
Sandfly-Borne Phleboviruses in Portugal: Four and Still Counting


Abstract
:1. Introduction
2. Sicilian phlebovirus
3. Toscana phlebovirus
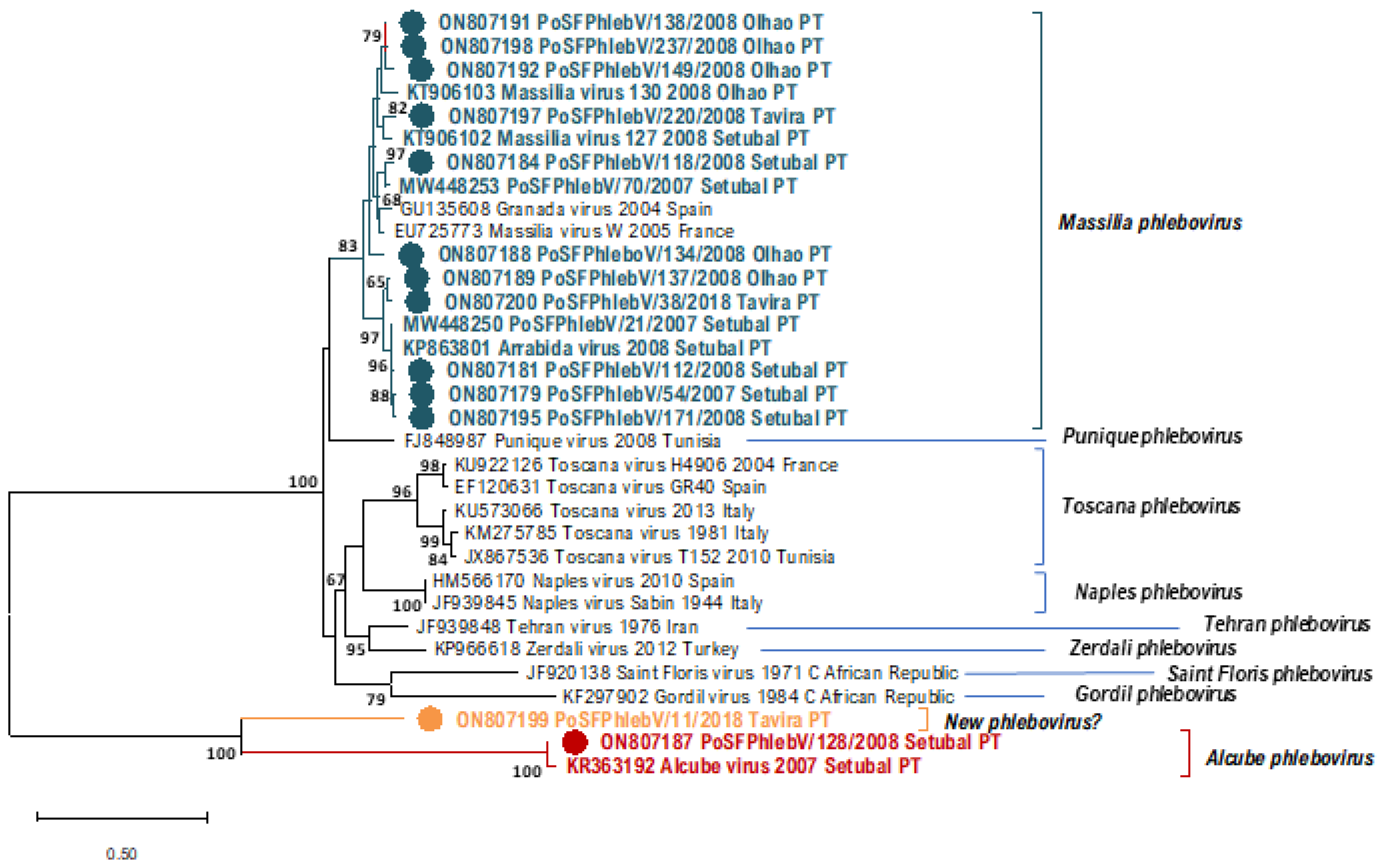
4. Massilia phlebovirus
5. Alcube phlebovirus
6. Other Genome Detections of Phleboviruses in Sandfly Pools in Portugal
7. Discussion
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Committee on Taxonomy of Viruses. Virus Taxonomy: 2021 Release. EC 53, Online, July 2021. Email Ratification March 2022 (MSL #37). Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 5 April 2022).
- Bouloy, M. Molecular Biology of Phleboviruses. In Bunyaviridae Molecular and Cellular Biology; Plyusnin, A., Elliott, R.M., Eds.; Caister Academic Press: Norfolk, UK, 2011; pp. 95–128. [Google Scholar]
- International Committee on Taxonomy of Viruses. Virus Taxonomy Release: 2020 Release. Email Ratification March 2021 (MSL #36). Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 30 April 2022).
- Amaro, F.; Zé-Zé, L.; Lourenço, J.; Giovanetti, M.; Becker, S.C.; Alves, M.J. Phylogenetic analysis of Massilia phlebovirus in Portugal. Viruses 2021, 13, 1412. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Calzolari, M. Taxonomy of phleboviruses, emphasizing those that are sandfly-borne. Viruses 2021, 13, 918. [Google Scholar] [CrossRef]
- Killick-Kendrick, R. The biology and control of phlebotomine sand flies. Clin. Dermatol. 1999, 17, 279–289. [Google Scholar] [CrossRef]
- World Health Organization. Leishmaniasis. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 13 June 2022).
- Centers for Disease Prevention and Control. Bartonella bacilliformis Infection. Available online: https://www.cdc.gov/bartonella/bartonella-bacilliformis/index.html (accessed on 25 May 2022).
- Ayhan, N.; Charrel, R.N. Sandfly-Borne Viruses of Demonstrated/Relevant Medical Importance. In Vectors and Vector-Borne Zoonotic Diseases; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Depaquit, J.; Grandadam, M.; Fouque, F.; Andry, P.E.; Peyrefitte, C. Arthropod-borne viruses transmitted by Phlebotomine sandflies in Europe: A review. Euro Surveill. 2010, 15, 19507. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, J.H.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Amarasingh, G.K.; Anthony, S.J.; Avšič-Županc, T.; Ayllón, M.A.; Bahl, J.; Balkema-Buschmann, A.; et al. 2020 taxonomic update for phylum Negarnaviricota (Riboviria: Orthornavirae), including the large orders Bunyavirales and Mononegavirales. Arch. Virol. 2020, 165, 3023–3072. [Google Scholar] [CrossRef] [PubMed]
- Moureau, G.; Bichaud, L.; Salez, N.; Ninove, L.; Hamrioui, B.; Belazzoug, S.; de Lamballerie, X.; Izri, A.; Charrel, R.N. Molecular and serological evidence for the presence of novel phleboviruses in sandflies from northern Algeria. Open. Virol. J. 2010, 4, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Alkan, C.; Bichaud, L.; de Lamballerie, X.; Alten, B.; Gould, E.A.; Charrel, R.N. Sandfly-borne phleboviruses of Eurasia and Africa: Epidemiology, genetic diversity, geographic range, control measures. Antiv. Res. 2013, 100, 54–74. [Google Scholar] [CrossRef]
- Izri, A.; Temmam, S.; Moureau, G.; Hamrioui, B.; de Lamballerie, X.; Charrel, R.N. Sandfly fever Sicilian virus, Algeria. Emerg. Infect. Dis. 2008, 14, 795–797. [Google Scholar] [CrossRef]
- Woyessa, A.B.; Omballa, V.; Wang, D.; Lambert, A.; Waiboci, L.; Ayele, W.; Ahmed, A.; Abera, N.A.; Cao, S.; Ochieng, M.; et al. An outbreak of acute febrile illness caused by Sandfly Fever Sicilian Virus in the Afar region of Ethiopia, 2011. Am. J. Trop. Med. Hyg. 2014, 91, 1250–1253. [Google Scholar] [CrossRef]
- Ayhan, N.; López-Roig, M.; Monastiri, A.; Charrel, R.N.; Serra-Cobo, J. Seroprevalence of Toscana Virus and Sandfly Fever Sicilian Virus in European bat colonies measured using a neutralization test. Viruses 2021, 13, 88. [Google Scholar] [CrossRef]
- Becker, M.; Zielen, S.; Schwarz, T.F.; Linde, R.; Hofmann, D. Pappataci-Fieber [Pappataci fever]. Klin. Padiatr. 1997, 209, 377–379. (In German) [Google Scholar] [CrossRef] [PubMed]
- Lesho, E.P.; Ludwig, G.V.; Wortmann, G. Encephalitis and Sandfly Fever (Sicilian) virus infection. Infect. Dis. Clin. Pract. 2004, 12, 352–354. [Google Scholar] [CrossRef]
- Selim, H.S.; El-Barrawy, M.A.; Rakha, M.E.; Yingst, S.L.; Baskharoun, M.F. Microbial study of meningitis and encephalitis cases. J. Egypt. Public Health Assoc. 2007, 82, 1–19. [Google Scholar]
- Ergunay, K.; Ismayilova, V.; Colpak, I.A.; Kansu, T.; Us, D. A case of central nervous system infection due to a novel Sandfly Fever Virus (SFV) variant: Sandfly Fever Turkey Virus (SFTV). J. Clin. Virol. 2012, 54, 79–82. [Google Scholar] [CrossRef]
- Temocin, F.; Sari, T.; Tulek, N. Sandfly fever with skin lesions: A case series from Turkey. J. Arthropod Borne Dis. 2016, 10, 608–612. [Google Scholar]
- Niklasson, B.; Eitrem, R. Sandfly fever among Swedish UN troops in Cyprus. Lancet 1985, 1, 1212. [Google Scholar] [CrossRef]
- Papa, A.; Konstantinou, G.; Pavlidou, V.; Antoniadis, A. Sandfly fever virus outbreak in Cyprus. Clin. Microbiol. Infect. 2006, 12, 192–194. [Google Scholar] [CrossRef] [PubMed]
- Eitrem, R.; Niklasson, B.; Weiland, O. Sandfly fever among Swedish tourists. Scand. J. Infect. Dis. 1991, 23, 451–457. [Google Scholar] [CrossRef]
- Carhan, A.; Uyar, Y.; Ozkaya, E.; Ertek, M.; Dobler, G.; Dilcher, M.; Wang, Y.; Spiegel, M.; Hufert, F.; Weidmann, M. Characterization of a sandfly fever Sicilian virus isolated during a sandfly fever epidemic in Turkey. J. Clin. Virol. 2010, 48, 264–269. [Google Scholar] [CrossRef]
- Guler, S.; Guler, E.; Caglayik, D.Y.; Kokoglu, O.F.; Ucmak, H.; Bayrakdar, F.; Uyar, Y. A sandfly fever virus outbreak in the East Mediterranean region of Turkey. Int. J. Infect. Dis. 2012, 16, e244–e246. [Google Scholar] [CrossRef]
- Torun, E.C.; Yağçı, Ç.D.; Uyar, Y.; Korukluoğlu, G.; Ertek, M.K. Sandfly fever outbreak in a province at Central Anatolia, Turkey. Mikrobiyol. Bul. 2010, 44, 431–439. [Google Scholar]
- Tezcan, S.; Dinçer, E.; Ülger, M.; Özgür, D.; Erdoğan, S.; Özkul, A.; Emekdaş, G.; Ergünay, K. Serological investigation of phlebovirus exposure in blood donors from the Mediterranean Province of Mersin, Turkey. Mikrobiyol. Bul. 2015, 49, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Ellis, S.B.; Appenzeller, G.; Lee, H.; Mullen, K.; Swenness, R.; Pimentel, G.; Mohareb, E.; Warner, C. Outbreak of sandfly fever in central Iraq, September 2007. Mil. Med. 2008, 173, 949–953. [Google Scholar] [CrossRef]
- Riddle, M.S.; Althoff, J.M.; Earhart, K.; Monteville, M.R.; Yingst, S.L.; Mohareb, E.W.; Putnam, S.D.; Sanders, J.W. Serological evidence of arboviral infection and self-reported febrile illness among U.S. troops deployed to Al Asad, Iraq. Epidemiol. Infect. 2008, 136, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Bichaud, L.; Piarroux, R.P.; Izri, A.; Ninove, L.; Mary, C.; De Lamballerie, X.; Charrel, R.N. Low seroprevalence of sandfly fever Sicilian virus antibodies in humans, Marseille, France. Clin. Microbiol. Infect. 2011, 17, 1189–1190. [Google Scholar] [CrossRef]
- Calamusa, G.; Valenti, R.M.; Vitale, F.; Mammina, C.; Romano, N.; Goedert, J.J.; Gori-Savellini, G.; Cusi, M.G.; Amodio, E. Seroprevalence of and risk factors for Toscana and Sicilian virus infection in a sample population of Sicily (Italy). J. Infect. 2012, 64, 212–217. [Google Scholar] [CrossRef]
- Marchi, S.; Trombetta, C.M.; Kistner, O.; Montomoli, E. Seroprevalence study of Toscana virus and viruses belonging to the Sandfly fever Naples antigenic complex in central and southern Italy. J. Infect. Public Health 2017, 10, 866–869. [Google Scholar] [CrossRef]
- Percivalle, E.; Cassaniti, I.; Calzolari, M.; Lelli, D.; Baldanti, F. Thirteen years of phleboviruses circulation in Lombardy, a Northern Italy region. Viruses 2021, 13, 209. [Google Scholar] [CrossRef] [PubMed]
- Tesh, R.B.; Papaevangelou, G. Effect of insecticide spraying for malaria control on the incidence of sandfly fever in Athens, Greece. Am. J. Trop. Med. Hyg. 1977, 26, 163–166. [Google Scholar] [CrossRef]
- Antoniadis, A.; Alexiou Daniel, S.; Malissiovas, N.; Doutsos, J.; Polyzoni, T.; LeDuc, J.W.; Peters, C.J.; Saviolakis, G. Seroepidemiological Survey for Antibodies to Arboviruses in Greece; In Hemorrhagic Fever with Renal Syndrome, Tick-and Mosquito-Borne Viruses; Springer: Vienna, Austria, 1990; pp. 277–285. [Google Scholar]
- Venturi, G.; Marchi, A.; Fiorentini, C.; Ramadani, N.; Quaglio, G.; Kalaveshi, A.; Bertinato, L.; Putoto, G.; Benedetti, E.; Rezza, G.; et al. Prevalence of antibodies to phleboviruses and flaviviruses in Peja, Kosovo. Clin. Microbiol. Infect. 2011, 17, 1180–1182. [Google Scholar] [CrossRef]
- Eitrem, R.; Stylianou, M.; Niklasson, B. High prevalence rates of antibody to three sandfly fever viruses (Sicilian, Naples, and Toscana) among Cypriots. Epidemiol. Infect. 1991, 107, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Billioud, G.; Tryfonos, C.; Richter, J. The Prevalence of Antibodies against Sandfly Fever Viruses and West Nile Virus in Cyprus. J. Arthropod. Borne. Dis. 2019, 13, 116–125, Published 30 March 2019. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.C.; Haberberger, R.L.; Salib, A.W.; Soliman, B.A.; El-Tigani, A.; Khalid, I.O.; Watts, D.M. Evaluation of arthropod-borne viruses and other infectious disease pathogens as the causes of febrile illnesses in the Khartoum Province of Sudan. J. Med. Virol. 1996, 48, 141–146. [Google Scholar] [CrossRef]
- Darwish, M.A.; Feinsod, F.M.; Scott, R.M.N.; Ksiazek, T.G.; Botros, B.A.M.; Farrag, I.H.; Said, S. Arboviral causes of non-specific fever and myalgia in a fever hospital patient population in Cairo, Egypt. Trans. R. Soc. Trop. Med. Hyg. 1987, 87, 1001–1003. [Google Scholar] [CrossRef]
- Saidi, S.; Tesh, R.B.; Javadian, E.; Sahabi, Z.; Nadim, A. Studies on the epidemiology of sandfly fever in Iran. II. The prevalence of human and animal infection with five phlebotomus fever virus serotypes in Isfahan province. Am. J. Trop. Med. Hyg. 1977, 26, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Shiraly, R.; Khosravi, A.; Farahangiz, S. Seroprevalence of sandfly fever virus infection in military personnel on the western border of Iran. J. Infect. Public Health 2017, 10, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Cohen, D.; Zaide, Y.; Karasenty, E.; Schwarz, M.; LeDuc, J.W.; Slepon, R.; Ksiazek, T.G.; Shemer, J.; Green, M.S. Prevalence of antibodies to West Nile fever, sandfly fever Sicilian, and sandfly fever Naples viruses in healthy adults in Israel. Public Health Rev. 1999, 27, 217–230. [Google Scholar]
- Darwish, M.A.; Hoogstraal, H.; Roberts, T.J.; Ghazi, R.; Amer, T. A seroepidemiological survey for Bunyaviridae and certain other arboviruses in Pakistan. Trans. R. Soc. Trop. Med. Hyg. 1983, 77, 446–450. [Google Scholar] [CrossRef]
- Gaidamovich, S.Y.; Baten, M.A.; Klisenko, G.A.; Melnikova, Y.E. Serological studies on sandfly fevers in the Republic of Bangladesh. Acta Virol. 1984, 28, 325–328. [Google Scholar]
- Nikolaev, V.P.; Perepelkin, V.S.; Raevskiĭ, K.K.; Prusakova, Z.M. [A natural focus of sandfly fever in the Republic of Afghanistan]. Zh. Mikrobiol. Epidemiol. Immunobiol. 1991, 3, 39–41. [Google Scholar]
- Tesh, R.B.; Saidi, S.; Gajdamovic, S.J.; Rodhain, F.; Vesenjak-Hirjan, J. Serological studies on the epidemiology of sandfly fever in the Old World. Bull. World Health Organ. 1976, 54, 663–674. [Google Scholar] [PubMed]
- Chaste, L.C.; Launay, H.; Bailly-Choumara, H.; Le Lay, G.; Beaucournu, J.C. Infections à arbovirus au Maroc: Sondage sérologique chez les petits mammifères du nord du pays [Arbovirus infections in Morocco: Serosurvey in small wild mammals in the northern part of the country]. Bull. Soc. Pathol. Exot. Filiales 1982, 75, 466–475. [Google Scholar]
- Chastel, C.; Bach-Hamba, D.; Launay, H.; Le Lay, G.; Hellal, H.; Beaucournu, J.C. Infections à arbovirus en Tunisie: Nouvelle enquête sérologique chez les petits mammifères sauvages [Arbovirus infections in Tunisia: New serological survey of small wild mammals]. Bull. Soc. Pathol. Exot. Filiales 1983, 76, 21–33. [Google Scholar] [PubMed]
- Chastel, C.; Launay, H.; Rogues, G.; Beaucournu, J.C. Infections à arbovirus en Espagne: Enquête sérologique chez les petits mammifères [Arbovirus infections in Spain: Serological survey on small mammals]. Bull. Soc. Pathol. Exot. Filiales 1980, 73, 384–390. [Google Scholar] [PubMed]
- Le Lay-Roguès, G.; Valle, M.; Chastel, C.; Beaucournu, J.C. Petits mammifères sauvages et arbovirus en Italie [Small wild mammals and arboviruses in Italy]. Bull Soc Pathol Exot Filiales 1983, 76, 333–345. [Google Scholar] [PubMed]
- Alwassouf, S.; Christodoulou, V.; Bichaud, L.; Ntais, P.; Mazeris, A.; Antoniou, M.; Charrel, R.N. Seroprevalence of Sandfly-Borne Phleboviruses Belonging to Three Serocomplexes (Sandfly fever Naples, Sandfly fever Sicilian and Salehabad) in Dogs from Greece and Cyprus Using Neutralization Test. PLoS Negl. Trop. Dis. 2016, 10, e0005063. [Google Scholar] [CrossRef] [PubMed]
- Sakhria, S.; Alwassouf, S.; Fares, W.; Bichaud, L.; Dachraoui, K.; Alkan, C.; Zoghlami, Z.; de Lamballerie, X.; Zhioua, E.; Charrel, R.N. Presence of sandfly-borne phleboviruses of two antigenic complexes (Sandfly fever Naples virus and Sandfly fever Sicilian virus) in two different bio-geographical regions of Tunisia demonstrated by a microneutralisation-based seroprevalence study in dogs. Parasit. Vectors 2014, 7, 476. [Google Scholar] [CrossRef]
- Ayhan, N.; Sherifi, K.; Taraku, A.; Bërxholi, K.; Charrel, R.N. High Rates of Neutralizing Antibodies to Toscana and Sandfly Fever Sicilian Viruses in Livestock, Kosovo. Emerg. Infect. Dis. 2017, 23, 989–992. [Google Scholar] [CrossRef]
- Filipe, A.R. Serological survey for antibodies to arboviruses in the human population of Portugal. Trans. R. Soc. Trop. Med. Hyg. 1974, 68, 311–314. [Google Scholar] [CrossRef]
- Guerra, A.B.; Gouveia, C.; Zé, L.; Amaro, F.; Ferreira, G.C.; Brito, M.J. Prolonged Febrile Illness Caused by Sicilian Virus Infection in Portugal. In Proceedings of the 36th Annual meeting of the European Society for Paediatric Infectious Diseases, Malmo, Sweden, 28 May–2 June 2018. [Google Scholar]
- Amaro, F.; Zé-Zé, L.; Luz, M.T.; Alves, M.J. Toscana Virus: Ten Years of Diagnostics in Portugal. Acta Med. Port. 2021, 34, 677–681. [Google Scholar] [CrossRef]
- Maia, C.; Ayhan, N.; Cristovão, J.M.; Pereira, A.; Charrel, R. Human seroprevalence of Toscana virus and Sicilian phlebovirus in the southwest of Portugal. Eur. J. Clin. Microbio.l Infect. Dis. 2022, 41, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Alwassouf, S.; Maia, C.; Ayhan, N.; Coimbra, M.; Cristovão, J.M.; Richet, H.; Bichaud, L.; Campino, L.; Charrel, R.N. Neutralization-based seroprevalence of Toscana virus and sandfly fever Sicilian virus in dogs and cats from Portugal. J. Gen. Virol. 2016, 97, 2816–2823. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.; Ayhan, N.; Cristóvão, J.M.; Vilhena, H.; Martins, Â.; Cachola, P.; Henriques, J.; Coimbra, M.; Catarino, A.; Lestinova, T.; et al. Antibody response to Toscana Virus and Sandfly Fever Sicilian Virus in cats naturally exposed to phlebotomine Sand Fly bites in Portugal. Microorganisms 2019, 7, 339. [Google Scholar] [CrossRef]
- Maia, C.; Alwassouf, S.; Cristóvão, J.M.; Ayhan, N.; Pereira, A.; Charrel, R.N.; Campino, L. Serological association between Leishmania infantum and sand fly fever Sicilian (but not Toscana) virus in sheltered dogs from southern Portugal. Parasit. Vectors 2017, 10, 92. [Google Scholar] [CrossRef]
- Verani, P.; Ciufolini, M.G.; Caciolli, S.; Renzi, A.; Nicoletti, L.; Sabatinelli, G.; Bartolozzi, D.; Volpi, G.K.; Amaducci, L.; Coluzzi, M. Ecology of viruses isolated from sand flies in Italy and characterized of a new Phlebovirus (Arbia virus). Am. J. Trop. Med. Hyg. 1988, 38, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Leoncini, F.; Bartolozzi, D.; Banchi, S. Il virus Toscana: Un nuovo Phlebovirus causa di malattie infiammatorie acute del SNC nell’uomo. Giorn. Mal. Inf. Parass. 1986, 38, 649–652. [Google Scholar]
- Charrel, R.N.; Gallian, P.; Navarro-Mari, J.M.; Nicoletti, L.; Papa, A.; Sánchez-Seco, M.P.; Tenorio, A.; de Lamballerie, X. Emergence of Toscana virus in Europe. Emer. Infec. Dis. 2005, 11, 1657–1663. [Google Scholar] [CrossRef]
- Charrel, R.N.; Bichaud, L.; de Lamballerie, X. Emergence of Toscana virus in the mediterranean area. World J. Virol. 2012, 1, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Laroche, L.; Jourdain, F.; Ayhan, N.; Bañuls, A.L.; Charrel, R.; Prudhomme, J. Incubation Period for Neuroinvasive Toscana Virus Infections. Emerg. Infect. Dis. 2021, 27, 3147–3150. [Google Scholar] [CrossRef]
- Pierro, A.; Ficarelli, S.; Ayhan, N.; Morini, S.; Raumer, L.; Bartoletti, M.; Mastroianni, A.; Prati, F.; Schivazappa, S.; Cenni, P.; et al. Characterization of antibody response in neuroinvasive infection caused by Toscana virus. Clin. Microbiol. Infect. 2017, 23, 868–873. [Google Scholar] [CrossRef]
- Ergunay, K.; Kaplan, B.; Okar, S.; Akkutay-Yoldar, Z.; Kurne, A.; Arsava, E.M.; Ozkul, A. Urinary detection of toscana virus nucleic acids in neuroinvasive infections. J. Clin. Virol. 2015, 70, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Matusali, G.; D’Abramo, A.; Terrosi, C.; Carletti, F.; Colavita, F.; Vairo, F.; Savellini, G.G.; Gandolfo, C.; Anichini, G.; Lalle, E.; et al. Infectious Toscana Virus in Seminal Fluid of Young Man Returning from Elba Island, Italy. Emer. Infect. Dis. 2022, 28, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Arboviral Diseases, Neuroinvasive and Non-Neuroinvasive. 2015 Case Definition. Available online: https://ndc.services.cdc.gov/case-definitions/arboviral-diseases-neuroinvasive-and-non-neuroinvasive-2015/ (accessed on 8 June 2022).
- Bartels, S.; de Boni, L.; Kretzschmar, H.A.; Heckmann, J.G. Lethal encephalitis caused by the Toscana virus in an elderly patient. J. Neurol. 2012, 259, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Popescu, C.P.; Cotar, A.I.; Dinu, S.; Zaharia, M.; Tardei, G.; Ceausu, E.; Badescu, D.; Ruta, S.; Ceianu, C.S.; Florescu, S.A. Emergence of Toscana Virus, Romania, 2017–2018. Emerg. Infect. Dis. 2021, 27, 1482–1485. [Google Scholar] [CrossRef]
- Rota, E.; Morelli, N.; Immovilli, P.; De Mitri, P.; Guidetti, D. Guillain-Barré-like axonal polyneuropathy associated with Toscana virus infection: A case report. Medicine (Baltimore). 2017, 96, e8081. [Google Scholar] [CrossRef]
- Okar, S.V.; Bekircan-Kurt, C.E.; Hacıoğlu, S.; Erdem-Özdamar, S.; Özkul, A.; Ergünay, K. Toscana virus associated with Guillain-Barré syndrome: A case-control study. Acta Neurol. Belg. 2021, 121, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Baldelli, F.; Ciufolini, M.G.; Francisci, D.; Marchi, A.; Venturi, G.; Fiorentini, C.; Luchetta, M.L.; Bruto, L.; Pauluzzi, S. Unusual presentation of life-threatening Toscana virus meningoencephalitis. Clin. Infect. Dis. 2004, 38, 515–520. [Google Scholar] [CrossRef]
- Oechtering, J.; Petzold, G.C. Acute hydrocephalus due to impaired CSF resorption in Toscana virus meningoencephalitis. Neurology 2012, 79, 829–831. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, F.A.; Moreno-Docón, A.; Segovia-Hernández, M.; Fernández-Barreiro, A. Sordera como secuela de meningitis por el virus Toscana [Deafness as a sequela of Toscana virus meningitis]. Med. Clin. 2008, 130, 639. (In Spanish) [Google Scholar] [CrossRef]
- Sanbonmatsu-Gámez, S.; Pérez-Ruiz, M.; Palop-Borrás, B.; Navarro-Marí, J.M. Unusual manifestation of toscana virus infection, Spain. Emerg. Infect. Dis. 2009, 15, 347–348. [Google Scholar] [CrossRef]
- Mosnier, E.; Charrel, R.; Vidal, B.; Ninove, L.; Schleinitz, N.; Harlé, J.R.; Bernit, E. Toscana virus myositis and fasciitis. Med. Mal. Infect. 2013, 43, 208–210. [Google Scholar] [CrossRef]
- Marlinge, M.; Crespy, L.; Zandotti, C.; Piorkowski, G.; Kaphan, E.; Charrel, R.N.; Ninove, L. Afebrile meningoencephalitis with transient central facial paralysis due to Toscana virus infection, southeastern France, 2014 [corrected]. Euro Surveill. Bull. Eur. Sur Les Mal. Transm. Eur. Commun. Dis. Bull. 2014, 19, 20974. [Google Scholar] [CrossRef]
- Serata, D.; Rapinesi, C.; Del Casale, A.; Simonetti, A.; Mazzarini, L.; Ambrosi, E.; Kotzalidis, G.D.; Fensore, C.; Girardi, P.; Tatarelli, R. Personality changes after Toscana virus (TOSV) encephalitis in a 49-year-old man: A case report. Intern. J. Neurosc. 2011, 121, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Zanelli, G.; Bianco, C.; Cusi, M.G. Testicular involvement during Toscana virus infection: An unusual manifestation? Infection 2013, 41, 735–736. [Google Scholar] [CrossRef] [PubMed]
- Mascitti, H.; Calin, R.; Dinh, A.; Makhloufi, S.; Davido, B. Testicular pain associated with clear fluid meningitis: How many cases of Toscana virus are we missing? Int. J. Infect. Dis. 2020, 93, 198–200. [Google Scholar] [CrossRef]
- Ayhan, N.; Charrel, R.N. An update on Toscana virus distribution, genetics, medical and diagnostic aspects. Clin. Microbiol Infect. 2020, 26, 1017–1023. [Google Scholar] [CrossRef]
- Sun, M.H.; Ji, Y.F.; Li, G.H.; Shao, J.W.; Chen, R.X.; Gong, H.Y.; Chen, S.Y.; Chen, J.M. Highly adaptive Phenuiviridae with biomedical importance in multiple fields. J. Med. Virol. 2022, 94, 2388–2401. [Google Scholar] [CrossRef]
- Sakhria, S.; Bichaud, L.; Mensi, M.; Salez, N.; Dachraoui, K.; Thirion, L.; Cherni, S.; Chelbi, I.; De Lamballerie, X.; Zhioua, E.; et al. Co-circulation of Toscana virus and Punique virus in northern Tunisia: A microneutralisation-based seroprevalence study. PLoS Negl. Trop. Dis. 2013, 7, e2429. [Google Scholar] [CrossRef]
- Valassina, M.; Valentini, M.; Pugliese, A.; Valensin, P.E.; Cusi, M.G. Serological survey of Toscana virus infections in a high-risk population in Italy. Clin. Diagn. Lab. Immunol. 2003, 10, 483–484. [Google Scholar] [CrossRef]
- Punda-Polić, V.; Jerončić, A.; Mohar, B.; Šiško Kraljević, K. Prevalence of Toscana virus antibodies in residents of Croatia. Clin. Microbiol. Infect. 2012, 18, E200–E203. [Google Scholar] [CrossRef]
- Tesh, R.B.; Lubroth, J.; Guzman, H. Simulation of arbovirus overwintering: Survival of Toscana virus (Bunyaviridae:Phlebovirus) in its natural sand fly vector Phlebotomus perniciosus. Am. J. Trop. Med. Hyg. 1992, 47, 574–581. [Google Scholar] [CrossRef]
- Ciufolini, M.G.; Maroli, M.; Guandalini, E.; Marchi, A.; Verani, P. Experimental studies on the maintenance of Toscana and Arbia viruses (Bunyaviridae: Phlebovirus). Am. J. Trop. Med. Hyg. 1989, 40, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Tesh, R.B.; Modi, G.B. Maintenance of Toscana virus in Phlebotomus perniciosus by vertical transmission. Am. J. Trop. Med. Hyg. 1987, 36, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Bichaud, L.; Dachraoui, K.; Piorkowski, G.; Chelbi, I.; Moureau, G.; Cherni, S.; De Lamballerie, X.; Sakhria, S.; Charrel, R.N.; Zhioua, E. Toscana virus isolated from sandflies, Tunisia. Emerg. Infect. Dis. 2013, 19, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Hacioglu, S.; Dincer, E.; Isler, C.T.; Karapinar, Z.; Ataseven, V.S.; Ozkul, A.; Ergunay, K. A Snapshot Avian Surveillance Reveals West Nile Virus and Evidence of Wild Birds Participating in Toscana Virus Circulation. Vector Borne Zoonotic Dis. 2017, 17, 698–708. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Marí, J.M.; Palop-Borrás, B.; Pérez-Ruiz, M.; Sanbonmatsu-Gámez, S. Serosurvey study of Toscana virus in domestic animals, Granada, Spain. Vector Borne Zoonotic Dis. 2011, 11, 583–587. [Google Scholar] [CrossRef]
- Dahmani, M.; Alwassouf, S.; Grech-Angelini, S.; Marié, J.L.; Davoust, B.; Charrel, R.N. Seroprevalence of Toscana virus in dogs from Corsica, France. Parasit. Vectors 2016, 9, 381. [Google Scholar] [CrossRef]
- Tahir, D.; Alwassouf, S.; Loudahi, A.; Davoust, B.; Charrel, R.N. Seroprevalence of Toscana virus in dogs from Kabylia (Algeria). Clin. Microbiol. Infect. 2016, 22, e16–e17. [Google Scholar] [CrossRef]
- Ehrnst, A.; Peters, C.J.; Niklasson, B.; Svedmyr, A.; Holmgren, B. Neurovirulent Toscana virus (a sandfly fever virus) in Swedish man after visit to Portugal. Lancet 1985, 1, 1212–1213. [Google Scholar] [CrossRef]
- Schwarz, T.F.; Jäger, G.; Gilch, S.; Pauli, C. Serosurvey and laboratory diagnosis of imported sandfly fever virus, serotype Toscana, infection in Germany. Epidemiol. Infect. 1995, 114, 501–510. [Google Scholar] [CrossRef]
- Santos, L.; Simões, J.; Costa, R.; Martins, S.; Lecour, H. Toscana virus meningitis in Portugal, 2002-2005. Euro Surveill. 2007, 12, E3–E4. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.; Cardoso, M.J.; Marinho, A.S.; Guimarães, T.; Sarmento, A. Seroprevalence survey of Toscana virus infection in Oporto region. Acta Med. Port. 2011, 24, 479–482. (In Portuguese) [Google Scholar]
- Amaro, F.; Luz, T.; Parreira, P.; Ciufolini, M.G.; Marchi, A.; Janeiro, N.; Zagalo, A.; Proença, P.; Ramos, M.I.; Alves, M.J. Toscana virus in the Portuguese population: Serosurvey and clinical cases. Acta Med. Port. 2011, 24 (Suppl. 2), 503–508. (In Portuguese) [Google Scholar] [PubMed]
- Amaro, F. Phlebovirus, Importância em Saúde Pública em Portugal. Ph.D. Thesis, Faculty of Sciences, University of Lisbon, Lisbon, Portugal, 14 December 2010. (In Portuguese). [Google Scholar]
- Alho, A.M.; Pita, J.; Amaro, A.; Amaro, F.; Schnyder, M.; Grimm, F.; Custódio, A.C.; Cardoso, L.; Deplazes, P.; de Carvalho, L.M. Seroprevalence of vector-borne pathogens and molecular detection of Borrelia afzelii in military dogs from Portugal. Parasit. Vectors 2016, 9, 225. [Google Scholar] [CrossRef]
- Charrel, R.N.; Moureau, G.; Temmam, S.; Izri, A.; Marty, P.; Parola, P.; da Rosa, A.T.; Tesh, R.B.; de Lamballerie, X. Massilia virus, a novel Phlebovirus (Bunyaviridae) isolated from sandflies in the Mediterranean. Vector Borne Zoonotic Dis. 2009, 9, 519–530. [Google Scholar] [CrossRef]
- Sanchez-Seco, M.P.; Vazquez, A.; Collao, X.; Hernandez, L.; Aranda, C.; Ruiz, S.; Escosa, R.; Marques, E.; Bustillo, M.A.; Molero, F.; et al. Surveillance of arboviruses in Spanish wetlands: Detection of new flavi- and phleboviruses. Vector Borne Zoonotic Dis. 2010, 10, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Collao, X.; Palacios, G.; de Ory, F.; Sanbonmatsu, S.; Pérez-Ruiz, M.; Navarro, J.M.; Molina, R.; Hutchison, S.K.; Lipkin, W.I.; Tenorio, A.; et al. Granada virus: A natural phlebovirus reassortant of the sandfly fever Naples serocomplex with low seroprevalence in humans. Am. J. Trop. Med. Hyg. 2010, 83, 760–765. [Google Scholar] [CrossRef]
- Navarro-Marí, J.M.; Gómez-Camarasa, C.; Pérez-Ruiz, M.; Sanbonmatsu-Gámez, S.; Pedrosa-Corral, I.; Jiménez-Valera, M. Clinic-epidemiologic study of human infection by Granada virus, a new phlebovirus within the sandfly fever Naples serocomplex. Am. J. Trop Med. Hyg. 2013, 88, 1003–1006. [Google Scholar] [CrossRef]
- Amaro, F.; Hanke, D.; Zé-Zé, L.; Alves, M.J.; Becker, S.C.; Höper, D. Genetic characterization of Arrabida virus, a novel phlebovirus isolated in South Portugal. Virus Res. 2016, 214, 19–25. [Google Scholar] [CrossRef]
- Amaro, F.; Zé-Zé, L.; Alves, M.J.; Börstler, J.; Clos, J.; Lorenzen, S.; Becker, S.C.; Schmidt-Chanasit, J.; Cadar, D. Co-circulation of a novel phlebovirus and Massilia virus in sandflies, Portugal. Virol. J. 2015, 12, 174. [Google Scholar] [CrossRef]
- Palacios, G.; Savji, N.; Travassos da Rosa, A.; Desai, A.; Sanchez-Seco, M.P.; Guzman, H.; Lipkin, W.I.; Tesh, R. Characterization of the Salehabad virus species complex of the genus Phlebovirus (Bunyaviridae). J. Gen. Virol. 2013, 94 (Pt 4), 837–842. [Google Scholar] [CrossRef]
- Alkan, C.; Alwassouf, S.; Piorkowski, G.; Bichaud, L.; Tezcan, S.; Dincer, E.; Ergunay, K.; Ozbel, Y.; Alten, B.; de Lamballerie, X.; et al. Isolation, genetic characterization, and seroprevalence of Adana virus, a novel phlebovirus belonging to the Salehabad virus complex, in Turkey. J. Virol. 2015, 89, 4080–4091. [Google Scholar] [CrossRef] [PubMed]
- Bichaud, L.; Dachraoui, K.; Alwassouf, S.; Alkan, C.; Mensi, M.; Piorkowski, G.; Sakhria, S.; Seston, M.; Fares, W.; De Lamballerie, X.; et al. Isolation, full genomic characterization and neutralization-based human seroprevalence of Medjerda Valley virus, a novel sandfly-borne phlebovirus belonging to the Salehabad virus complex in northern Tunisia. J. Gen. Virol. 2016, 97, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Velo, E.; Bino, S. A novel phlebovirus in Albanian sandflies. Clin. Microbiol. Infect. 2011, 17, 585–587. [Google Scholar] [CrossRef] [PubMed]
- Anagnostou, V.; Pardalos, G.; Athanasiou-Metaxa, M.; Papa, A. Novel phlebovirus in febrile child, Greece. Emerg. Infect. Dis. 2011, 17, 940–941. [Google Scholar] [CrossRef]
- Lambert, A.J.; Lanciotti, R.S. Consensus amplification and novel multiplex sequencing method for S segment species identification of 47 viruses of the Orthobunyavirus, Phlebovirus, and Nairovirus genera of the family Bunyaviridae. J. Clin. Microbiol. 2009, 47, 2398–2404. [Google Scholar] [CrossRef]
- Sánchez-Seco, M.P.; Echevarría, J.M.; Hernández, L.; Estévez, D.; Navarro-Marí, J.M.; Tenorio, A. Detection and identification of Toscana and other phleboviruses by RT-nested-PCR assays with degenerated primers. J. Med. Virol. 2003, 71, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2020. [Google Scholar]
- Ayhan, N.; Charrel, R.N. Of phlebotomines (sandflies) and viruses: A comprehensive perspective on a complex situation. Curr. Opin. Insect. Sci. 2017, 22, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Ergunay, K.; Ayhan, N.; Charrel, R.N. Novel and emergent sandfly-borne phleboviruses in Asia Minor: A systematic review. Rev. Med. Virol. 2017, 27, e1898. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Weinberg, A.N.; Muth, D.J.; Lazuick, J.S. Toscana virus infection in United States citizen returning from Italy. Lancet 1987, 329, 165–166. [Google Scholar] [CrossRef]
- Sonderegger, B.; Hachler, H.; Dobler, G.; Frei, M. Imported aseptic meningitis due to Toscana virus acquired on the island of Elba, Italy, August 2008. Euro Surveill. 2009, 14, 19079. [Google Scholar] [CrossRef]
- Schultze, D.; Korte, W.; Rafeiner, P.; Niedrig, M. First report of sandfly fever virus infection imported from Malta into Switzerland, October 2011. Euro Surveill. 2012, 17, 20209. [Google Scholar] [CrossRef]
- Arden, K.E.; Heney, C.; Shaban, B.; Nimmo, G.R.; Nissen, M.D.; Sloots, T.P.; Mackay, I.M. Detection of Toscana virus from an adult traveler returning to Australia with encephalitis. J. Med. Virol. 2017, 89, 1861–1864. [Google Scholar] [CrossRef]
- Oerther, S.; Jöst, H.; Heitmann, A.; Lühken, R.; Krüger, A.; Steinhausen, I.; Brinker, C.; Lorentz, S.; Marx, M.; Schmidt-Chanasit, J.; et al. Phlebotomine sand flies in Southwest Germany: An update with records in new locations. Parasit. Vectors 2020, 13, 173. [Google Scholar] [CrossRef]
- Grimm, F.; Gessler, M.; Jenni, L. Aspects of sandfly biology in southern Switzerland. Med. Vet. Entomol. 1993, 7, 170–176. [Google Scholar] [CrossRef]
- Poeppl, W.; Obwaller, A.G.; Weiler, M.; Burgmann, H.; Mooseder, G.; Lorentz, S.; Rauchenwald, F.; Aspöck, H.; Walochnik, J.; Naucke, T.J. Emergence of sandflies (Phlebotominae) in Austria, a Central European country. Parasitol. Res. 2013, 112, 4231–4237. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Van Bortel, W.; Zeller, H.; Alten, B. A summary of the evidence for the change in European distribution of phlebotomine sand flies (Diptera: Psychodidae) of public health importance. J. Vector Ecol. 2014, 39, 72–77. [Google Scholar] [CrossRef]
- La Bella, S.; Scorrano, G.; Greco, R.; Trotta, D.; Aricò, M. Cyprus virus infection: A rare but life-threatening diagnosis in children. Pediatr. Infect. Dis. J. 2022, 41, e299–e300. [Google Scholar] [CrossRef] [PubMed]
- Sanbonmatsu-Gámez, S.; Pedrosa-Corral, I.; Navarro-Marí, J.M.; Pérez-Ruiz, M. Update in Diagnostics of Toscana Virus Infection in a Hyperendemic Region (Southern Spain). Viruses 2021, 13, 1438. [Google Scholar] [CrossRef] [PubMed]
- Collao, X.; Palacios, G.; Sanbonmatsu-Gámez, S.; Pérez-Ruiz, M.; Negredo, A.I.; Navarro-Marí, J.M.; Grandadam, M.; Aransay, A.M.; Lipkin, W.I.; Tenorio, A.; et al. Genetic diversity of Toscana virus. Emerg. Infect. Dis. 2009, 15, 574–577. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.Y.; Tesh, R.B.; Travassos da Rosa, A.P.A.; Peters, C.J.; Yang, Z.; Guzman, H.; Xiao, S.Y. Phylogenetic relationships among members of the genus Phlebovirus (Bunyaviridae) based on partial M segment sequence analyses. J. Gen. Virol. 2003, 84 (Pt 2), 465–473. [Google Scholar] [CrossRef]
- Venturi, G.; Madeddu, G.; Rezza, G.; Ciccozzi, M.; Pettinato, M.L.; Cilliano, M.; Fiorentini, C.; Mura, M.S.; Ciufolini, M.G. Detection of Toscana virus central nervous system infections in Sardinia Island, Italy. J. Clin. Virol. 2007, 40, 90–91. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, C.; Ayhan, N.; Ortuño, M.; Ortiz, J.; Gould, E.A.; Maia, C.; Berriatua, E.; Charrel, R.N. Experimental infection of dogs with Toscana Virus and Sandfly Fever Sicilian Virus to determine their potential as possible vertebrate Hosts. Microorganisms 2020, 8, 596. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Designation in GenBank | Collection Date | Collection County | S Segment (nr of bp/) | M Segment (nr of bp) | L Segment (nr of bp) | Accession Number | Species ID | Reference |
|---|---|---|---|---|---|---|---|---|
| Alcube virus strain S20 1 | July 2007 | Setúbal | 1758 | 4164 | 6405 | KR363190-192 | Alcube phlebovirus | [110] |
| PoSFPhlebV/21/2007 1 | July 2007 | Setúbal | 1854 | 4221 | 6404 | MW448250-252 | Massilia phlebovirus | [4] |
| PoSFPhlebV/54/2007 1 | August 2007 | Setúbal | 608 | - | 227 | ON807179-180 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/70/2007 2 | September 2007 | Setúbal | 1873 | 4229 | 6386 | MW448253-255 | Massilia phlebovirus | [4] |
| PoSFPhlebV/112/2008 2 | May 2008 | Setúbal | 547 | 461 | 224 | ON807181-183 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/118/2008 2 | June 2008 | Setúbal | 670 | 461 | 227 | ON807184 | Massilia phlebovirus * | This publication |
| Arrabida virus strain PoSFPhlebV/126/2008 1 | June 2008 | Setúbal | 1840 | 4198 | 6391 | KP863799-801 | Massilia phlebovirus | [109] |
| Massilia virus strain 127 1 | June 2008 | Setúbal | 1864 | 4225 | 6404 | KT906098, 100, 102 | Massilia phlebovirus | [110] |
| PoSFPhlebV/128/2008 1 | June 2008 | Setúbal | 310 | - | - | ON807187 | Alcube phlebovirus * | This publication |
| Massilia virus strain 130 3 | June 2008 | Olhão | 1864 | 4225 | 6404 | KT906099, 101, 103 | Massilia phlebovirus | [110] |
| PoSFPhlebV/134/2008 3 | June 2008 | Olhão | 338 | - | - | ON807188 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/137/2008 3 | June 2008 | Olhão | 653 | - | 212 | ON807189-190 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/138/2008 3 | June 2008 | Olhão | 670 | - | - | ON807191 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/149/2008 4 | June 2008 | Olhão | 392 | 461 | 1041 | ON807192-194 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/171/2008 5 | July 2008 | Setúbal | 333 | 461 | - | ON807195-196 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/220/2008 6 | August 2008 | Tavira | 631 | - | - | ON807197 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/237/2008 4 | September 2008 | Olhão | 452 | - | - | ON807198 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/11/2018 7 | May 2018 | Tavira | 381 | - | - | ON807199 | New phlebovirus? | This publication |
| PoSFPhlebV/38/2018 7 | June 2018 | Tavira | 530 | - | - | ON807200 | Massilia phlebovirus * | This publication |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amaro, F.; Zé-Zé, L.; Alves, M.J. Sandfly-Borne Phleboviruses in Portugal: Four and Still Counting. Viruses 2022, 14, 1768. https://doi.org/10.3390/v14081768
Amaro F, Zé-Zé L, Alves MJ. Sandfly-Borne Phleboviruses in Portugal: Four and Still Counting. Viruses. 2022; 14(8):1768. https://doi.org/10.3390/v14081768
Chicago/Turabian StyleAmaro, Fátima, Líbia Zé-Zé, and Maria João Alves. 2022. "Sandfly-Borne Phleboviruses in Portugal: Four and Still Counting" Viruses 14, no. 8: 1768. https://doi.org/10.3390/v14081768
APA StyleAmaro, F., Zé-Zé, L., & Alves, M. J. (2022). Sandfly-Borne Phleboviruses in Portugal: Four and Still Counting. Viruses, 14(8), 1768. https://doi.org/10.3390/v14081768