. Aust. Vet. J. 1998, 76, 813–818. [Google Scholar] [CrossRef] [PubMed]
- McLean, R.K.; Graham, S.P. Vaccine Development for Nipah Virus Infection in Pigs. Front. Vet. Sci. 2019, 6, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.T.; Goh, K.J.; Wong, K.T.; Sarji, S.A.; Chua, K.B.; Chew, N.K.; Murugasu, P.; Loh, Y.L.; Chong, H.T.; Tan, K.S.; et al. Relapsed and late-onset Nipah encephalitis. Ann. Neurol. 2002, 51, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.B. Epidemiology, surveillance and control of Nipah virus infections in Malaysia. Malays. J. Pathol. 2010, 32, 69–73. [Google Scholar]
- Baldock, F.C.; Douglas, I.C.; Halpin, K.; Field, H.; Young, P.L.; Black, P. Epidemiological investigations into the 1994 equine morbillivirus outbreaks in Queensland, Australia. Sing. Vet. J. 1996, 20, 57–61. [Google Scholar]
- Rockx, B.; Winegar, R.; Freiberg, A.N. Recent progress in henipavirus research: Molecular biology, genetic diversity, animal models. Antivir. Res. 2012, 95, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Escaffre, O.; Borisevich, V.; Rockx, B. Pathogenesis of Hendra and Nipah virus infection in humans. J. Infect. Dev. Ctries. 2013, 7, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Weingartl, H.M.; Berhane, Y.; Czub, M. Animal models of henipavirus infection: A review. Vet. J. 2009, 181, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Riera Romo, M.; Perez-Martinez, D.; Castillo Ferrer, C. Innate immunity in vertebrates: An overview. Immunology 2016, 148, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Gould, A.R. Comparison of the deduced matrix and fusion protein sequences of equine morbillivirus with cognate genes of the Paramyxoviridae. Virus Res. 1996, 43, 17–31. [Google Scholar] [CrossRef]
- Middleton, D.J.; Morrissy, C.J.; van der Heide, B.M.; Russell, G.M.; Braun, M.A.; Westbury, H.A.; Halpin, K.; Daniels, P.W. Experimental Nipah virus infection in pteropid bats (Pteropus poliocephalus). J. Comp. Pathol. 2007, 136, 266–272. [Google Scholar] [CrossRef]
- Williamson, M.M.; Hooper, P.T.; Selleck, P.W.; Westbury, H.A.; Slocombe, R.F. Experimental hendra virus infectionin pregnant guinea-pigs and fruit Bats (Pteropus poliocephalus). J. Comp. Pathol. 2000, 122, 201–207. [Google Scholar] [CrossRef]
- Field, H.; de Jong, C.; Melville, D.; Smith, C.; Smith, I.; Broos, A.; Kung, Y.H.; McLaughlin, A.; Zeddeman, A. Hendra virus infection dynamics in Australian fruit bats. PLoS ONE 2011, 6, e28678. [Google Scholar] [CrossRef]
- Plowright, R.K.; Field, H.E.; Smith, C.; Divljan, A.; Palmer, C.; Tabor, G.; Daszak, P.; Foley, J.E. Reproduction and nutritional stress are risk factors for Hendra virus infection in little red flying foxes (Pteropus scapulatus). Proc. Biol. Sci. 2008, 275, 861–869. [Google Scholar] [CrossRef] [Green Version]
- Wynne, J.W.; Wang, L.F. Bats and viruses: Friend or foe? PLoS Pathog. 2013, 9, e1003651. [Google Scholar] [CrossRef]
- Baker, M.L.; Schountz, T.; Wang, L.F. Antiviral immune responses of bats: A review. Zoonoses Public Health 2013, 60, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Brook, C.E.; Dobson, A.P. Bats as ‘special’ reservoirs for emerging zoonotic pathogens. Trends Microbiol. 2015, 23, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Allendorf, S.D.; Cortez, A.; Heinemann, M.B.; Harary, C.M.; Antunes, J.M.; Peres, M.G.; Vicente, A.F.; Sodre, M.M.; da Rosa, A.R.; Megid, J. Rabies virus distribution in tissues and molecular characterization of strains from naturally infected non-hematophagous bats. Virus Res. 2012, 165, 119–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamm, D.D.; Kissling, R.E.; Eidson, M.E. Experimental rabies infection in insectivorous bats. J. Infect. Dis. 1956, 98, 10–14. [Google Scholar] [CrossRef]
- Suu-Ire, R.; Begeman, L.; Banyard, A.C.; Breed, A.C.; Drosten, C.; Eggerbauer, E.; Freuling, C.M.; Gibson, L.; Goharriz, H.; Horton, D.L.; et al. Pathogenesis of bat rabies in a natural reservoir: Comparative susceptibility of the straw-colored fruit bat (Eidolon helvum) to three strains of Lagos bat virus. PLoS Negl. Trop. Dis. 2018, 12, e0006311. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Setien, A.; Loza-Rubio, E.; Salas-Rojas, M.; Brisseau, N.; Cliquet, F.; Pastoret, P.P.; Rojas-Dotor, S.; Tesoro, E.; Kretschmer, R. Salivary excretion of rabies virus by healthy vampire bats. Epidemiol. Infect. 2005, 133, 517–522. [Google Scholar] [CrossRef]
- Holmes, D.J.; Ottinger, M.A. Birds as long-lived animal models for the study of aging. Exp. Gerontol. 2003, 38, 1365–1375. [Google Scholar] [CrossRef]
- Mandl, J.N.; Schneider, C.; Schneider, D.S.; Baker, M.L. Going to Bat(s) for Studies of Disease Tolerance. Front. Immunol. 2018, 9, 2112. [Google Scholar] [CrossRef] [Green Version]
- THOMAS, S.P.; SUTHERS, R.A. The Physiology and Energetics of Bat Flight. J. Exp. Biol. 1972, 57, 317–335. [Google Scholar] [CrossRef]
- Shen, Y.Y.; Liang, L.; Zhu, Z.H.; Zhou, W.P.; Irwin, D.M.; Zhang, Y.P. Adaptive evolution of energy metabolism genes and the origin of flight in bats. Proc. Natl. Acad. Sci. USA 2010, 107, 8666–8671. [Google Scholar] [CrossRef] [Green Version]
- Beck, M.A.; Handy, J.; Levander, O.A. The role of oxidative stress in viral infections. Ann. N. Y. Acad. Sci. 2000, 917, 906–912. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.V.; Bartosch, B.; Isaguliants, M.G. Oxidative Stress in Infection and Consequent Disease. Oxid. Med. Cell. Longev. 2017, 2017, 3496043. [Google Scholar] [CrossRef] [PubMed]
- Botten, J.; Mirowsky, K.; Kusewitt, D.; Bharadwaj, M.; Yee, J.; Ricci, R.; Feddersen, R.M.; Hjelle, B. Experimental infection model for Sin Nombre hantavirus in the deer mouse (Peromyscus maniculatus). Proc. Natl. Acad. Sci. USA 2000, 97, 10578–10583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, A.L.; Okumura, A.; Ferris, M.T.; Green, R.; Feldmann, F.; Kelly, S.M.; Scott, D.P.; Safronetz, D.; Haddock, E.; LaCasse, R.; et al. Host genetic diversity enables Ebola hemorrhagic fever pathogenesis and resistance. Science 2014, 346, 987–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, S.K.; Chakravarty, A.K. Analysis of immunocompetent cells in the bat, Pteropus giganteus: Isolation and scanning electron microscopic characterization. Dev. Comp. Immunol. 1991, 15, 423–430. [Google Scholar] [CrossRef]
- Turmelle, A.S.; Ellison, J.A.; Mendonca, M.T.; McCracken, G.F. Histological assessment of cellular immune response to the phytohemagglutinin skin test in Brazilian free-tailed bats (Tadarida brasiliensis). J. Comp. Physiol. B 2010, 180, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Yong, K.S.M.; Ng, J.H.J.; Her, Z.; Hey, Y.Y.; Tan, S.Y.; Tan, W.W.S.; Irac, S.E.; Liu, M.; Chan, X.Y.; Gunawan, M.; et al. Bat-mouse bone marrow chimera: A novel animal model for dissecting the uniqueness of the bat immune system. Sci. Rep. 2018, 8, 4726. [Google Scholar] [CrossRef] [Green Version]
- Ishii, K.J.; Akira, S. Innate immune recognition of nucleic acids: Beyond toll-like receptors. Int J. Cancer 2005, 117, 517–523. [Google Scholar] [CrossRef]
- Iha, K.; Omatsu, T.; Watanabe, S.; Ueda, N.; Taniguchi, S.; Fujii, H.; Ishii, Y.; Kyuwa, S.; Akashi, H.; Yoshikawa, Y. Molecular cloning and expression analysis of bat toll-like receptors 3, 7 and 9. J. Vet. Med. Sci. 2010, 72, 217–220. [Google Scholar] [CrossRef] [Green Version]
- Cowled, C.; Baker, M.; Tachedjian, M.; Zhou, P.; Bulach, D.; Wang, L.F. Molecular characterisation of Toll-like receptors in the black flying fox Pteropus alecto. Dev. Comp. Immunol. 2011, 35, 7–18. [Google Scholar] [CrossRef]
- Shi, Z.; Cai, Z.; Sanchez, A.; Zhang, T.; Wen, S.; Wang, J.; Yang, J.; Fu, S.; Zhang, D. A Novel Toll-like Receptor That Recognizes Vesicular Stomatitis Virus. J. Biol. Chem. 2011, 286, 4517–4524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, A.; Rapin, N.; Bollinger, T.; Misra, V. Lack of inflammatory gene expression in bats: A unique role for a transcription repressor. Sci. Rep. 2017, 7, 2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papenfuss, A.T.; Baker, M.L.; Feng, Z.P.; Tachedjian, M.; Crameri, G.; Cowled, C.; Ng, J.; Janardhana, V.; Field, H.E.; Wang, L.F. The immune gene repertoire of an important viral reservoir, the Australian black flying fox. BMC Genom. 2012, 13, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowled, C.; Baker, M.L.; Zhou, P.; Tachedjian, M.; Wang, L.F. Molecular characterisation of RIG-I-like helicases in the black flying fox, Pteropus alecto. Dev. Comp. Immunol. 2012, 36, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Habjan, M.; Andersson, I.; Klingstrom, J.; Schumann, M.; Martin, A.; Zimmermann, P.; Wagner, V.; Pichlmair, A.; Schneider, U.; Muhlberger, E.; et al. Processing of genome 5′ termini as a strategy of negative-strand RNA viruses to avoid RIG-I-dependent interferon induction. PLoS ONE 2008, 3, e2032. [Google Scholar] [CrossRef] [PubMed]
- Wagner, N.D.; Liu, H.; Rohrs, H.W.; Amarasinghe, G.K.; Gross, M.L.; Leung, D.W. Nipah Virus V Protein Binding Alters MDA5 Helicase Folding Dynamics. ACS Infect. Dis. 2022, 8, 118–128. [Google Scholar] [CrossRef]
- Woon, A.P.; Boyd, V.; Todd, S.; Smith, I.; Klein, R.; Woodhouse, I.B.; Riddell, S.; Crameri, G.; Bingham, J.; Wang, L.F.; et al. Acute experimental infection of bats and ferrets with Hendra virus: Insights into the early host response of the reservoir host and susceptible model species. PLoS Pathog. 2020, 16, e1008412. [Google Scholar] [CrossRef]
- Randall, R.E.; Goodbourn, S. Interferons and viruses: An interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 2008, 89, 1–47. [Google Scholar] [CrossRef]
- Hardy, M.P.; Owczarek, C.M.; Jermiin, L.S.; Ejdeback, M.; Hertzog, P.J. Characterization of the type I interferon locus and identification of novel genes. Genomics 2004, 84, 331–345. [Google Scholar] [CrossRef]
- Fox, B.A.; Sheppard, P.O.; O’Hara, P.J. The role of genomic data in the discovery, annotation and evolutionary interpretation of the interferon-lambda family. PLoS ONE 2009, 4, e4933. [Google Scholar] [CrossRef]
- Kepler, T.B.; Sample, C.; Hudak, K.; Roach, J.; Haines, A.; Walsh, A.; Ramsburg, E.A. Chiropteran types I and II interferon genes inferred from genome sequencing traces by a statistical gene-family assembler. BMC Genom. 2010, 11, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Cowled, C.; Marsh, G.A.; Shi, Z.; Wang, L.F.; Baker, M.L. Type III IFN receptor expression and functional characterisation in the pteropid bat, Pteropus alecto. PLoS ONE 2011, 6, e25385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, W.E.; Scott, W.D.; Sulkin, S.E. Relative sensitivities of viruses to different species of interferon. J. Virol. 1969, 4, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crameri, G.; Todd, S.; Grimley, S.; McEachern, J.A.; Marsh, G.A.; Smith, C.; Tachedjian, M.; De Jong, C.; Virtue, E.R.; Yu, M.; et al. Establishment, immortalisation and characterisation of pteropid bat cell lines. PLoS ONE 2009, 4, e8266. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Cowled, C.; Todd, S.; Crameri, G.; Virtue, E.R.; Marsh, G.A.; Klein, R.; Shi, Z.; Wang, L.F.; Baker, M.L. Type III IFNs in pteropid bats: Differential expression patterns provide evidence for distinct roles in antiviral immunity. J. Immunol. 2011, 186, 3138–3147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virtue, E.R.; Marsh, G.A.; Baker, M.L.; Wang, L.F. Interferon production and signaling pathways are antagonized during henipavirus infection of fruit bat cell lines. PLoS ONE 2011, 6, e22488. [Google Scholar] [CrossRef] [Green Version]
- Virtue, E.R.; Marsh, G.A.; Wang, L.F. Interferon signaling remains functional during henipavirus infection of human cell lines. J. Virol. 2011, 85, 4031–4034. [Google Scholar] [CrossRef] [Green Version]
- Samuel, C.E. Antiviral actions of interferons. Clin. Microbiol. Rev. 2001, 14, 778–809, table of contents. [Google Scholar] [CrossRef] [Green Version]
- Brzozka, K.; Finke, S.; Conzelmann, K.K. Inhibition of interferon signaling by rabies virus phosphoprotein P: Activation-dependent binding of STAT1 and STAT2. J. Virol. 2006, 80, 2675–2683. [Google Scholar] [CrossRef] [Green Version]
- Fujii, H.; Watanabe, S.; Yamane, D.; Ueda, N.; Iha, K.; Taniguchi, S.; Kato, K.; Tohya, Y.; Kyuwa, S.; Yoshikawa, Y.; et al. Functional analysis of Rousettus aegyptiacus “signal transducer and activator of transcription 1” (STAT1). Dev. Comp. Immunol. 2010, 34, 598–602. [Google Scholar] [CrossRef]
- De La Cruz-Rivera, P.C.; Kanchwala, M.; Liang, H.; Kumar, A.; Wang, L.F.; Xing, C.; Schoggins, J.W. The IFN Response in Bats Displays Distinctive IFN-Stimulated Gene Expression Kinetics with Atypical RNASEL Induction. J. Immunol. 2018, 200, 209–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, S.F. Cytokine mediators of immunity and inflammation. Arch. Surg. 1993, 128, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Iha, K.; Omatsu, T.; Watanabe, S.; Ueda, N.; Taniguchi, S.; Fujii, H.; Ishii, Y.; Kyuwa, S.; Akashi, H.; Yoshikawa, Y. Molecular cloning and sequencing of the cDNAs encoding the bat interleukin (IL)-2, IL-4, IL-6, IL-10, IL-12p40, and tumor necrosis factor-alpha. J. Vet. Med. Sci. 2009, 71, 1691–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cogswell-Hawkinson, A.C.; McGlaughlin, M.E.; Calisher, C.H.; Adams, R.; Schountz, T. Molecular and phylogenetic characterization of cytokine genes from Seba’s short-tailed bat (Carollia perspicillata). Open Immunol. J. 2011, 4, 31–39. [Google Scholar] [CrossRef]
- Zhang, G.; Cowled, C.; Shi, Z.; Huang, Z.; Bishop-Lilly, K.A.; Fang, X.; Wynne, J.W.; Xiong, Z.; Baker, M.L.; Zhao, W. Comparative analysis of bat genomes provides insight into the evolution of flight and immunity. Science 2013, 339, 456–460. [Google Scholar] [CrossRef] [Green Version]
- Kacprzyk, J.; Hughes, G.M.; Palsson-McDermott, E.M.; Quinn, S.R.; Puechmaille, S.J.; O’neill, L.A.; Teeling, E.C. A potent anti-inflammatory response in bat macrophages may be linked to extended longevity and viral tolerance. Acta Chiropterologica 2017, 19, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Pavlovich, S.S.; Lovett, S.P.; Koroleva, G.; Guito, J.C.; Arnold, C.E.; Nagle, E.R.; Kulcsar, K.; Lee, A.; Thibaud-Nissen, F.; Hume, A.J. The Egyptian rousette genome reveals unexpected features of bat antiviral immunity. Cell 2018, 173, 1098–1110.e18. [Google Scholar] [CrossRef] [Green Version]
- Cohen, O.; Weissman, D.; Fauci, A.; Paul, W. Fundamental Immunology; Lippincott-Raven: Philadelphia, PA, USA, 1999. [Google Scholar]
- Butler, J.E.; Wertz, N.; Zhao, Y.; Zhang, S.; Bao, Y.; Bratsch, S.; Kunz, T.H.; Whitaker, J.O., Jr.; Schountz, T. The two suborders of chiropterans have the canonical heavy-chain immunoglobulin (Ig) gene repertoire of eutherian mammals. Dev. Comp. Immunol. 2011, 35, 273–284. [Google Scholar] [CrossRef]
- Zhao, Y.; Cui, H.; Whittington, C.M.; Wei, Z.; Zhang, X.; Zhang, Z.; Yu, L.; Ren, L.; Hu, X.; Zhang, Y. Ornithorhynchus anatinus (platypus) links the evolution of immunoglobulin genes in eutherian mammals and nonmammalian tetrapods. J. Immunol. 2009, 183, 3285–3293. [Google Scholar] [CrossRef] [Green Version]
- Ohta, Y.; Flajnik, M. IgD, like IgM, is a primordial immunoglobulin class perpetuated in most jawed vertebrates. Proc. Natl. Acad. Sci. USA 2006, 103, 10723–10728. [Google Scholar] [CrossRef] [Green Version]
- Bratsch, S.; Wertz, N.; Chaloner, K.; Kunz, T.H.; Butler, J.E. The little brown bat, M. lucifugus, displays a highly diverse VH, DH and JH repertoire but little evidence of somatic hypermutation. Dev. Comp. Immunol. 2011, 35, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.L.; Tachedjian, M.; Wang, L.-F. Immunoglobulin heavy chain diversity in Pteropid bats: Evidence for a diverse and highly specific antigen binding repertoire. Immunogenetics 2010, 62, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Radic, M.; Mackle, J.; Erikson, J.; Mol, C.; Anderson, W.; Weigert, M. Residues that mediate DNA binding of autoimmune antibodies. J. Immunol. 1993, 150, 4966–4977. [Google Scholar] [PubMed]
- Birtalan, S.; Zhang, Y.; Fellouse, F.A.; Shao, L.; Schaefer, G.; Sidhu, S.S. The intrinsic contributions of tyrosine, serine, glycine and arginine to the affinity and specificity of antibodies. J. Mol. Biol. 2008, 377, 1518–1528. [Google Scholar] [CrossRef] [PubMed]
- Sulkin, S.E.; Allen, R.; Sims, R. Studies of arthropod-borne virus infections in Chiroptera. Am. J. Trop. Med. Hyg. 1966, 15, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Peel, A.J.; Baker, K.S.; Hayman, D.T.; Broder, C.C.; Cunningham, A.A.; Fooks, A.R.; Garnier, R.; Wood, J.L.; Restif, O. Support for viral persistence in bats from age-specific serology and models of maternal immunity. Sci. Rep. 2018, 8, 3859. [Google Scholar] [CrossRef]
- Brook, C.E.; Ranaivoson, H.C.; Broder, C.C.; Cunningham, A.A.; Héraud, J.M.; Peel, A.J.; Gibson, L.; Wood, J.L.; Metcalf, C.J.; Dobson, A.P. Disentangling serology to elucidate henipa-and filovirus transmission in Madagascar fruit bats. J. Anim. Ecol. 2019, 88, 1001–1016. [Google Scholar] [CrossRef]
- Chakraborty, A.K.; k Chakravarty, A. Dichotomy of lymphocyte population and cell-mediated immune responses in a fruit bat, Pteropus giganteus. J. Indian Inst. Sci. 1983, 64, 157. [Google Scholar]
- Wellehan, J.F., Jr.; Green, L.G.; Duke, D.G.; Bootorabi, S.; Heard, D.J.; Klein, P.A.; Jacobson, E.R. Detection of specific antibody responses to vaccination in variable flying foxes (Pteropus hypomelanus). Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 379–394. [Google Scholar] [CrossRef]
- Hatten, B.A.; Allen, R.; Sulkin, S.E. Studies on the immune capabilities of Chiroptera: I. Quantitative and qualitative nature of the immune responses in bats to bacteriophage φX174. J. Immunol. 1970, 105, 872–878. [Google Scholar]
- Sohayati, A.; Hassan, L.; Sharifah, S.; Lazarus, K.; Zaini, C.; Epstein, J.; Naim, N.S.; Field, H.; Arshad, S.S.; Aziz, J.A. Evidence for Nipah virus recrudescence and serological patterns of captive Pteropus vampyrus. Epidemiol. Infect. 2011, 139, 1570–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glennon, E.E.; Becker, D.J.; Peel, A.J.; Garnier, R.; Suu-Ire, R.D.; Gibson, L.; Hayman, D.T.S.; Wood, J.L.N.; Cunningham, A.A.; Plowright, R.K.; et al. What is stirring in the reservoir? Modelling mechanisms of henipavirus circulation in fruit bat hosts. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20190021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakravarty, A.; Sarkar, S. Immunofluorescence analysis of immunoglobulin bearing lymphocytes in the Indian fruit bat: Pteropus giganteus. Lymphology 1994, 27, 97–104. [Google Scholar] [PubMed]
- Omatsu, T.; Nishimura, Y.; Bak, E.J.; Ishii, Y.; Tohya, Y.; Kyuwa, S.; Akashi, H.; Yoshikawa, Y. Molecular cloning and sequencing of the cDNA encoding the bat CD4. Vet. Immunol. Immunopathol. 2006, 111, 309–313. [Google Scholar]
- McMurray, D.N.; Thomas, M.E. Cell-mediated immunity in two species of bats. J. Mammal. 1979, 60, 576–581. [Google Scholar] [CrossRef]
- Paul, B.N.; Chakravarty, A.K. Invitro analysis of delayed immune response in a bat, Pteropusgiganteus: Process of con-A mediated activation. Dev. Comp. Immunol. 1986, 10, 55–67. [Google Scholar] [CrossRef]
- Derks, R.A.; Burlingham, W.J. In vitro parameters of donor-antigen-specific tolerance. Curr. Opin. Immunol. 2005, 17, 560–564. [Google Scholar] [CrossRef]
- Schad, J.; Dechmann, D.K.; Voigt, C.C.; Sommer, S. MHC class II DRB diversity, selection pattern and population structure in a neotropical bat species, Noctilio albiventris. Heredity 2011, 107, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Mayer, F.; Brunner, A. Non-neutral evolution of the major histocompatibility complex class II gene DRB 1 in the sac-winged bat Saccopteryx bilineata. Heredity 2007, 99, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Kuzmin, I.V.; Schwarz, T.M.; Ilinykh, P.A.; Jordan, I.; Ksiazek, T.G.; Sachidanandam, R.; Basler, C.F.; Bukreyev, A. Innate immune responses of bat and human cells to filoviruses: Commonalities and distinctions. J. Virol. 2017, 91, e02471-16. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.E.; Schuh, A.J.; Amman, B.R.; Sealy, T.K.; Zaki, S.R.; Nichol, S.T.; Towner, J.S. Experimental Inoculation of Egyptian Rousette Bats (Rousettus aegyptiacus) with Viruses of the Ebolavirus and Marburgvirus Genera. Viruses 2015, 7, 3420–3442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westbury, H.A.; Hooper, P.T.; Brouwer, S.L.; Selleck, P.W. Susceptibility of cats to equine morbillivirus. Aust. Vet. J. 1996, 74, 132–134. [Google Scholar] [CrossRef] [PubMed]
- Hooper, P.T.; Westbury, H.A.; Russell, G.M. The lesions of experimental equine morbillivirus disease in cats and guinea pigs. Vet. Pathol. 1997, 34, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Mungall, B.A.; Middleton, D.; Crameri, G.; Bingham, J.; Halpin, K.; Russell, G.; Green, D.; McEachern, J.; Pritchard, L.I.; Eaton, B.T.; et al. Feline model of acute nipah virus infection and protection with a soluble glycoprotein-based subunit vaccine. J. Virol. 2006, 80, 12293–12302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mungall, B.A.; Middleton, D.; Crameri, G.; Halpin, K.; Bingham, J.; Eaton, B.T.; Broder, C.C. Vertical transmission and fetal replication of Nipah virus in an experimentally infected cat. J. Infect. Dis. 2007, 196, 812–816. [Google Scholar] [CrossRef] [Green Version]
- Hooper, P.; Zaki, S.; Daniels, P.; Middleton, D. Comparative pathology of the diseases caused by Hendra and Nipah viruses. Microbes Infect. 2001, 3, 315–322. [Google Scholar] [CrossRef]
- Halim, S.; Polkinghorne, B.; Bell, G.; van den Berg, D.; Sheppeard, V. Outbreak-related Hendra virus infection in a NSW pet dog. Public Health Res. Pract. 2015, 25, e2541547. [Google Scholar] [CrossRef] [Green Version]
- Kirkland, P.D.; Gabor, M.; Poe, I.; Neale, K.; Chaffey, K.; Finlaison, D.S.; Gu, X.; Hick, P.M.; Read, A.J.; Wright, T.; et al. Hendra Virus Infection in Dog, Australia, 2013. Emerg. Infect. Dis. 2015, 21, 2182–2185. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.N.; Alim, A.N.; Bunning, M.L.; Lee, O.B.; Wagoner, K.D.; Amman, B.R.; Stockton, P.C.; Ksiazek, T.G. Nipah virus infection in dogs, Malaysia, 1999. Emerg. Infect. Dis. 2009, 15, 950–952. [Google Scholar] [CrossRef]
- Parashar, U.D.; Sunn, L.M.; Ong, F.; Mounts, A.W.; Arif, M.T.; Ksiazek, T.G.; Kamaluddin, M.A.; Mustafa, A.N.; Kaur, H.; Ding, L.M.; et al. Case-control study of risk factors for human infection with a new zoonotic paramyxovirus, Nipah virus, during a 1998–1999 outbreak of severe encephalitis in Malaysia. J. Infect. Dis. 2000, 181, 1755–1759. [Google Scholar] [CrossRef]
- Pallister, J.; Middleton, D.; Wang, L.F.; Klein, R.; Haining, J.; Robinson, R.; Yamada, M.; White, J.; Payne, J.; Feng, Y.R.; et al. A recombinant Hendra virus G glycoprotein-based subunit vaccine protects ferrets from lethal Hendra virus challenge. Vaccine 2011, 29, 5623–5630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leon, A.J.; Borisevich, V.; Boroumand, N.; Seymour, R.; Nusbaum, R.; Escaffre, O.; Xu, L.; Kelvin, D.J.; Rockx, B. Host gene expression profiles in ferrets infected with genetically distinct henipavirus strains. PLoS Negl. Trop. Dis. 2018, 12, e0006343. [Google Scholar] [CrossRef] [PubMed]
- Mire, C.E.; Chan, Y.P.; Borisevich, V.; Cross, R.W.; Yan, L.; Agans, K.N.; Dang, H.V.; Veesler, D.; Fenton, K.A.; Geisbert, T.W.; et al. A Cross-Reactive Humanized Monoclonal Antibody Targeting Fusion Glycoprotein Function Protects Ferrets Against Lethal Nipah Virus and Hendra Virus Infection. J. Infect. Dis. 2020, 221 (Suppl. 4), S471–S479. [Google Scholar] [CrossRef] [PubMed]
- Bossart, K.N.; Zhu, Z.; Middleton, D.; Klippel, J.; Crameri, G.; Bingham, J.; McEachern, J.A.; Green, D.; Hancock, T.J.; Chan, Y.P.; et al. A neutralizing human monoclonal antibody protects against lethal disease in a new ferret model of acute nipah virus infection. PLoS Pathog. 2009, 5, e1000642. [Google Scholar] [CrossRef]
- Clayton, B.A.; Middleton, D.; Arkinstall, R.; Frazer, L.; Wang, L.F.; Marsh, G.A. The Nature of Exposure Drives Transmission of Nipah Viruses from Malaysia and Bangladesh in Ferrets. PLoS Negl. Trop. Dis. 2016, 10, e0004775. [Google Scholar] [CrossRef]
- Clayton, B.A.; Middleton, D.; Bergfeld, J.; Haining, J.; Arkinstall, R.; Wang, L.; Marsh, G.A. Transmission routes for nipah virus from Malaysia and Bangladesh. Emerg. Infect. Dis. 2012, 18, 1983–1993. [Google Scholar] [CrossRef]
- Pallister, J.; Middleton, D.; Crameri, G.; Yamada, M.; Klein, R.; Hancock, T.J.; Foord, A.; Shiell, B.; Michalski, W.; Broder, C.C.; et al. Chloroquine administration does not prevent.t Nipah virus infection and disease in ferrets. J. Virol. 2009, 83, 11979–11982. [Google Scholar] [CrossRef] [Green Version]
- Satterfield, B.A.; Borisevich, V.; Foster, S.L.; Rodriguez, S.E.; Cross, R.W.; Fenton, K.A.; Agans, K.N.; Basler, C.F.; Geisbert, T.W.; Mire, C.E. Antagonism of STAT1 by Nipah virus P gene products modulates disease course but not lethal outcome in the ferret model. Sci. Rep. 2019, 9, 16710. [Google Scholar] [CrossRef] [Green Version]
- Pallister, J.A.; Klein, R.; Arkinstall, R.; Haining, J.; Long, F.; White, J.R.; Payne, J.; Feng, Y.R.; Wang, L.F.; Broder, C.C.; et al. Vaccination of ferrets with a recombinant G glycoprotein subunit vaccine provides protection against Nipah virus disease for over 12 months. Virol. J. 2013, 10, 237. [Google Scholar] [CrossRef] [Green Version]
- Hooper, P.T.; Ketterer, P.J.; Hyatt, A.D.; Russell, G.M. Lesions of experimental equine morbillivirus pneumonia in horses. Vet. Pathol. 1997, 34, 312–322. [Google Scholar] [CrossRef]
- Kung, N.; McLaughlin, A.; Taylor, M.; Moloney, B.; Wright, T.; Field, H. Hendra virus and horse owners--risk perception and management. PLoS ONE 2013, 8, e80897. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.; Hodge, A.; Klein, R.; Edwards, N.; Huang, J.A.; Middleton, D.; Watts, S.P. Virus-neutralising antibody responses in horses following vaccination with Equivac(R) HeV: A field study. Aust. Vet. J. 2018, 96, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Ching, P.K.; de los Reyes, V.C.; Sucaldito, M.N.; Tayag, E.; Columna-Vingno, A.B.; Malbas, F.F., Jr.; Bolo, G.C., Jr.; Sejvar, J.J.; Eagles, D.; Playford, G.; et al. Outbreak of henipavirus infection, Philippines, 2014. Emerg. Infect. Dis. 2015, 21, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Rockx, B.; Bossart, K.N.; Feldmann, F.; Geisbert, J.B.; Hickey, A.C.; Brining, D.; Callison, J.; Safronetz, D.; Marzi, A.; Kercher, L.; et al. A novel model of lethal Hendra virus infection in African green monkeys and the effectiveness of ribavirin treatment. J. Virol. 2010, 84, 9831–9839. [Google Scholar] [CrossRef] [Green Version]
- Prasad, A.N.; Woolsey, C.; Geisbert, J.B.; Agans, K.N.; Borisevich, V.; Deer, D.J.; Mire, C.E.; Cross, R.W.; Fenton, K.A.; Broder, C.C.; et al. Resistance of Cynomolgus Monkeys to Nipah and Hendra Virus Disease Is Associated With Cell-Mediated and Humoral Immunity. J. Infect. Dis. 2020, 221 (Suppl. 4), S436–S447. [Google Scholar] [CrossRef]
- Bossart, K.N.; Rockx, B.; Feldmann, F.; Brining, D.; Scott, D.; LaCasse, R.; Geisbert, J.B.; Feng, Y.R.; Chan, Y.P.; Hickey, A.C.; et al. A Hendra virus G glycoprotein subunit vaccine protects African green monkeys from Nipah virus challenge. Sci. Transl. Med. 2012, 4, 146ra107. [Google Scholar] [CrossRef] [Green Version]
- Geisbert, T.W.; Feldmann, H.; Broder, C.C. Animal challenge models of henipavirus infection and pathogenesis. Curr. Top. Microbiol. Immunol. 2012, 359, 153–177. [Google Scholar] [CrossRef]
- Geisbert, J.B.; Borisevich, V.; Prasad, A.N.; Agans, K.N.; Foster, S.L.; Deer, D.J.; Cross, R.W.; Mire, C.E.; Geisbert, T.W.; Fenton, K.A. An Intranasal Exposure Model of Lethal Nipah Virus Infection in African Green Monkeys. J. Infect. Dis. 2020, 221 (Suppl. 4), S414–S418. [Google Scholar] [CrossRef]
- Hammoud, D.A.; Lentz, M.R.; Lara, A.; Bohannon, J.K.; Feuerstein, I.; Huzella, L.; Jahrling, P.B.; Lackemeyer, M.; Laux, J.; Rojas, O.; et al. Aerosol exposure to intermediate size Nipah virus particles induces neurological disease in African green monkeys. PLoS Negl. Trop. Dis. 2018, 12, e0006978. [Google Scholar] [CrossRef]
- Liu, J.; Coffin, K.M.; Johnston, S.C.; Babka, A.M.; Bell, T.M.; Long, S.Y.; Honko, A.N.; Kuhn, J.H.; Zeng, X. Nipah virus persists in the brains of nonhuman primate survivors. JCI Insight 2019, 4, e129629. [Google Scholar] [CrossRef]
- Cong, Y.; Lentz, M.R.; Lara, A.; Alexander, I.; Bartos, C.; Bohannon, J.K.; Hammoud, D.; Huzella, L.; Jahrling, P.B.; Janosko, K.; et al. Loss in lung volume and changes in the immune response demonstrate disease progression in African green monkeys infected by small-particle aerosol and intratracheal exposure to Nipah virus. PLoS Negl. Trop. Dis. 2017, 11, e0005532. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Hammoud, D.A.; Cong, Y.; Huzella, L.M.; Castro, M.A.; Solomon, J.; Laux, J.; Lackemeyer, M.; Bohannon, J.K.; Rojas, O.; et al. The Use of Large-Particle Aerosol Exposure to Nipah Virus to Mimic Human Neurological Disease Manifestations in the African Green Monkey. J. Infect. Dis. 2020, 221 (Suppl. 4), S419–S430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, A.N.; Agans, K.N.; Sivasubramani, S.K.; Geisbert, J.B.; Borisevich, V.; Mire, C.E.; Lawrence, W.S.; Fenton, K.A.; Geisbert, T.W. A Lethal Aerosol Exposure Model of Nipah Virus Strain Bangladesh in African Green Monkeys. J. Infect. Dis. 2020, 221 (Suppl. 4), S431–S435. [Google Scholar] [CrossRef] [PubMed]
- Lara, A.; Cong, Y.; Jahrling, P.B.; Mednikov, M.; Postnikova, E.; Yu, S.; Munster, V.; Holbrook, M.R. Peripheral immune response in the African green monkey model following Nipah-Malaysia virus exposure by intermediate-size particle aerosol. PLoS Negl. Trop. Dis. 2019, 13, e0007454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marianneau, P.; Guillaume, V.; Wong, T.; Badmanathan, M.; Looi, R.Y.; Murri, S.; Loth, P.; Tordo, N.; Wild, F.; Horvat, B.; et al. Experimental infection of squirrel monkeys with nipah virus. Emerg. Infect. Dis. 2010, 16, 507–510. [Google Scholar] [CrossRef]
- Black, P.F.; Cronin, J.P.; Morrissy, C.J.; Westbury, H.A. Serological examination for evidence of infection with Hendra and Nipah viruses in Queensland piggeries. Aust. Vet. J. 2001, 79, 424–426. [Google Scholar] [CrossRef]
- Hayman, D.T.; Wang, L.F.; Barr, J.; Baker, K.S.; Suu-Ire, R.; Broder, C.C.; Cunningham, A.A.; Wood, J.L. Antibodies to henipavirus or henipa-like viruses in domestic pigs in Ghana, West Africa. PLoS ONE 2011, 6, e25256. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Embury-Hyatt, C.; Weingartl, H.M. Experimental inoculation study indicates swine as a potential host for Hendra virus. Vet. Res. 2010, 41, 33. [Google Scholar] [CrossRef] [Green Version]
- Pickering, B.S.; Hardham, J.M.; Smith, G.; Weingartl, E.T.; Dominowski, P.J.; Foss, D.L.; Mwangi, D.; Broder, C.C.; Roth, J.A.; Weingartl, H.M. Protection against henipaviruses in swine requires both, cell-mediated and humoral immune response. Vaccine 2016, 34, 4777–4786. [Google Scholar] [CrossRef]
- Berhane, Y.; Weingartl, H.M.; Lopez, J.; Neufeld, J.; Czub, S.; Embury-Hyatt, C.; Goolia, M.; Copps, J.; Czub, M. Bacterial infections in pigs experimentally infected with Nipah virus. Transbound. Emerg. Dis. 2008, 55, 165–174. [Google Scholar] [CrossRef]
- Stachowiak, B.; Weingartl, H.M. Nipah virus infects specific subsets of porcine peripheral blood mononuclear cells. PLoS ONE 2012, 7, e30855. [Google Scholar] [CrossRef] [PubMed]
- Weingartl, H.M.; Berhane, Y.; Caswell, J.L.; Loosmore, S.; Audonnet, J.C.; Roth, J.A.; Czub, M. Recombinant nipah virus vaccines protect pigs against challenge. J. Virol. 2006, 80, 7929–7938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedrera, M.; Macchi, F.; McLean, R.K.; Franceschi, V.; Thakur, N.; Russo, L.; Medfai, L.; Todd, S.; Tchilian, E.Z.; Audonnet, J.C.; et al. Bovine Herpesvirus-4-Vectored Delivery of Nipah Virus Glycoproteins Enhances T Cell Immunogenicity in Pigs. Vaccines 2020, 8, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuai, L.; Ge, J.; Wen, Z.; Wang, J.; Wang, X.; Bu, Z. Immune responses in mice and pigs after oral vaccination with rabies virus vectored Nipah disease vaccines. Vet. Microbiol. 2020, 241, 108549. [Google Scholar] [CrossRef]
- Kalodimou, G.; Veit, S.; Jany, S.; Kalinke, U.; Broder, C.C.; Sutter, G.; Volz, A. A Soluble Version of Nipah Virus Glycoprotein G Delivered by Vaccinia Virus MVA Activates Specific CD8 and CD4 T Cells in Mice. Viruses 2019, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- Yun, T.; Park, A.; Hill, T.E.; Pernet, O.; Beaty, S.M.; Juelich, T.L.; Smith, J.K.; Zhang, L.; Wang, Y.E.; Vigant, F.; et al. Efficient reverse genetics reveals genetic determinants of budding and fusogenic differences between Nipah and Hendra viruses and enables real-time monitoring of viral spread in small animal models of henipavirus infection. J. Virol. 2015, 89, 1242–1253. [Google Scholar] [CrossRef] [Green Version]
- Dups, J.; Middleton, D.; Yamada, M.; Monaghan, P.; Long, F.; Robinson, R.; Marsh, G.A.; Wang, L.F. A new model for Hendra virus encephalitis in the mouse. PLoS ONE 2012, 7, e40308. [Google Scholar] [CrossRef] [Green Version]
- Dups, J.; Middleton, D.; Long, F.; Arkinstall, R.; Marsh, G.A.; Wang, L.F. Subclinical infection without encephalitis in mice following intranasal exposure to Nipah virus-Malaysia and Nipah virus-Bangladesh. Virol. J. 2014, 11, 102. [Google Scholar] [CrossRef] [Green Version]
- Iampietro, M.; Aurine, N.; Dhondt, K.P.; Dumont, C.; Pelissier, R.; Spanier, J.; Vallve, A.; Raoul, H.; Kalinke, U.; Horvat, B. Control of Nipah Virus Infection in Mice by the Host Adaptors Mitochondrial Antiviral Signaling Protein (MAVS) and Myeloid Differentiation Primary Response 88 (MyD88). J. Infect. Dis. 2020, 221 (Suppl. 4), S401–S406. [Google Scholar] [CrossRef]
- Keshwara, R.; Shiels, T.; Postnikova, E.; Kurup, D.; Wirblich, C.; Johnson, R.F.; Schnell, M.J. Rabies-based vaccine induces potent immune responses against Nipah virus. npj Vaccines 2019, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, R.; Wang, M.; Liu, Y.; Yin, Y.; Zai, X.; Song, X.; Chen, Y.; Xu, J.; Chen, W. Fc-Based Recombinant Henipavirus Vaccines Elicit Broad Neutralizing Antibody Responses in Mice. Viruses 2020, 12, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loomis, R.J.; DiPiazza, A.T.; Falcone, S.; Ruckwardt, T.J.; Morabito, K.M.; Abiona, O.M.; Chang, L.A.; Caringal, R.T.; Presnyak, V.; Narayanan, E.; et al. Chimeric Fusion (F) and Attachment (G) Glycoprotein Antigen Delivery by mRNA as a Candidate Nipah Vaccine. Front. Immunol. 2021, 12, 772864. [Google Scholar] [CrossRef] [PubMed]
- Stroh, E.; Fischer, K.; Schwaiger, T.; Sauerhering, L.; Franzke, K.; Maisner, A.; Groschup, M.H.; Blohm, U.; Diederich, S. Henipavirus-like particles induce a CD8 T cell response in C57BL/6 mice. Vet. Microbiol. 2019, 237, 108405. [Google Scholar] [CrossRef] [PubMed]
- Freiberg, A.N.; Worthy, M.N.; Lee, B.; Holbrook, M.R. Combined chloroquine and ribavirin treatment does not prevent death in a hamster model of Nipah and Hendra virus infection. J. Gen. Virol. 2010, 91, 765–772. [Google Scholar] [CrossRef]
- Guillaume, V.; Wong, K.T.; Looi, R.Y.; Georges-Courbot, M.C.; Barrot, L.; Buckland, R.; Wild, T.F.; Horvat, B. Acute Hendra virus infection: Analysis of the pathogenesis and passive antibody protection in the hamster model. Virology 2009, 387, 459–465. [Google Scholar] [CrossRef] [Green Version]
- Ploquin, A.; Szecsi, J.; Mathieu, C.; Guillaume, V.; Barateau, V.; Ong, K.C.; Wong, K.T.; Cosset, F.L.; Horvat, B.; Salvetti, A. Protection against henipavirus infection by use of recombinant adeno-associated virus-vector vaccines. J. Infect. Dis. 2013, 207, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Rockx, B.; Brining, D.; Kramer, J.; Callison, J.; Ebihara, H.; Mansfield, K.; Feldmann, H. Clinical outcome of henipavirus infection in hamsters is determined by the route and dose of infection. J. Virol. 2011, 85, 7658–7671. [Google Scholar] [CrossRef] [Green Version]
- DeBuysscher, B.L.; de Wit, E.; Munster, V.J.; Scott, D.; Feldmann, H.; Prescott, J. Comparison of the pathogenicity of Nipah virus isolates from Bangladesh and Malaysia in the Syrian hamster. PLoS Negl. Trop. Dis. 2013, 7, e2024. [Google Scholar] [CrossRef] [Green Version]
- Guillaume, V.; Contamin, H.; Loth, P.; Georges-Courbot, M.C.; Lefeuvre, A.; Marianneau, P.; Chua, K.B.; Lam, S.K.; Buckland, R.; Deubel, V.; et al. Nipah virus: Vaccination and passive protection studies in a hamster model. J. Virol. 2004, 78, 834–840. [Google Scholar] [CrossRef] [Green Version]
- Guillaume, V.; Contamin, H.; Loth, P.; Grosjean, I.; Courbot, M.C.; Deubel, V.; Buckland, R.; Wild, T.F. Antibody prophylaxis and therapy against Nipah virus infection in hamsters. J. Virol. 2006, 80, 1972–1978. [Google Scholar] [CrossRef] [Green Version]
- Escaffre, O.; Hill, T.; Ikegami, T.; Juelich, T.L.; Smith, J.K.; Zhang, L.; Perez, D.E.; Atkins, C.; Park, A.; Lawrence, W.S.; et al. Experimental Infection of Syrian Hamsters With Aerosolized Nipah Virus. J. Infect. Dis. 2018, 218, 1602–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georges-Courbot, M.C.; Contamin, H.; Faure, C.; Loth, P.; Baize, S.; Leyssen, P.; Neyts, J.; Deubel, V. Poly(I)-poly(C12U) but not ribavirin prevents death in a hamster model of Nipah virus infection. Antimicrob. Agents Chemother. 2006, 50, 1768–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walpita, P.; Cong, Y.; Jahrling, P.B.; Rojas, O.; Postnikova, E.; Yu, S.; Johns, L.; Holbrook, M.R. A VLP-based vaccine provides complete protection against Nipah virus challenge following multiple-dose or single-dose vaccination schedules in a hamster model. npj Vaccines 2017, 2, 21. [Google Scholar] [CrossRef]
- van Doremalen, N.; Lambe, T.; Sebastian, S.; Bushmaker, T.; Fischer, R.; Feldmann, F.; Haddock, E.; Letko, M.; Avanzato, V.A.; Rissanen, I.; et al. A single-dose ChAdOx1-vectored vaccine provides complete protection against Nipah Bangladesh and Malaysia in Syrian golden hamsters. PLoS Negl. Trop. Dis. 2019, 13, e0007462. [Google Scholar] [CrossRef]
- Yoneda, M.; Georges-Courbot, M.C.; Ikeda, F.; Ishii, M.; Nagata, N.; Jacquot, F.; Raoul, H.; Sato, H.; Kai, C. Recombinant measles virus vaccine expressing the Nipah virus glycoprotein protects against lethal Nipah virus challenge. PLoS ONE 2013, 8, e58414. [Google Scholar] [CrossRef]
- DeBuysscher, B.L.; Scott, D.; Thomas, T.; Feldmann, H.; Prescott, J. Peri-exposure protection against Nipah virus disease using a single-dose recombinant vesicular stomatitis virus-based vaccine. npj Vaccines 2016, 1, 16002. [Google Scholar] [CrossRef]
- Welch, S.R.; Tilston, N.L.; Lo, M.K.; Whitmer, S.L.M.; Harmon, J.R.; Scholte, F.E.M.; Spengler, J.R.; Duprex, W.P.; Nichol, S.T.; Spiropoulou, C.F. Inhibition of Nipah Virus by Defective Interfering Particles. J. Infect. Dis. 2020, 221 (Suppl. 4), S460–S470. [Google Scholar] [CrossRef]
- Lo, M.K.; Spengler, J.R.; Krumpe, L.R.H.; Welch, S.R.; Chattopadhyay, A.; Harmon, J.R.; Coleman-McCray, J.D.; Scholte, F.E.M.; Hotard, A.L.; Fuqua, J.L.; et al. Griffithsin Inhibits Nipah Virus Entry and Fusion and Can Protect Syrian Golden Hamsters From Lethal Nipah Virus Challenge. J. Infect. Dis. 2020, 221 (Suppl. 4), S480–S492. [Google Scholar] [CrossRef] [Green Version]
- Lo, M.K.; Spengler, J.R.; Welch, S.R.; Harmon, J.R.; Coleman-McCray, J.D.; Scholte, F.E.M.; Shrivastava-Ranjan, P.; Montgomery, J.M.; Nichol, S.T.; Weissman, D.; et al. Evaluation of a Single-Dose Nucleoside-Modified Messenger RNA Vaccine Encoding Hendra Virus-Soluble Glycoprotein Against Lethal Nipah virus Challenge in Syrian Hamsters. J. Infect. Dis. 2020, 221 (Suppl. 4), S493–S498. [Google Scholar] [CrossRef] [Green Version]
- Torres-Velez, F.J.; Shieh, W.J.; Rollin, P.E.; Morken, T.; Brown, C.; Ksiazek, T.G.; Zaki, S.R. Histopathologic and immunohistochemical characterization of Nipah virus infection in the guinea pig. Vet. Pathol. 2008, 45, 576–585. [Google Scholar] [CrossRef]
- Tanimura, N.; Imada, T.; Kashiwazaki, Y.; Sharifah, S.H. Distribution of viral antigens and development of lesions in chicken embryos inoculated with nipah virus. J. Comp. Pathol. 2006, 135, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Dimitrov, A.S.; Bossart, K.N.; Crameri, G.; Bishop, K.A.; Choudhry, V.; Mungall, B.A.; Feng, Y.R.; Choudhary, A.; Zhang, M.Y.; et al. Potent neutralization of Hendra and Nipah viruses by human monoclonal antibodies. J. Virol. 2006, 80, 891–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEachern, J.A.; Bingham, J.; Crameri, G.; Green, D.J.; Hancock, T.J.; Middleton, D.; Feng, Y.R.; Broder, C.C.; Wang, L.F.; Bossart, K.N. A recombinant subunit vaccine formulation protects against lethal Nipah virus challenge in cats. Vaccine 2008, 26, 3842–3852. [Google Scholar] [CrossRef] [PubMed]
- Marsh, G.A.; de Jong, C.; Barr, J.A.; Tachedjian, M.; Smith, C.; Middleton, D.; Yu, M.; Todd, S.; Foord, A.J.; Haring, V.; et al. Cedar virus: A novel Henipavirus isolated from Australian bats. PLoS Pathog. 2012, 8, e1002836. [Google Scholar] [CrossRef]
- Playford, E.G.; McCall, B.; Smith, G.; Slinko, V.; Allen, G.; Smith, I.; Moore, F.; Taylor, C.; Kung, Y.-H.; Field, H. Human Hendra virus encephalitis associated with equine outbreak, Australia, 2008. Emerg. Infect. Dis. 2010, 16, 219. [Google Scholar] [CrossRef]
- Leeth, C.; Adkins, J.; Hay, A.; Bogers, S.; Potter, A.; Witonsky, S.; Zhu, J. Engrafting Horse Immune Cells into Mouse Hosts for the Study of the Acute Equine Immune Responses. Animals 2021, 11, 2962. [Google Scholar] [CrossRef]
- Dhondt, K.P.; Horvat, B. Henipavirus infections: Lessons from animal models. Pathogens 2013, 2, 264–287. [Google Scholar] [CrossRef] [Green Version]
- Eaton, B.T.; Broder, C.C.; Wang, L.F. Hendra and Nipah viruses: Pathogenesis and therapeutics. Curr. Mol. Med. 2005, 5, 805–816. [Google Scholar] [CrossRef]
- Kasloff, S.B.; Leung, A.; Pickering, B.S.; Smith, G.; Moffat, E.; Collignon, B.; Embury-Hyatt, C.; Kobasa, D.; Weingartl, H.M. Pathogenicity of Nipah henipavirus Bangladesh in a swine host. Sci. Rep. 2019, 9, 5230. [Google Scholar] [CrossRef] [Green Version]
- Pernet, O.; Schneider, B.S.; Beaty, S.M.; LeBreton, M.; Yun, T.E.; Park, A.; Zachariah, T.T.; Bowden, T.A.; Hitchens, P.; Ramirez, C.M.; et al. Evidence for henipavirus spillover into human populations in Africa. Nat. Commun. 2014, 5, 5342. [Google Scholar] [CrossRef] [Green Version]
- Atherstone, C.; Diederich, S.; Weingartl, H.M.; Fischer, K.; Balkema-Buschmann, A.; Grace, D.; Alonso, S.; Dhand, N.K.; Ward, M.P.; Mor, S.M. Evidence of exposure to henipaviruses in domestic pigs in Uganda. Transbound. Emerg. Dis. 2019, 66, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Glennon, E.E.; Restif, O.; Sbarbaro, S.R.; Garnier, R.; Cunningham, A.A.; Suu-Ire, R.D.; Osei-Amponsah, R.; Wood, J.L.N.; Peel, A.J. Domesticated animals as hosts of henipaviruses and filoviruses: A systematic review. Vet. J. 2018, 233, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Middleton, D.J.; Weingartl, H.M. Henipaviruses in their natural animal hosts. Curr. Top. Microbiol. Immunol. 2012, 359, 105–121. [Google Scholar] [CrossRef] [PubMed]
- Baseler, L.; Scott, D.P.; Saturday, G.; Horne, E.; Rosenke, R.; Thomas, T.; Meade-White, K.; Haddock, E.; Feldmann, H.; de Wit, E. Identifying Early Target Cells of Nipah Virus Infection in Syrian Hamsters. PLoS Negl. Trop. Dis. 2016, 10, e0005120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillaume, V.; Lefeuvre, A.; Faure, C.; Marianneau, P.; Buckland, R.; Lam, S.K.; Wild, T.F.; Deubel, V. Specific detection of Nipah virus using real-time RT-PCR (TaqMan). J. Virol. Methods 2004, 120, 229–237. [Google Scholar] [CrossRef] [PubMed]
- de Wit, E.; Prescott, J.; Falzarano, D.; Bushmaker, T.; Scott, D.; Feldmann, H.; Munster, V.J. Foodborne transmission of nipah virus in Syrian hamsters. PLoS Pathog. 2014, 10, e1004001. [Google Scholar] [CrossRef]
- Dang, H.V.; Cross, R.W.; Borisevich, V.; Bornholdt, Z.A.; West, B.R.; Chan, Y.P.; Mire, C.E.; Da Silva, S.C.; Dimitrov, A.S.; Yan, L.; et al. Broadly neutralizing antibody cocktails targeting Nipah virus and Hendra virus fusion glycoproteins. Nat. Struct. Mol. Biol. 2021, 28, 426–434. [Google Scholar] [CrossRef]





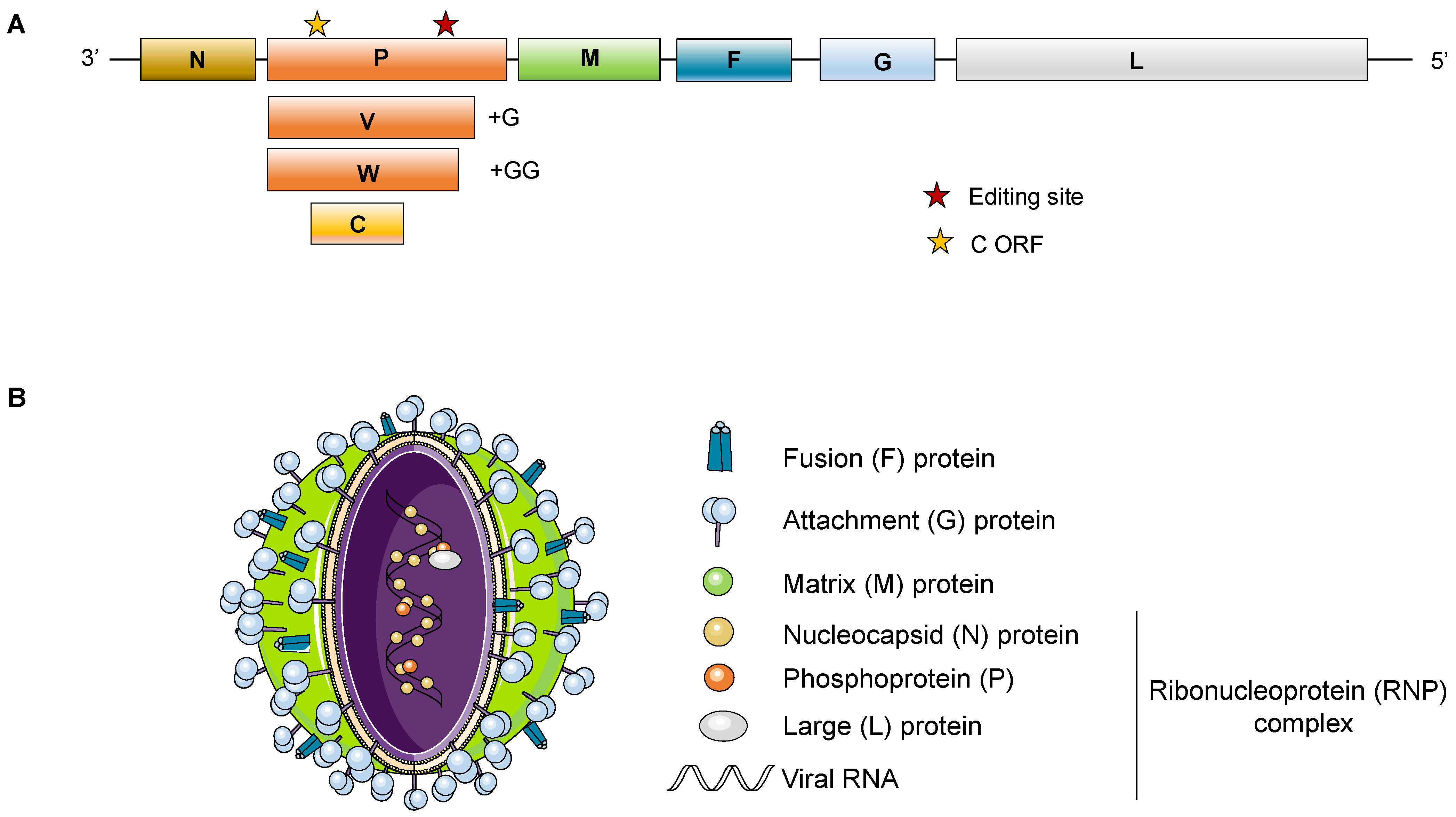{kind=link}
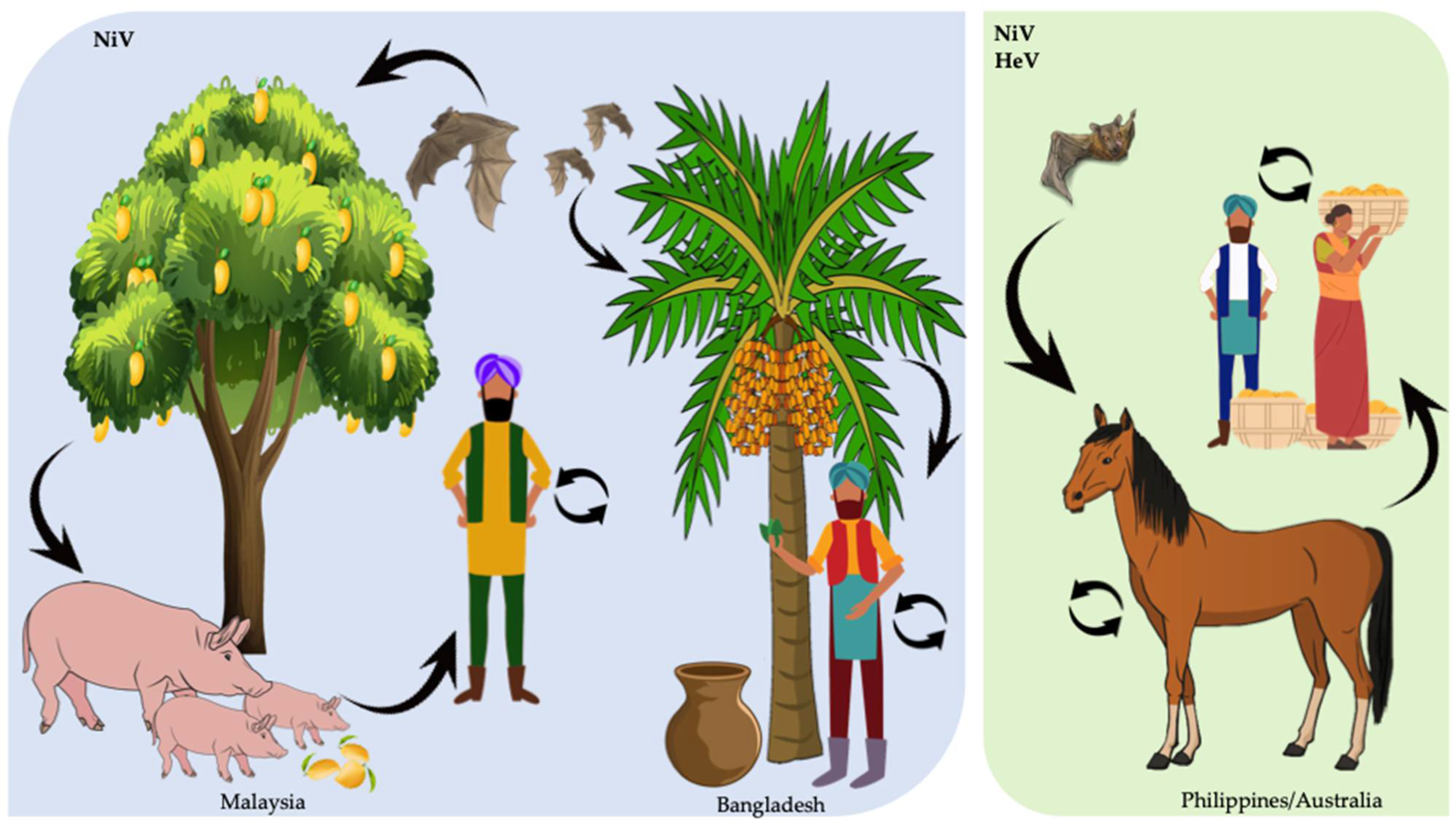{kind=link}
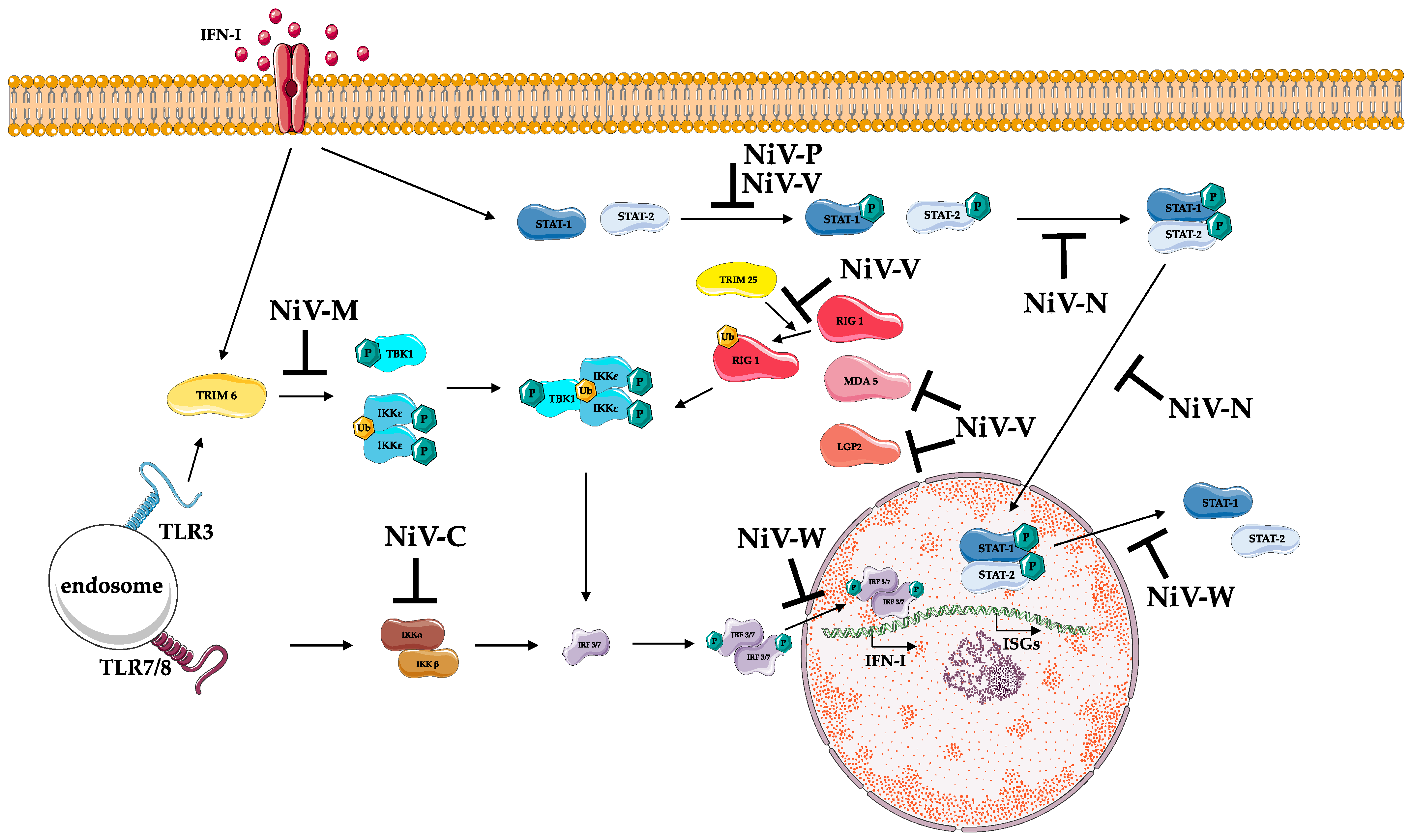{kind=link}
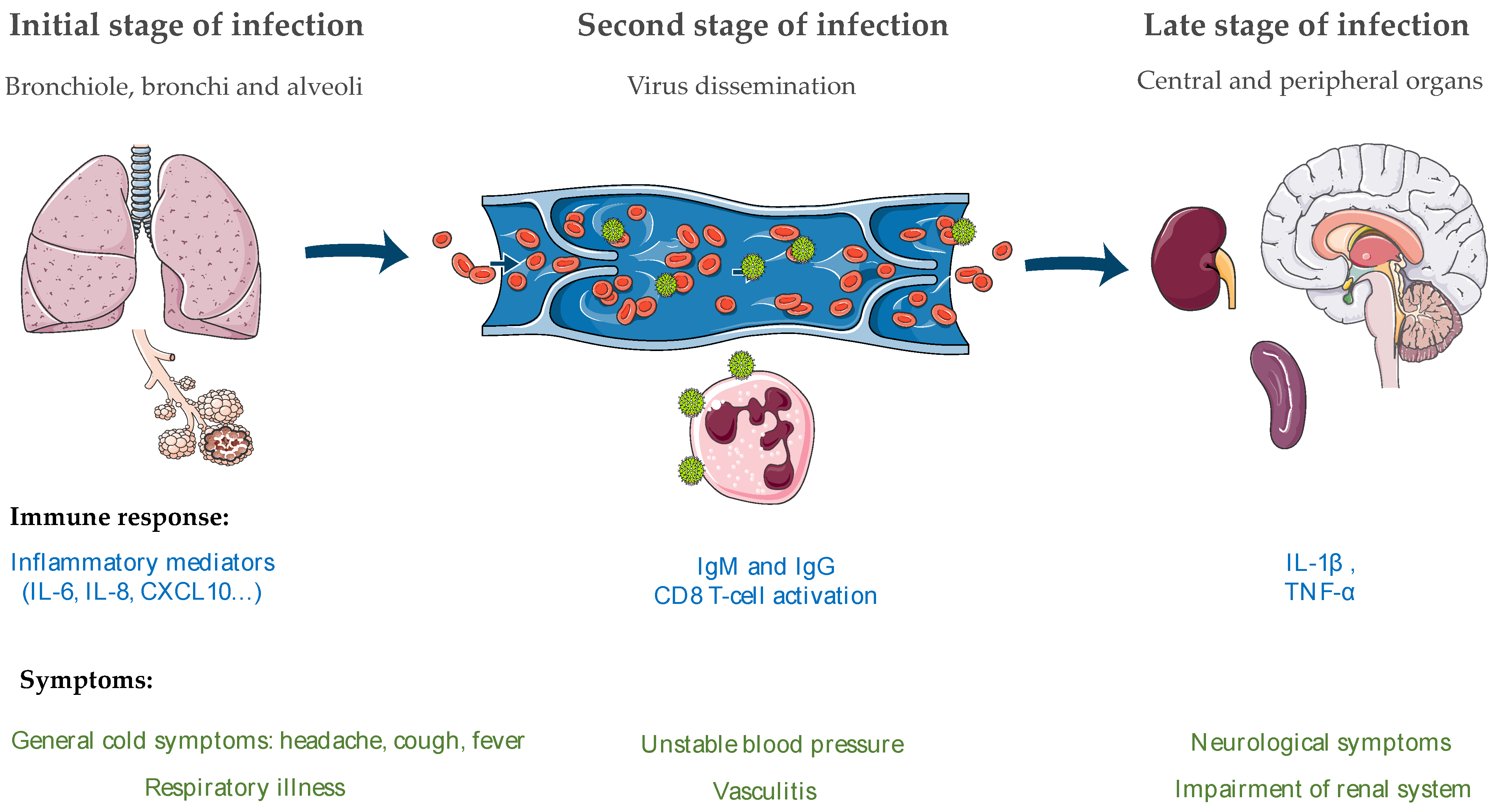{kind=link}
| Animals | Type of Infection | Onset of Illness (Days) | First Clinical Signs | Symptoms | Virus-Positive Tissues or Fluids | References |
|---|---|---|---|---|---|---|
| Bat (P. alecto, P. poliocephalus) | s.c. | N/A | vascular lesions | N/A | blood, kidney, urine, uterus | [67,68,149] |
| s.c. | N/A | mild nephritis, focal vasculitis, cell infiltration in liver | N/A | kidney, urine, rectum | [42,159] | |
| Cat | o.i., i.n., s.c. | 5–7/9 | inappetence, increased respiratory rate | severe respiratory disease, pneumonia, hemorrhagic lungs, vascular lesions, syncytial cells | arteries, veins, lung, spleen, kidney, brain, gastrointestinal tract, urine | [52,149,236,237] |
| o.n.i., s.c. | 4–10/12 | fever, respiratory and neurological disease, depression, constipation, | ulcera, inflammation, meningitis, meningeal vasculitis | oropharynx, tonsil, trachea, lung, brain, kidney, liver, uterus | [17,238,239,240] | |
| Dog | s.c. natural | N/A | no signs of ill health, although wincing several times | reddening and dark patchy discoloration in the lung and the tonsils, white streaks in kidney, spleen and liver inflammation, fibrinoid necrosis, vasculitis and inflammatory infiltrates in kidney, brain, LN, spleen, liver, intestine, lung, expanded meninges | kidney, brain, lymph nodes, spleen, and liver Equivocal NAb | [52,241,242] |
| natural | N/A | fever, respiratory distress, conjunctivitis, nasal discharge | severe pulmonary edema, atrophy, vascular degeneration, syncytia, necrosis, glomerulonephritis, meningitis | N/A | [46,240,243,244] | |
| Ferret | o.n.i., i.n. | 6–9 | fever, depression, serous nasal discharge, dyspnea, hind limb paresis and generalized tremors | cutaneous petechiation, s.c. edema in head and neck, pulmonary petechiation, hemorrhagic lymph nodes, systemic vasculitis, necrosis, splenitis, bronchoalveolitis, endothelial and epithelial syncytial cells | kidney, heart, bladder, LN, lung, spleen, brain, nose, endothelial cells, neurons, bronchoalveolar epithelium, urine | [190,245,246,247] |
| o.n.i., i.n. | 5–9 | pyrexia, depression, cough, dyspnea, hind limb paresis, generalized tremors | multisystemic inflammatory lesions in respiratory tract, spleen, kidneys and liver, neurologic disease, meningitis, encephalitis, hemorrhaging, necrosis, syncytia, bronchoalveolitis, tonsillitis, nasopharyngitis, thrombocytopenia, multisystemic vasculitis | nasal turbinates, pharynx, retropharyngeal lymph nodes, spleen, lung, liver, kidney, LN, uterus, ovaries, heart, brain, bladder, mouth, rectum, vascular endothelium, feces, neurons, glial cells, urine | [246,247,248,249,250,251,252,253] | |
| Horse | s.c., i.n., o.n.i., i.v. | 5–16 | fever, high cardiac rates, depression, dyspnea, recumbency, loss of appetite, neurological signs | lung edema, thrombosis, hemorrhage, tissue necrosis, syncytia, vascular damage | kidney, lung, mouth, nasal cavities, urine | [48,49,89,149,254,255,256] |
| natural | N/A | neurological signs | vascular damage, meningitis (only brain and spinal cord were analyzed) | brain, spinal cord seroconversion | [240,257] | |
| Monkey (African green monkey, AGM) | i.n., i.t. | 7 | piloerection, respiratory distress, nasal discharge, depression, seizures, muscle fasciculations | severe systemic vasculitis, necrosis, hemorrhage and edema in most organs, splenomegaly, hemorrhaging, syncytial cells, meningitis | tonsils, trachea, lungs, heart, liver, spleen, kidney, pancreas, intestine, LN, brain, testes/ovaries, bone marrow, urine | [115,258,259] |
| i.n., i.t., o.i. | 7–12 | fever, loss of appetite, respiratory disease, lethargy, rash, depression, behavioral changes | severe systemic vasculitis, hemorrhage and edema in most organs, thrombocytopenia, meningitis | blood, trachea, lungs, heart, liver, spleen, kidney, pancreas, intestine, LN, brain, testes/ovaries, bone marrow, rectum, urine | [24,59,259,260,261,262,263,264,265,266,267,268] | |
| Monkey (Squirrel Monkey) | N/A | N/A | N/A | N/A | N/A | N/A |
| i.v., i.n. | 7–19 | respiratory disease, loss of appetite, depression, uncoordinated motor movements | inflammation of lung parenchyma, mild vasculitis | spleen, liver, lung, heart, bladder, kidney, LN, spinal cord, brain | [269] | |
| Pig | i.n., o.n.i. | 4/5 | fever, loss of appetite, cough, respiratory distress, depression, uncoordinated movements | pulmonary edema, hemorrhages in lung, kidney and LN, syncytial cells, inflammation, vasculitis, necrosis | Tonsils, lung, nasal turbinates, LN, olfactory bulb, mouth, ocular secretions, rectum | [270,271,272,273] |
| o.n.i., o.i., s.c., i.n., oc.i. | 7–21 | fever, nasal discharge, coughing, locomotor disturbances, agitation, muscle fasciculations, paresis, seizures | systemic vasculitis, vasculopathy, alveolitis, thrombosis, cell necrosis, pulmonary edema and inflammation, renal tubular degeneration, syncytia, meningitis, | tonsils, lung, LN, olfactory bulb, nose, oropharynx, spleen, endothelium, lymphatic vessels, kidneys, brain, blood, urine | [14,17,146,244,273,274,275,276,277,278] | |
| ID mice (IFNAR-KO, NSG) | i.n., i.p. | 3–21 | agitation, lack of grooming, grimace, loss in body weight, lordosis, aggression, locomotor disability, head tilt, and paralysis | lung and brain congestion, hemorrhages, vasculitis, necrosis, meningitis, encephalitis | brain, lung, spleen, liver | [23] |
| i.p., i.n., i.cer. | 6–10 | agitation, lack of grooming, grimace, loss in body weight, lordosis, aggression, locomotor disability, head tilt, and paralysis | inflammation, edema, focal necrosis in lung and vasculitis microscopic lesions in the brain, liver and kidney inflammation, syncytial cells, meningeal inflammation | brain, lung, spleen, liver | [23,121,279,280] | |
| IC mice (C57BL/6, BALB/c, CB6F1/J) | i.n., s.c. | 10–21 | depression, ataxia, hypersensitivity, and tremors | necrosis, ulceration, encephalitis | respiratory tract, olfactory epithelium, brain | [52,281] |
| i.n., i.p. i.cer. | 4 | N/A | subclinical, self-limiting respiratory infection | lung, spleen | [23,278,282,283,284,285,286,287] | |
| Hamster | i.p. | 3–25 | breathing difficulties, paralysis, and trembling limbs | pulmonary edema, inflammation in lung and spleen, necrosis, syncytia cells, meningitis | lung, heart, liver, spleen, brain, serum, urine | [288,289,290,291] |
| i.n., i.p., a.e., fom., d.c. | 4–15 | breathing difficulties, imbalance, limb paralysis, lethargy, muscle twitching | damage in lung, liver, kidney, heart, and brain, fibrinoid necrosis with surrounding inflammation in blood vessels, syncytial cells, necrosis, vasculitis, thrombosis, meningitis | lung, kidney, spleen, liver, heart, spinal cord, brain, urine | [22,93,111,124,147,288,289,292,293,294,295,296,297,298,299,300,301,302,303] | |
| Guinea pig | s.c., i.n., i.d. | 7–15 days | inappetence, increased respiratory rate, head tilt, ataxia, torticollis, depression | pneumonia, cyanose, oedema in gastrointestinal tract, systemic vascular disease in arteries, veins, lung, kidney, spleen, lymph nodes, syncytia in lungs | arteries, veins, lung, kidney, spleen, lymph nodes | [21,52,160,237] |
| i.p., i.n. | 7–9 days | ruffled hair, weight loss, abnormal behavior, ataxia | oedema, systemic vasculitis, endothelial syncytial cells, cell necrosis, lung hemorrhages | heart, spleen, kidney, lung, brain, LN, thymus, blood, ovaries, uterus | [22,159,304] | |
| Rat | s.c. | N/A | no histological lesions | N/A | virus not observed Equivocal NAb | [52] |
| N/A | N/A | N/A | N/A | N/A | N/A | |
| Rabbit | s.c. | N/A | no histological lesions | N/A | virus not observed | [52] |
| N/A | N/A | N/A | N/A | N/A | N/A | |
| Chicken | s.c. | N/A | no histological lesions | N/A | virus not observed | [52] |
| allantoic or yolk sac inoculation | 5–7 | only reported in embryos: 4–7 | only reported in embryos: brain hemorrhage, congestion and hemorrhage in the skin of the toes and in the kidneys, syncytial cells, necrosis | only reported in embryos, lung, heart, liver, kidney, spleen, skin, CNS, blood vessels | [305] |
| Host | Human | Bat | Cat | Dog | Ferret | Horse | Monkey | Pig | IC Mice | Hamster | Guinea Pig | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Innate | Inflammation | |||||||||||
| Interferon expression | ||||||||||||
| Complement | ||||||||||||
| Adaptative | Lymphocyte activation | |||||||||||
| Immunoglobulines | ||||||||||||
| Neutralizing antibodies | ||||||||||||
| Lymphopenia | ||||||||||||
| Inhibition or decreased levels or not present with henipavirus infection | |
| Activation or presence with henipavirus infection | |
| Not yet described with henipavirus infection |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lawrence, P.; Escudero-Pérez, B. Henipavirus Immune Evasion and Pathogenesis Mechanisms: Lessons Learnt from Natural Infection and Animal Models. Viruses 2022, 14, 936. https://doi.org/10.3390/v14050936
Lawrence P, Escudero-Pérez B. Henipavirus Immune Evasion and Pathogenesis Mechanisms: Lessons Learnt from Natural Infection and Animal Models. Viruses. 2022; 14(5):936. https://doi.org/10.3390/v14050936
Chicago/Turabian StyleLawrence, Philip, and Beatriz Escudero-Pérez. 2022. "Henipavirus Immune Evasion and Pathogenesis Mechanisms: Lessons Learnt from Natural Infection and Animal Models" Viruses 14, no. 5: 936. https://doi.org/10.3390/v14050936
APA StyleLawrence, P., & Escudero-Pérez, B. (2022). Henipavirus Immune Evasion and Pathogenesis Mechanisms: Lessons Learnt from Natural Infection and Animal Models. Viruses, 14(5), 936. https://doi.org/10.3390/v14050936