
Glucose Reduces Norovirus Binding to Enterobacter cloacae and Alters Gene Expression of Bacterial Surface Structures in a Growth Phase Dependent Manner


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Strains and Virus Production
2.2. Norovirus-Bacteria Attachment Assay
2.3. RNA Extraction and DNase Treatment
2.4. RT-qPCR
2.5. Statistical Analysis
3. Results
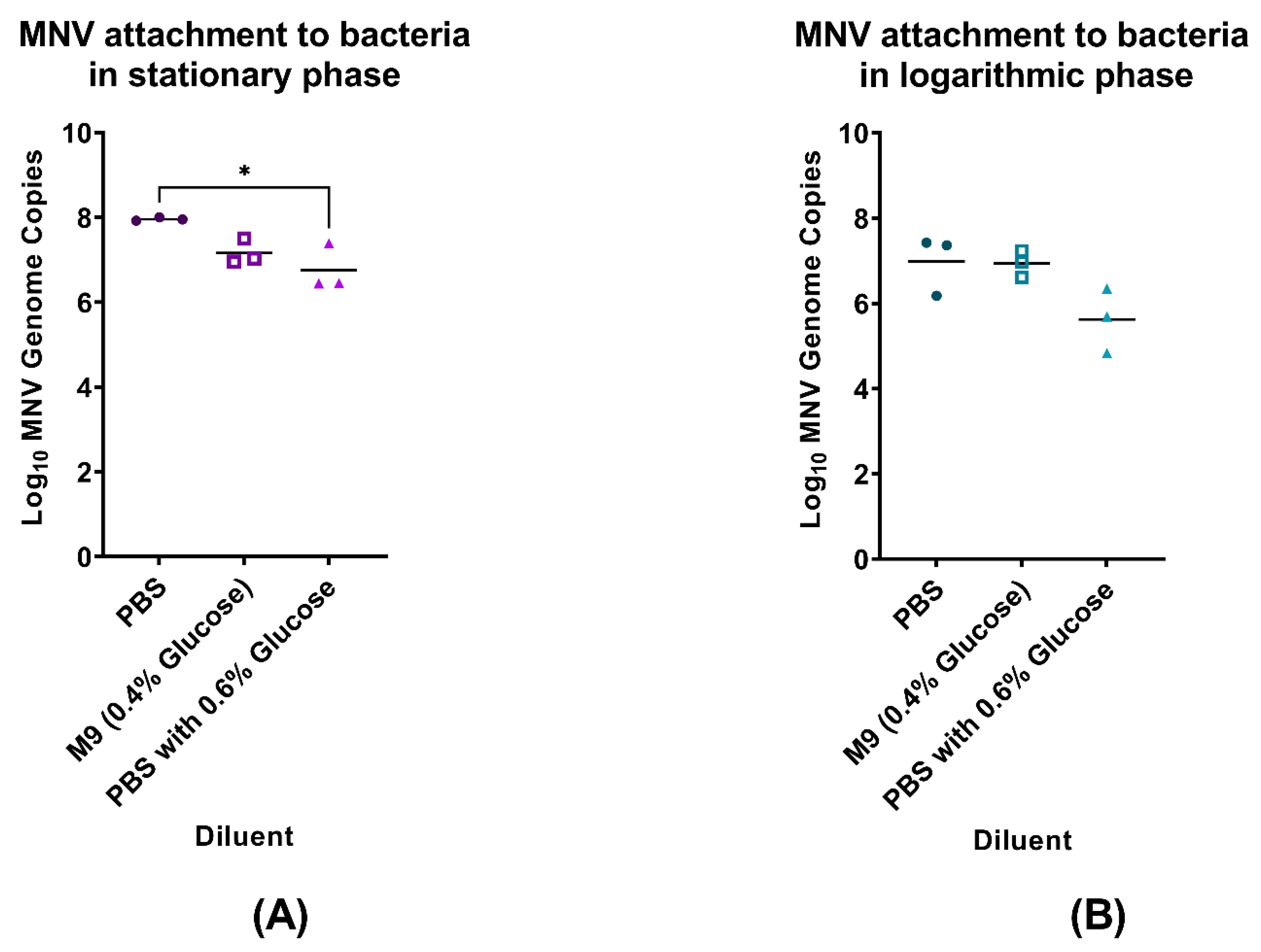
3.1. Impact of Bacterial Growth Medium on MNV Binding to Enterobacter cloacae
3.2. Effect of Bacterial Medium, Growth Phase, and MNV Attachment on E. cloacae Gene Expression
3.3. E. cloacae Gene Expression during HNoV-VLP Attachment Is Not Altered by Incubation Medium or Growth Phase
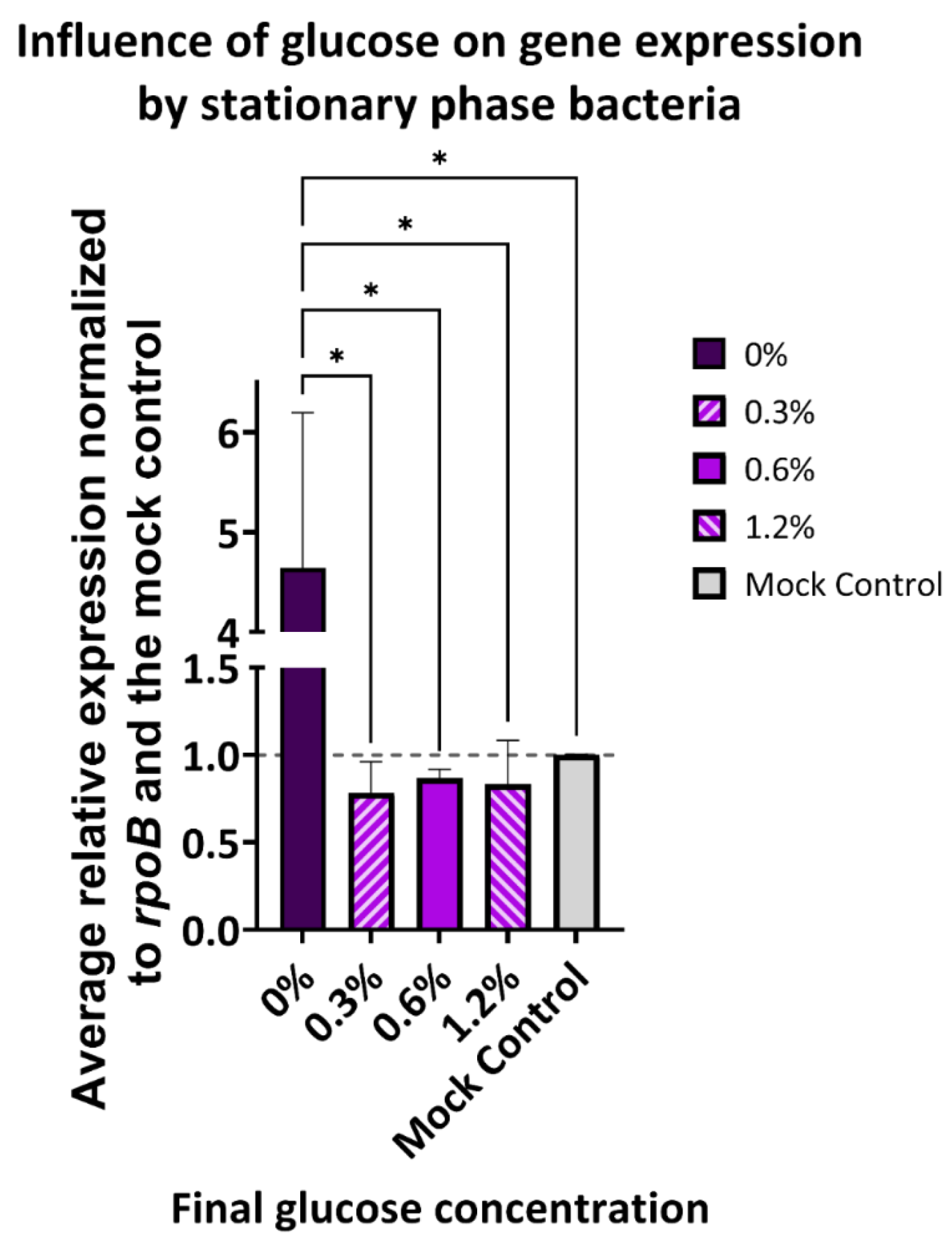
3.4. Stationary Phase E. cloacae MNV Attachment Levels Are Altered by Glucose While flgB Expression Following MNV Incubation Is Not
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Almand, E.A.; Moore, M.D.; Outlaw, J.; Jaykus, L.-A. Human norovirus binding to select bacteria representative of the human gut microbiota. PLoS ONE 2017, 12, e0173124. [Google Scholar] [CrossRef] [PubMed]
- Riddle, M.S.; Walker, R.I. Status of vaccine research and development for norovirus. Vaccine 2016, 34, 2895–2899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Graaf, M.; Van Beek, J.; Koopmans, M. Human norovirus transmission and evolution in a changing world. Nat. Rev. Genet. 2016, 14, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Green, K.Y. Norovirus infection in immunocompromised hosts. Clin. Microbiol. Infect. 2014, 20, 717–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, P.; Farkas, T.; Marionneau, S.; Zhong, W.; Ruvoën-Clouet, N.; Morrow, A.L.; Altaye, M.; Pickering, L.K.; Newburg, D.S.; Le Pendu, J.; et al. Noroviruses Bind to Human ABO, Lewis, and Secretor Histo–Blood Group Antigens: Identification of 4 Distinct Strain-Specific Patterns. J. Infect. Dis. 2003, 188, 19–31. [Google Scholar] [CrossRef]
- Marionneau, S.; Ruvoën, N.; Le Moullac–Vaidye, B.; Clement, M.; Cailleau–Thomas, A.; Ruiz–Palacois, G.; Huang, P.; Jiang, X.; Le Pendu, J. Norwalk virus binds to histo-blood group antigens present on gastroduodenal epithelial cells of secretor individuals. Gastroenterology 2002, 122, 1967–1977. [Google Scholar] [CrossRef]
- Jones, M.K.; Watanabe, M.; Zhu, S.; Graves, C.L.; Keyes, L.R.; Grau, K.R.; Gonzalez-Hernandez, M.B.; Iovine, N.M.; Wobus, C.E.; Vinjé, J.; et al. Enteric bacteria promote human and mouse norovirus infection of B cells. Science 2014, 346, 755–759. [Google Scholar] [CrossRef] [Green Version]
- Springer, G.F.; Williamson, P.; Brandes, W.C. Blood group activity of gram-negative bacteria. J. Exp. Med. 1961, 113, 1077–1093. [Google Scholar] [CrossRef]
- Miura, T.; Sano, D.; Suenaga, A.; Yoshimura, T.; Fuzawa, M.; Nakagomi, T.; Nakagomi, O.; Okabe, S. Histo-Blood Group Antigen-Like Substances of Human Enteric Bacteria as Specific Adsorbents for Human Noroviruses. J. Virol. 2013, 87, 9441–9451. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Breiman, A.; Le Pendu, J.; Uyttendaele, M. Binding to histo-blood group antigen-expressing bacteria protects human norovirus from acute heat stress. Front. Microbiol. 2015, 6, 659. [Google Scholar] [CrossRef]
- Baldridge, M.T.; Nice, T.J.; McCune, B.T.; Yokoyama, C.C.; Kambal, A.; Wheadon, M.; Diamond, M.S.; Ivanova, Y.; Artyomov, M.; Virgin, H.W. Commensal microbes and interferon-λ determine persistence of enteric murine norovirus infection. Science 2015, 347, 266–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosby, C.A.; Bhar, S.; Phillips, M.B.; Edelmann, M.J.; Jones, M.K. Interaction with mammalian enteric viruses alters outer membrane vesicle production and content by commensal bacteria. J. Extracell. Vesicles 2022, 11, e12172. [Google Scholar] [CrossRef] [PubMed]
- Ishihama, A. Modulation of the nucleoid, the transcription apparatus, and the translation machinery in bacteria for stationary phase survival. Genes Cells 1999, 4, 135–143. [Google Scholar] [CrossRef]
- Bleotu, C.; Chifiriuc, M.C.; Mircioaga, D.; Sandulescu, O.; Aldea, I.M.; Banu, O.; Ion, D.; Diaconu, C.D.; Marinescu, F.; Lazar, V. The influence of nutrient culture media on Escherichia coli adhesion and biofilm formation ability. Rom. Biotechnol. Lett. 2017, 22, 12483–12491. Available online: http://gateway.webofknowledge.com/gateway/Gateway.cgi?GWVersion=2&SrcAuth=ORCID&SrcApp=OrcidOrg&DestLinkType=FullRecord&DestApp=WOS_CPL&KeyUT=WOS:000403059300021&KeyUID=WOS:000403059300021 (accessed on 22 March 2022).
- Yang, J.-N.; Wang, C.; Guo, C.; Peng, X.-X.; Li, H. Outer membrane proteome and its regulation networks in response to glucose concentration changes in Escherichia coli. Mol. Biosyst. 2011, 7, 3087–3093. [Google Scholar] [CrossRef]
- Zeraik, A.E.; Nitschke, M. Influence of growth media and temperature on bacterial adhesion to polystyrene surfaces. Braz. Arch. Biol. Technol. 2012, 55, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Adler, J.; Templeton, B. The Effect of Environmental Conditions on the Motility of Escherichia coli. J. Gen. Microbiol. 1967, 46, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Holst, J.J.; Gribble, F.; Horowitz, M.; Rayner, C.K. Roles of the Gut in Glucose Homeostasis. Diabetes Care 2016, 39, 884–892. [Google Scholar] [CrossRef] [Green Version]
- Anhê, F.F.; Barra, N.G.; Schertzer, J.D. Glucose alters the symbiotic relationships between gut microbiota and host physiology. Am. J. Physiol. Metab. 2020, 318, E111–E116. [Google Scholar] [CrossRef]
- Madrigal, J.L.; Bhar, S.; Hackett, S.; Engelken, H.; Joseph, R.; Keyhani, N.O.; Jones, M.K. Attach Me If You Can: Murine Norovirus Binds to Commensal Bacteria and Fungi. Viruses 2020, 12, 759. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-H.; Hsu, C.-C.; Yang, C.-D.; Ju, Y.-W.; Chen, Y.-P.; Tseng, C.-P. Negative Effect of Glucose on ompA mRNA Stability: A Potential Role of Cyclic AMP in the Repression of hfq in Escherichia coli. J. Bacteriol. 2011, 193, 5833–5840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Huang, Q.; Cui, A.; Liu, X.; Hou, B.; Zhang, L.; Liu, M.; Meng, X.; Li, S. Overexpression of Outer Membrane Protein X (OmpX) Compensates for the Effect of TolC Inactivation on Biofilm Formation and Curli Production in Extraintestinal Pathogenic Escherichia coli (ExPEC). Front. Cell. Infect. Microbiol. 2018, 8, 208. [Google Scholar] [CrossRef] [PubMed]
- Rossi, E.; Paroni, M.; Landini, P. Biofilm and motility in response to environmental and host-related signals in Gram negative opportunistic pathogens. J. Appl. Microbiol. 2018, 125, 1587–1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janderová, B.; Kapralek, F.; Julak, J. Effect of glucose on the biochemical properties of the bacterial cytoplasmic membrane. Folia Microbiol. 1980, 25, 457–463. [Google Scholar] [CrossRef]
- Zhu, L.; Liu, X.-K.; Zhao, G.; Zhi, Y.-D.; Bu, X.; Ying, T.-Y.; Feng, E.-L.; Wang, J.; Zhang, X.-M.; Huang, P.-T.; et al. Dynamic Proteome Changes of Shigella flexneri 2a During Transition from Exponential Growth to Stationary Phase. Genom. Proteom. Bioinform. 2007, 5, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Naves, P.; del Prado, G.; Huelves, L.; Gracia, M.; Ruiz, V.; Blanco, J.; Rodríguez-Cerrato, V.; Ponte, M.; Soriano, F. Measurement of biofilm formation by clinical isolates of Escherichia coliis method-dependent. J. Appl. Microbiol. 2008, 105, 585–590. [Google Scholar] [CrossRef]
- Tao, H.; Bausch, C.; Richmond, C.; Blattner, F.R.; Conway, T. Functional Genomics: Expression Analysis of Escherichia coli Growing on Minimal and Rich Media. J. Bacteriol. 1999, 181, 6425–6440. [Google Scholar] [CrossRef] [Green Version]
- Kubota, H.; Senda, S.; Nomura, N.; Tokuda, H.; Uchiyama, H. Biofilm Formation by Lactic Acid Bacteria and Resistance to Environmental Stress. J. Biosci. Bioeng. 2008, 106, 381–386. [Google Scholar] [CrossRef]
- Makinoshima, H.; Aizawa, S.-I.; Hayashi, H.; Miki, T.; Nishimura, A.; Ishihama, A. Growth Phase-Coupled Alterations in Cell Structure and Function of Escherichia coli. J. Bacteriol. 2003, 185, 1338–1345. [Google Scholar] [CrossRef] [Green Version]
- Mañas, P.; Mackey, B.M. Morphological and Physiological Changes Induced by High Hydrostatic Pressure in Exponential- and Stationary-Phase Cells of Escherichia coli: Relationship with Cell Death. Appl. Environ. Microbiol. 2004, 70, 1545–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Wood, T.K. OmpA influences Escherichia coli biofilm formation by repressing cellulose production through the CpxRA two-component system. Environ. Microbiol. 2009, 11, 2735–2746. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.L.; Desa, N.; Hansen, E.E.; Knirel, Y.A.; Gordon, J.I.; Gagneux, P.; Nizet, V.; Varki, A. Innovations in host and microbial sialic acid biosynthesis revealed by phylogenomic prediction of nonulosonic acid structure. Proc. Natl. Acad. Sci. USA 2009, 106, 13552–13557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Park, Y.-H.; Lee, C.-R.; Kim, Y.-R.; Seok, Y.-J. Glucose induces delocalization of a flagellar biosynthesis protein from the flagellated pole. Mol. Microbiol. 2016, 101, 795–808. [Google Scholar] [CrossRef]
- El-Kazzaz, W.; Morita, T.; Tagami, H.; Inada, T.; Aiba, H. Metabolic block at early stages of the glycolytic pathway activates the Rcs phosphorelay system via increased synthesis of dTDP-glucose in Escherichia coli. Mol. Microbiol. 2004, 51, 1117–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taube, S.; Perry, J.W.; Yetming, K.; Patel, S.P.; Auble, H.; Shu, L.; Nawar, H.F.; Lee, C.H.; Connell, T.D.; Shayman, J.A.; et al. Ganglioside-Linked Terminal Sialic Acid Moieties on Murine Macrophages Function as Attachment Receptors for Murine Noroviruses. J. Virol. 2009, 83, 4092–4101. [Google Scholar] [CrossRef] [Green Version]
- Carlin, A.F.; Uchiyama, S.; Chang, Y.-C.; Lewis, A.L.; Nizet, V.; Varki, A. Molecular mimicry of host sialylated glycans allows a bacterial pathogen to engage neutrophil Siglec-9 and dampen the innate immune response. Blood 2009, 113, 3333–3336. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Protein Name | Function |
|---|---|---|
| ompA | Outer membrane protein A | Outer membrane protein; receptor, adhesin, and biofilm formation |
| ompX | Outer membrane protein X | Outer membrane protein; adhesion, cell defense |
| flgB | Flagellar basal body rod protein | Structural component of flagellum |
| ftsL | Cell division protein | Role in cell division |
| rcsC | Sensor histidine kinase | Sensor kinase; biofilm formation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, K.J.; Mosby, C.A.; Jones, M.K. Glucose Reduces Norovirus Binding to Enterobacter cloacae and Alters Gene Expression of Bacterial Surface Structures in a Growth Phase Dependent Manner. Viruses 2022, 14, 1596. https://doi.org/10.3390/v14081596
Long KJ, Mosby CA, Jones MK. Glucose Reduces Norovirus Binding to Enterobacter cloacae and Alters Gene Expression of Bacterial Surface Structures in a Growth Phase Dependent Manner. Viruses. 2022; 14(8):1596. https://doi.org/10.3390/v14081596
Chicago/Turabian StyleLong, Kendall J., Chanel A. Mosby, and Melissa K. Jones. 2022. "Glucose Reduces Norovirus Binding to Enterobacter cloacae and Alters Gene Expression of Bacterial Surface Structures in a Growth Phase Dependent Manner" Viruses 14, no. 8: 1596. https://doi.org/10.3390/v14081596
APA StyleLong, K. J., Mosby, C. A., & Jones, M. K. (2022). Glucose Reduces Norovirus Binding to Enterobacter cloacae and Alters Gene Expression of Bacterial Surface Structures in a Growth Phase Dependent Manner. Viruses, 14(8), 1596. https://doi.org/10.3390/v14081596