
Detection of Lassa Virus-Reactive IgG Antibodies in Wild Rodents: Validation of a Capture Enzyme-Linked Immunological Assay


 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of Laboratory Samples
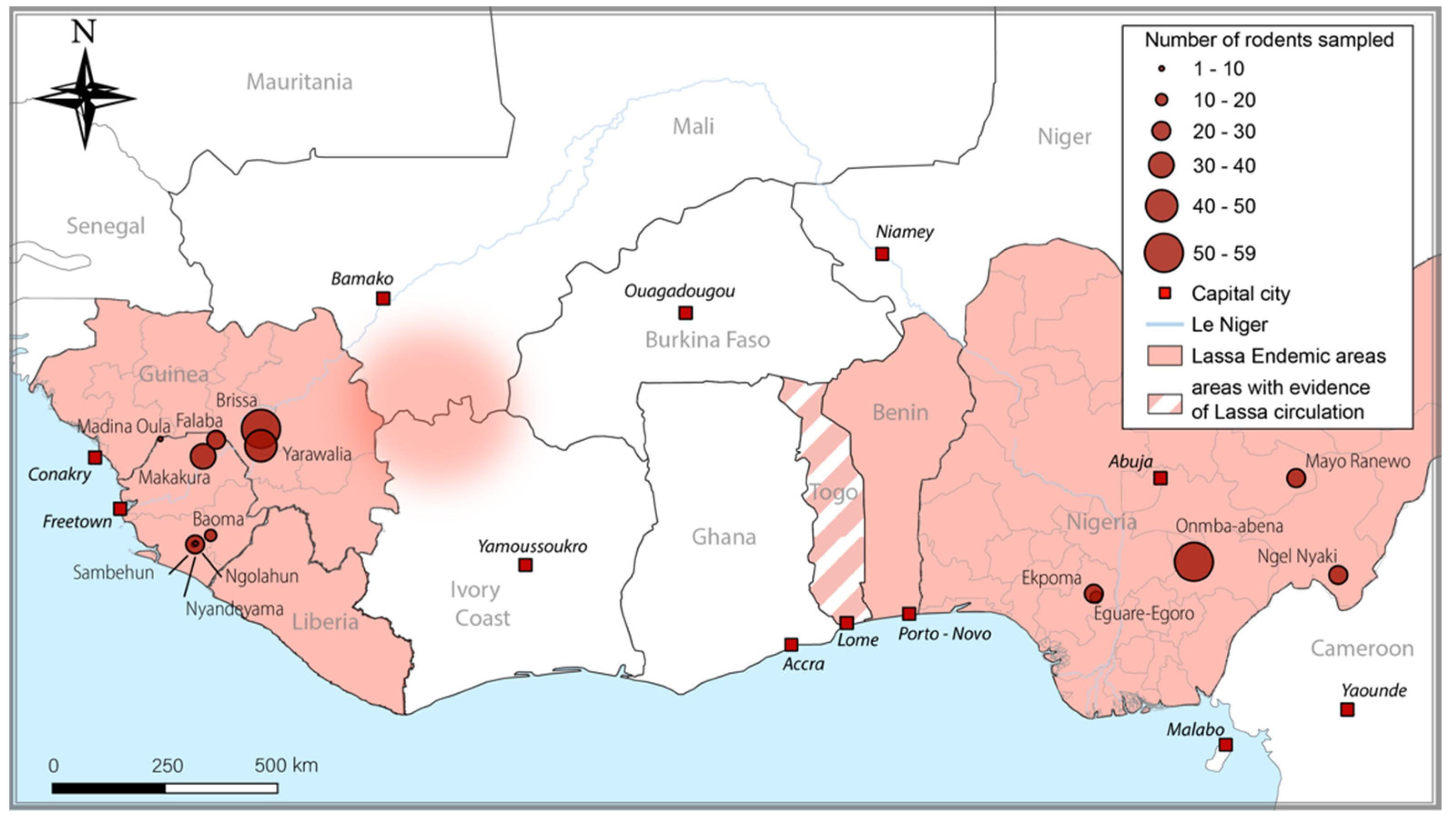
2.2. Origin of Wild Samples
2.3. ELISA
2.4. IFA
2.5. Elution of Dried Blood on Filter Paper
2.6. Statistical Analysis
3. Results
3.1. Laboratory-Bred Mastomys
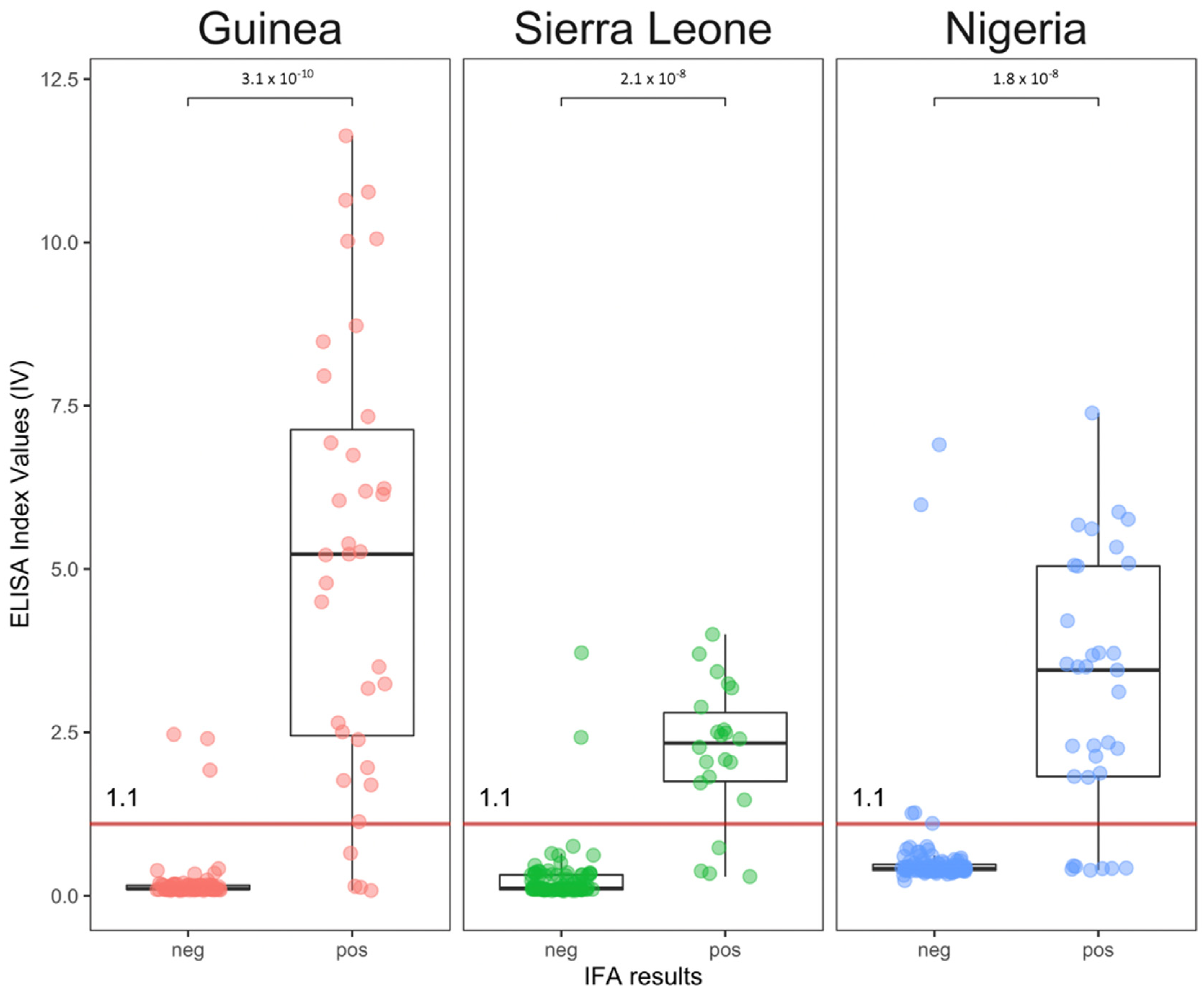
3.2. Wild Mastomys
3.3. Concordance between the Tests
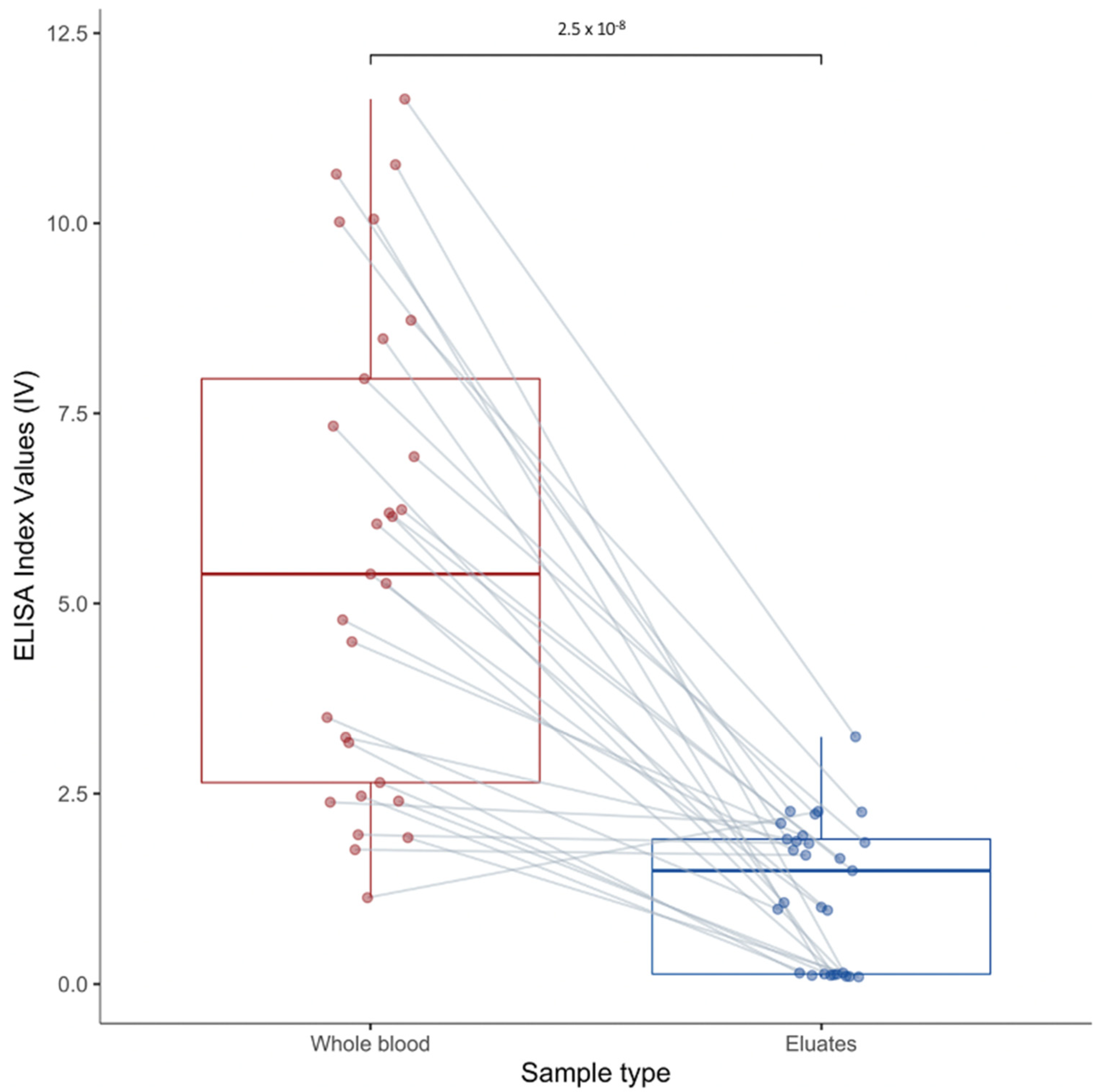
3.4. Whole Blood vs. Dried Blood on Filter Paper
4. Discussion
4.1. ELISA versus IFA
4.2. Dried Blood Spot and Serology
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frame, J.D.; Baldwin, J.M.J.; Gocke, D.J.; Troup, J.M. Lassa fever, a new virus disease of man from West Africa. Am. J. Trop. Med. Hyg. 1970, 19, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Demby, A.H.; Inapogui, A.; Kargbo, K.; Koninga, J.; Kourouma, K.; Kanu, J.; Coulibaly, M.; Wagoner, K.D.; Ksiazek, T.G.; Peters, C.J.; et al. Lassa fever in Guinea: II. Distribution and prevalence of Lassa virus infection in small mammals. Vector Borne Zoonotic Dis. 2001, 1, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Olayemi, A.; Cadar, D.; Magassouba, N.; Obadare, A.; Kourouma, F.; Oyeyiola, A.; Fasogbon, S.; Igbokwe, J.; Rieger, T.; Bockholt, S.; et al. New hosts of the Lassa virus. Sci. Rep. 2016, 6, 25280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carey, D.E.; Kemp, G.E.; White, H.A.; Pinneo, L.; Addy, R.F.; Dom, A.L.M.D.; Stroh, G.; Casals, J.; Henderson, B.E. Lassa fever epidemiological aspects of the 1970 epidemic, Jos, Nigeria. R. Soc. Trop. Med. Hyg. 1972, 66, 402–408. [Google Scholar] [CrossRef]
- Kenmoe, S.; Tchatchouang, S.; Ebogo-Belobo, J.T.; Ka’e, A.C.; Mahamat, G.; Guiamdjo Simo, R.E.; Bowo-Ngandji, A.; Demeni Emoh, C.P.; Che, E.; Tchami Ngongang, D.; et al. Systematic review and meta-analysis of the epidemiology of Lassa virus in humans, rodents and other mammals in sub-Saharan Africa. PLoS Negl. Trop. Dis. 2020, 14, e0008589. [Google Scholar] [CrossRef]
- Sogoba, N.; Rosenke, K.; Adjemian, J.; Diawara, S.I.; Maiga, O.; Keita, M.; Konaté, D.; Keita, A.S.; Sissoko, I.; Boisen, M.; et al. Lassa virus seroprevalence in Sibirilia commune, Bougouni district, Southern Mali. Emerg. Infect. Dis. 2016, 22, 657–663. [Google Scholar] [CrossRef] [Green Version]
- Fichet-Calvet, E.; Becker-Ziaja, B.; Koivogui, L.; Gunther, S. Lassa serology in natural populations of rodents and horizontal transmission. Vector Borne Zoonotic Dis. 2014, 14, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Wulff, H.; Lange, J.V. Indirect immunofluorescence for the diagnosis of Lassa fever infection. Bull. World Health Organ. 1975, 52, 429–436. [Google Scholar]
- Jahrling, P.B.; Frame, J.D.; Rhoderick, J.B.; Monson, M.H. Endemic Lassa fever in Liberia. IV. Selection of optimally effective plasma for treatment by passive immunization. Trans. R. Soc. Trop. Med. Hyg. 1985, 79, 380–384. [Google Scholar] [CrossRef]
- McCormick, J.B.; Webb, P.A.; Krebs, J.W.; Johnson, K.M.; Smith, E.S. A prospective study of the epidemiology and ecology of Lassa fever. J. Infect. Dis. 1987, 155, 437–444. [Google Scholar] [CrossRef]
- Bausch, D.G.; Rollin, P.E.; Demby, A.H.; Coulibaly, M.; Kanu, J.; Conteh, A.S.; Wagoner, K.D.; McMullan, L.K.; Bowen, M.D.; Peters, C.J.; et al. Diagnosis and clinical virology of Lassa fever as evaluated by enzyme-linked immunosorbent assay, indirect fluorescent-antibody test, and virus isolation. J. Clin. Microbiol. 2000, 38, 2670–2677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branco, L.M.; Grove, J.N.; Boisen, M.L.; Shaffer, J.G.; Goba, A.; Fullah, M.; Momoh, M.; Grant, D.S.; Garry, R.F. Emerging trends in Lassa fever: Redefining the role of immunoglobulin M and inflammation in diagnosing acute infection. Virol. J. 2011, 8, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriel, M.; Adomeh, D.I.; Ehimuan, J.; Oyakhilome, J.; Omomoh, E.O.; Ighodalo, Y.; Olokor, T.; Bonney, K.; Pahlmann, M.; Emmerich, P.; et al. Development and evaluation of antibody-capture immunoassays for detection of Lassa virus nucleoprotein-specific immunoglobulin M and G. PLoS Negl. Trop. Dis. 2018, 12, e0006361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaffer, J.G.; Schieffelin, J.S.; Momoh, M.; Goba, A.; Kanneh, L.; Alhasan, F.; Gbakie, M.; Engel, E.J.; Bond, N.G.; Hartnett, J.N.; et al. Space-time trends in Lassa fever in Sierra Leone by ELISA serostatus, 2012–2019. Microorganisms 2021, 9, 586. [Google Scholar] [CrossRef]
- Bangura, U.; Buanie, J.; Lamin, J.; Davis, C.; Bongo, G.N.; Dawson, M.; Ansumana, R.; Sondufu, D.; Thomson, E.C.; Sahr, F.; et al. Lassa virus circulation in small mammal populations in Bo district, Sierra Leone. Biology 2021, 10, 28. [Google Scholar] [CrossRef]
- Borremans, B.; Vossen, R.; Becker-Ziaja, B.; Gryseels, S.; Hughes, N.; Van Gestel, M.; Van Houtte, N.; Gunther, S.; Leirs, H. Shedding dynamics of Morogoro virus, an African arenavirus closely related to Lassa virus, in its natural reservoir host Mastomys natalensis. Sci. Rep. 2015, 5, 10445. [Google Scholar] [CrossRef]
- Marien, J.; Lo Iacono, G.; Rieger, T.; Magassouba, N.; Gunther, S.; Fichet-Calvet, E. Households as hotspots of Lassa fever? Assessing the spatial distribution of Lassa virus-infected rodents in rural villages of Guinea. Emerg. Microbes Infect. 2020, 9, 1055–1064. [Google Scholar] [CrossRef]
- Olayemi, A.; Oyeyiola, A.; Obadare, A.; Igbokwe, J.; Adesina, A.S.; Onwe, F.; Ukwaja, K.N.; Ajayi, N.A.; Rieger, T.; Gunther, S.; et al. Widespread arenavirus occurrence and seroprevalence in small mammals, Nigeria. Parasites Vectors 2018, 11, 416. [Google Scholar] [CrossRef]
- Safronetz, D.; Rosenke, K.; Fischer, R.J.; LaCasse, R.A.; Scott, D.P.; Saturday, G.; Hanley, P.W.; Maiga, O.; Sogoba, N.; Schwan, T.G.; et al. Establishment of a genetically confirmed breeding colony of mastomys natalensis from wild-caught founders from West Africa. Viruses 2021, 13, 590. [Google Scholar] [CrossRef]
- Emmerich, P.; Mika, A.; Schmitz, H. Detection of serotype-specific antibodies to the four dengue viruses using an immune complex binding (ICB) ELISA. PLoS Negl. Trop. Dis. 2013, 7, e2580. [Google Scholar] [CrossRef] [Green Version]
- Borremans, B. Ammonium improves elution of fixed dried blood spots without affecting immunofluorescence assay quality. Trop. Med. Int. Health 2014, 19, 413–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Drug Administration. Statistical Guidance on Reporting Results from Studies Evaluating Diagnostic Tests. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/statistical-guidance-reporting-results-studies-evaluating-diagnostic-tests-guidance-industry-and-fda (accessed on 5 May 2022).
- Watson, P.F.; Petrie, A. Method agreement analysis: A review of correct methodology. Theriogenology 2010, 73, 1167–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHugh, M.L. Interrater reliability: The kappa statistic. Biochem. Med. 2012, 22, 276–282. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Gettings, J.R.; Lopez, J.E.; Krishnavahjala, A.; Armstrong, B.A.; Thompson, A.T.; Yabsley, M.J. Antibodies to borrelia turicatae in experimentally infected dogs cross-react with borrelia burgdorferi serologic assays. J. Clin. Microbiol. 2019, 57, e00628-19. [Google Scholar] [CrossRef] [Green Version]
- Lepper, A.W.; Pearson, C.W. The indirect fluorescent antibody test for the detection of circulating antibodies in bovine tuberculosis. Aust. Vet. J. 1975, 51, 256–261. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Barbic, L.; Stevanovic, V.; Savic, V.; Mrzljak, A.; Bogdanic, M.; Tabain, I. Comparison of indirect immunofluorescence and western blot method in the diagnosis of hantavirus infections. World J. Methodol. 2021, 11, 294–301. [Google Scholar] [CrossRef]
- Xie, W.; Ao, C.; Yang, Y.; Liu, Y.; Liang, R.; Zeng, Z.; Ye, G.; Xiao, S.; Fu, Z.F.; Dong, W.; et al. Two critical N-terminal epitopes of the nucleocapsid protein contribute to the cross-reactivity between porcine epidemic diarrhea virus and porcine transmissible gastroenteritis virus. J. Gen. Virol. 2019, 100, 206–216. [Google Scholar] [CrossRef]
- Biral, N.V.; Azevedo Santos, H.; Senne, N.A.; Paulino, P.G.; Camilo, T.A.; Tassinari, W.S.; Silva, V.L.; Santos, F.N.; Angelo, I.D.C. A cross-sectional study of Leishmania spp. in draft horses from the Distrito Federal, Brazil: Seroprevalence, spatial distribution, and associated factors. Prev. Vet. Med. 2021, 195, 105467. [Google Scholar] [CrossRef]
- Deelder, A.M.; Kornelis, D. A comparison of the IFA and the ELISA for the demonstration of antibodies against schistosome gut-associated polysaccharide antigens in schistosomiasis. Z. Parasitenkd. 1980, 64, 65–75. [Google Scholar] [CrossRef]
- Douglas, K.O.; Samuels, T.A.; Iheozor-Ejiofor, R.; Vapalahti, O.; Sironen, T.; Gittens-St Hilaire, M. Serological evidence of human orthohantavirus infections in Barbados, 2008 to 2016. Pathogens 2021, 10, 571. [Google Scholar] [CrossRef]
- Borremans, B.; Leirs, H.; Gryseels, S.; Gunther, S.; Makundi, R.; de Bellocq, J.G. Presence of mopeia virus, an African arenavirus, related to biotope and individual rodent host characteristics: Implications for virus transmission. Vector Borne Zoonotic Dis. 2011, 11, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- de Bellocq, J.G.; Borremans, B.; Katakweba, A.; Makundi, R.; Baird, S.J.; Becker-Ziaja, B.; Günther, S.; Leirs, H. Sympatric occurrence of 3 arenaviruses, Tanzania. Emerg. Infect. Dis. 2010, 16, 692–695. [Google Scholar] [CrossRef] [PubMed]
- Marien, J.; Borremans, B.; Gryseels, S.; Broecke, B.V.; Becker-Ziaja, B.; Makundi, R.; Massawe, A.; Reijniers, J.; Leirs, H. Arenavirus dynamics in experimentally and naturally infected rodents. Ecohealth 2017, 14, 463–473. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cohort | ELISA + | ELISA − | Total |
|---|---|---|---|
| Positive | 59 | 3 | 62 |
| Negative | 0 | 42 | 42 |
| Total | 59 | 45 | 104 |
| Country | N | ELISA + (% of N) | IFA + (% of N) | p-Values * | X2 |
|---|---|---|---|---|---|
| Guinea | 118 | 34 (28.8) | 35 (29.7) | 0.886 | 0.021 |
| Sierra Leone | 106 | 20 (18.9) | 22 (20.8) | 0.730 | 0.119 |
| Nigeria | 137 | 31 (22.6) | 33 (24.1) | 0.775 | 0.082 |
| Total | 361 | 85 (23.5) | 90 (24.9) | 0.664 | 0.189 |
| IFA − (%) | IFA + (%) | Total | |
|---|---|---|---|
| ELISA − | 250 (91) | 26 (9) | 276 |
| ELISA + | 10 (12) | 75 (88) | 85 |
| Total | 260 (75) | 101 (25) | 361 |
| Guinea (95% CI) | Sierra Leone (95% CI) | Nigeria (95% CI) | Full Dataset (95% CI) | |
|---|---|---|---|---|
| Overall agreement | 94.1% (89.8–98.3) | 94.3% (89.9–98.7) | 91.2% (86.5–96) | 93.1% (90.5–95.7) |
| Positive agreement | 88.6% (78–99.1) | 81.8% (65.7–97.9) | 78.8% (64.8–92.7) | 83.3% (75.6–91) |
| Negative agreement | 96.4% (92.4–100.4) | 97.6% (94.4–100.9) | 95.2% (91.1–99.3) | 96.3% (94.1–98.6) |
| Cohen kappa | 0.857 (0.75–0.96) | 0.822 (0.68–0.96) | 0.755 (0.62–0.89) | 0.811 (0.74–0.88) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soubrier, H.; Bangura, U.; Hoffmann, C.; Olayemi, A.; Adesina, A.S.; Günther, S.; Oestereich, L.; Fichet-Calvet, E. Detection of Lassa Virus-Reactive IgG Antibodies in Wild Rodents: Validation of a Capture Enzyme-Linked Immunological Assay. Viruses 2022, 14, 993. https://doi.org/10.3390/v14050993
Soubrier H, Bangura U, Hoffmann C, Olayemi A, Adesina AS, Günther S, Oestereich L, Fichet-Calvet E. Detection of Lassa Virus-Reactive IgG Antibodies in Wild Rodents: Validation of a Capture Enzyme-Linked Immunological Assay. Viruses. 2022; 14(5):993. https://doi.org/10.3390/v14050993
Chicago/Turabian StyleSoubrier, Hugo, Umaru Bangura, Chris Hoffmann, Ayodeji Olayemi, Adetunji Samuel Adesina, Stephan Günther, Lisa Oestereich, and Elisabeth Fichet-Calvet. 2022. "Detection of Lassa Virus-Reactive IgG Antibodies in Wild Rodents: Validation of a Capture Enzyme-Linked Immunological Assay" Viruses 14, no. 5: 993. https://doi.org/10.3390/v14050993
APA StyleSoubrier, H., Bangura, U., Hoffmann, C., Olayemi, A., Adesina, A. S., Günther, S., Oestereich, L., & Fichet-Calvet, E. (2022). Detection of Lassa Virus-Reactive IgG Antibodies in Wild Rodents: Validation of a Capture Enzyme-Linked Immunological Assay. Viruses, 14(5), 993. https://doi.org/10.3390/v14050993