
Self-Assembly of Porcine Parvovirus Virus-like Particles and Their Application in Serological Assay

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Swine Serum Samples
2.2. Gene Amplification and Optimization
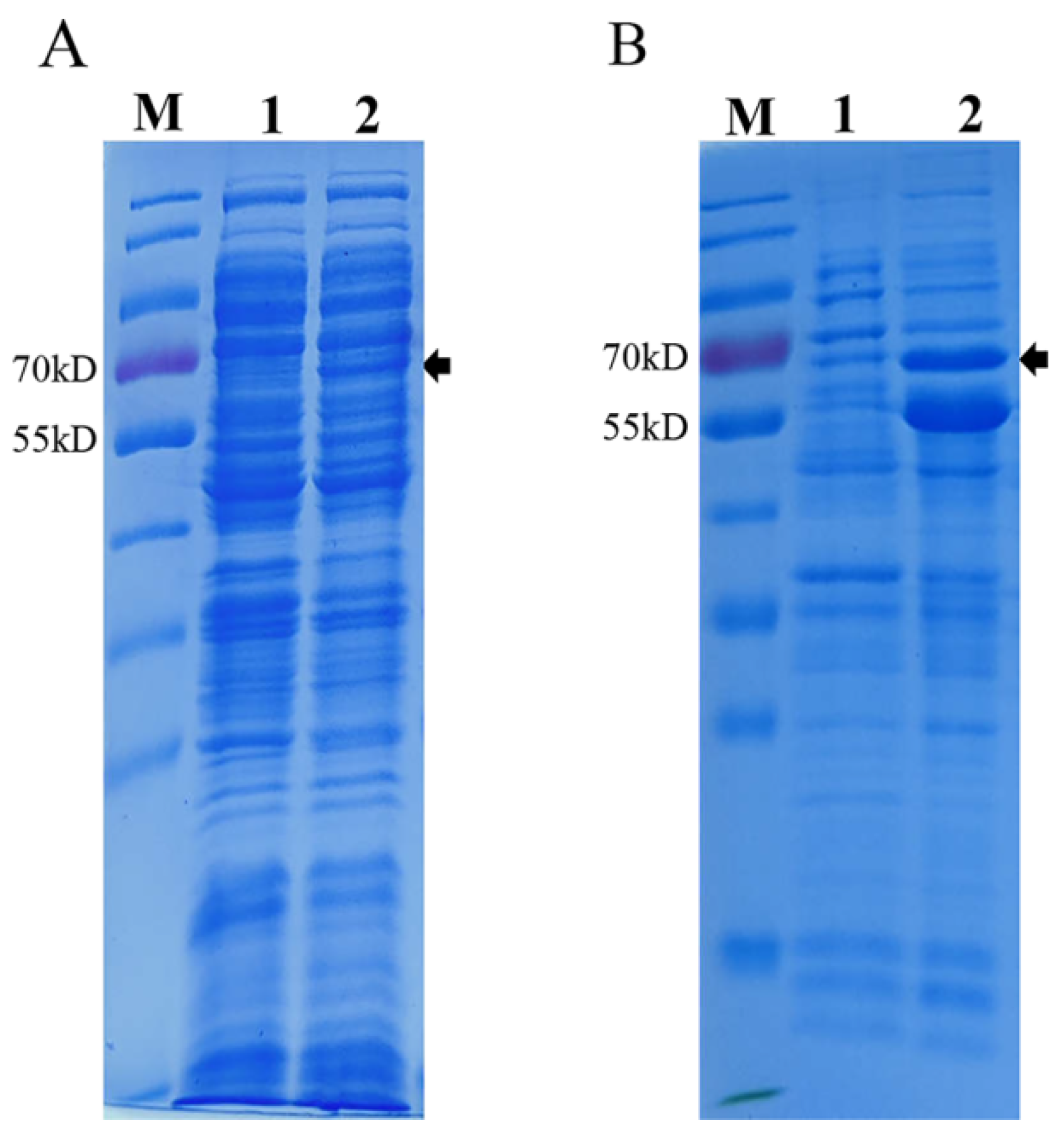
2.3. Construction and Expression of Recombinant VP2 Protein in E. coli
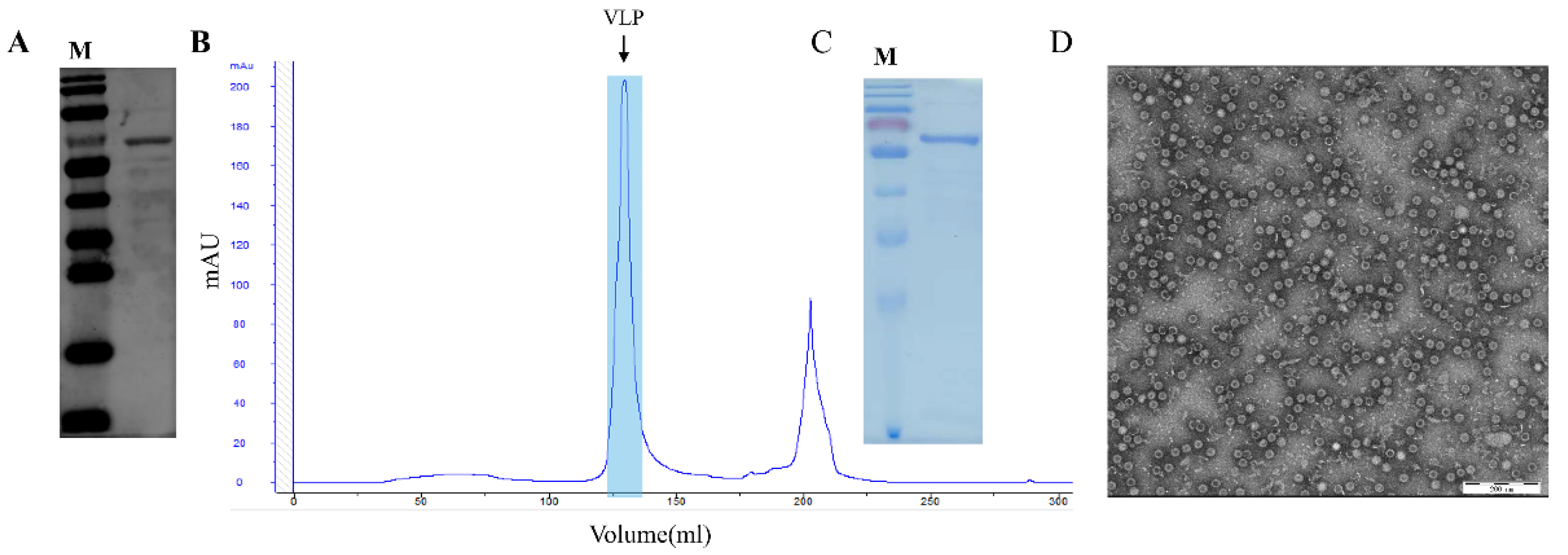
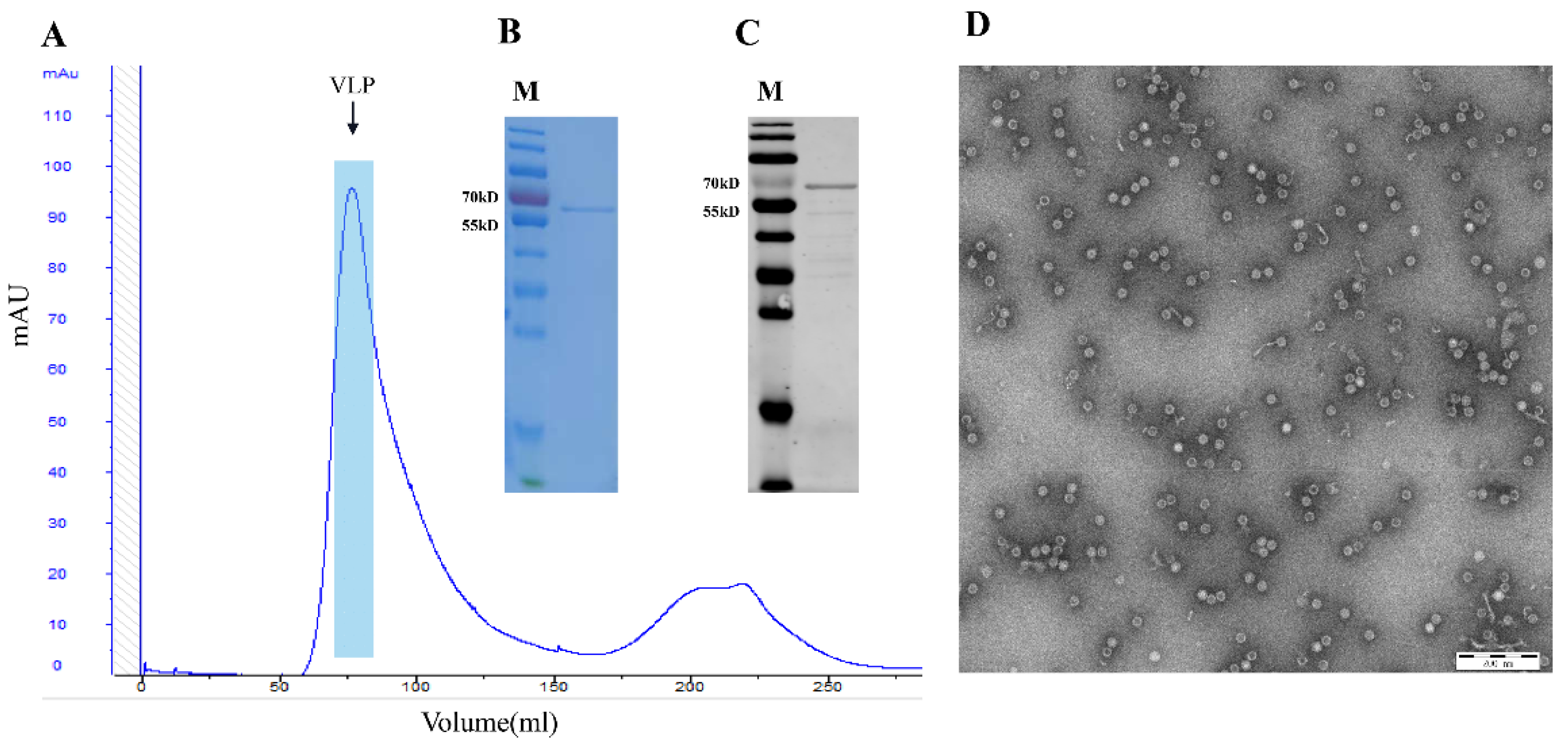
2.4. Purification of PPV-VLPs
2.5. TEM Procedure of PPV-VLPs
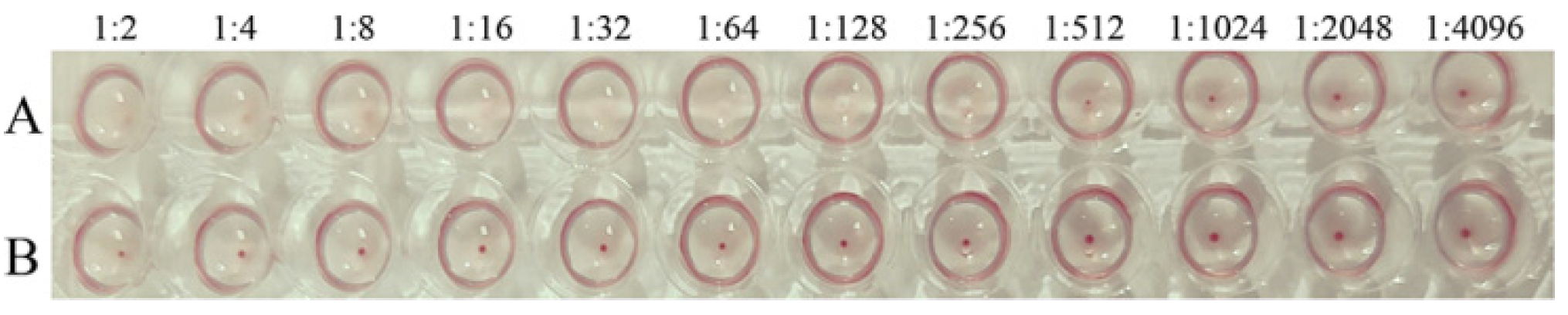
2.6. Determination of Hemagglutination of PPV-VLPs
2.7. Optimization of the PPV-VLP-ELISA Procedure
2.8. Standardization of PPV-VLP-ELISA Procedure
2.9. Determination of Cut-Off Value
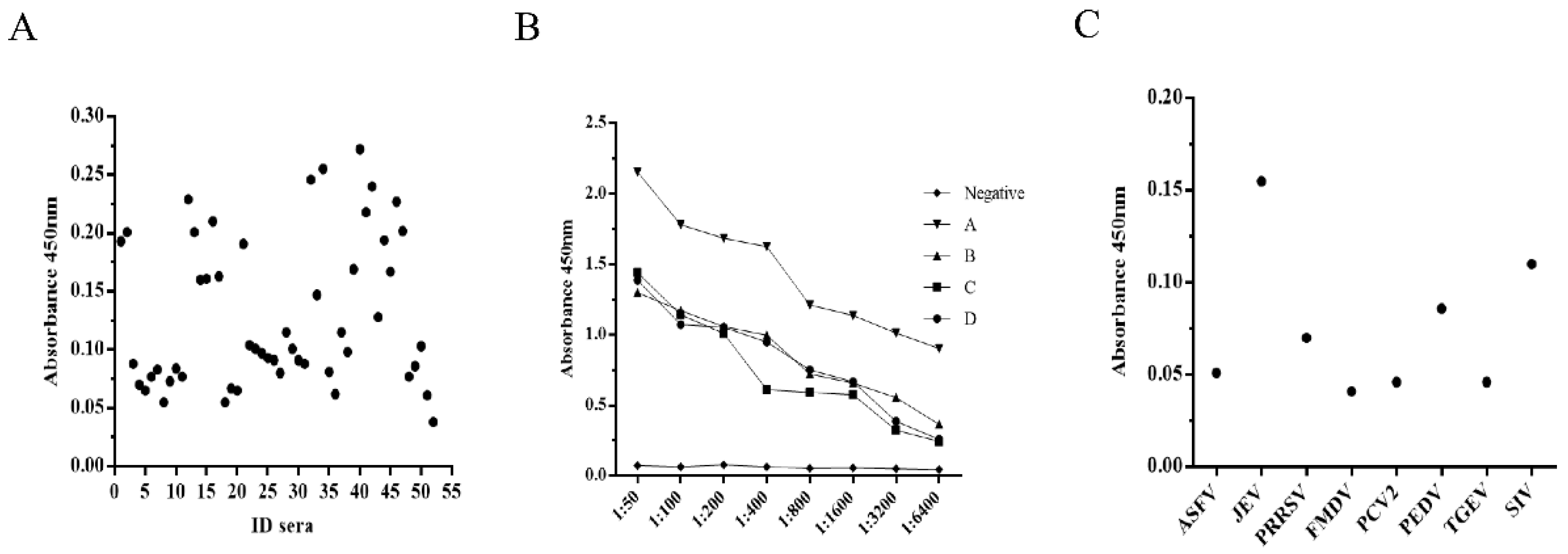
2.10. Reproducibility and Cross-Reactivity Assay
2.11. Cultivation and Proliferation of PPV
2.12. Comparison of the PPV-VLP-ELISA with the Commercial PPV ELISA Kit for Detection of Anti-PPV Antibodies
2.13. Indirect Immunofluorescence Assay Verification of Five Positive Samples Determined by PPV-VLP-ELISA
3. Results
3.1. Expression and Purification of PPV-VLPs from E. coli
3.2. Hemagglutination Activity of PPV-VLPs
3.3. Standardization of the PPV-VLP-ELISA Procedure
3.4. Coincidence Rate with the Ingezim PPV ELISA Kit for Detection of Anti-PPV Antibodies
3.5. Confirmation of PPV-VLP-ELISA Tested Positive Samples with Indirect Immunofluorescence Assay
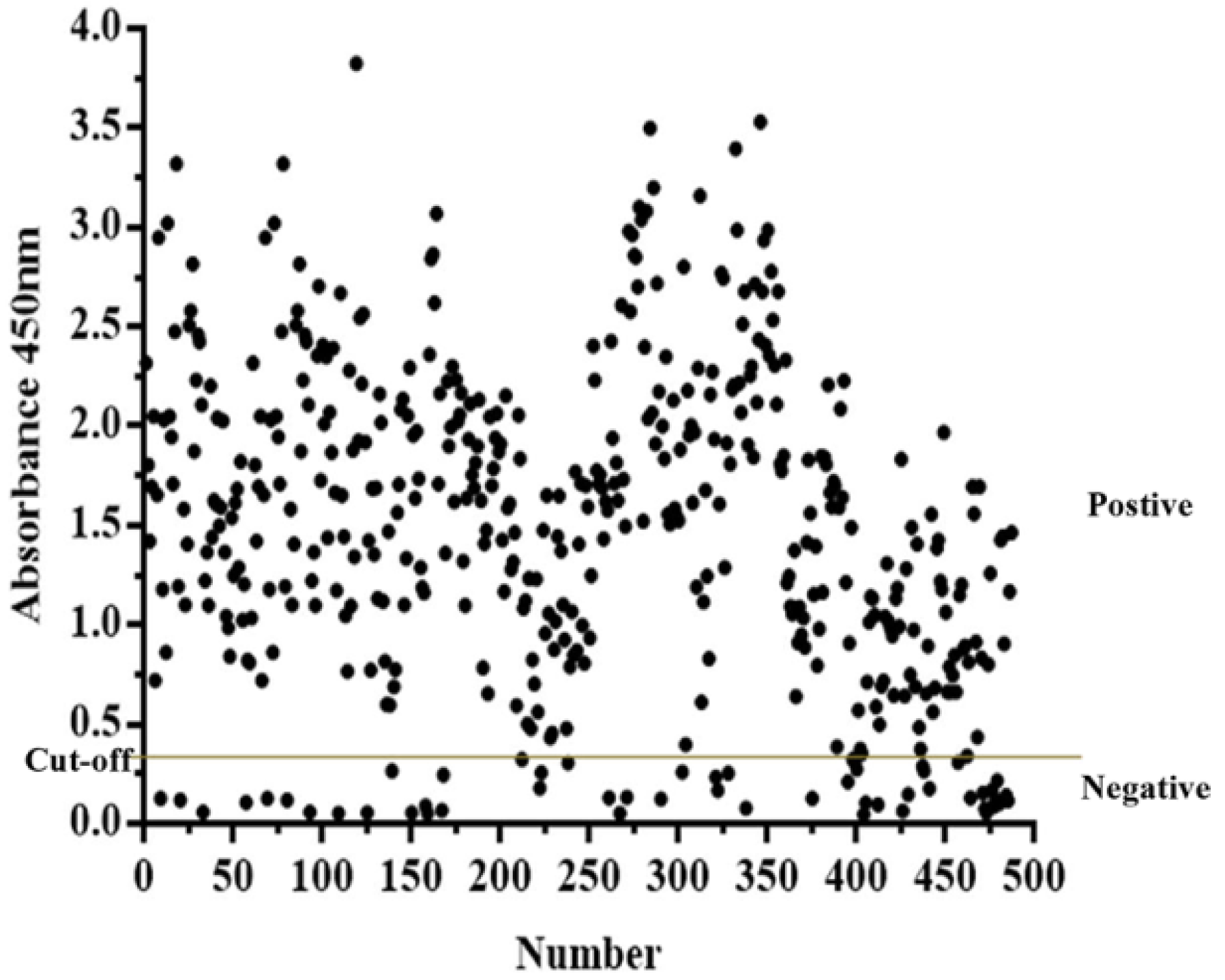
3.6. Application of PPV-VLP-ELISA to Screen Clinical Pig Serum Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Mayr, A.; Bachmann, P.A.; Siegl, G.; Mahnel, H.; Sheffy, B.E. Characterization of a Small Porcine DNA Virus. Arch. Gesamte Virusforsch. 1968, 25, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Cadar, D.; Csagola, A.; Kiss, T.; Tuboly, T. Capsid protein evolution and comparative phylogeny of novel porcine parvoviruses. Mol. Phylogenet. Evol. 2013, 66, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.K.; Wu, G.; Wang, D.; Bayles, D.O.; Lager, K.M.; Vincent, A.L. Identification and molecular cloning of a novel porcine parvovirus. Arch. Virol. 2010, 155, 801–806. [Google Scholar] [CrossRef]
- Hijikata, M.; Abe, K.; Win, K.M.; Shimizu, Y.K.; Keicho, N.; Yoshikura, H. Identification of new parvovirus DNA sequence in swine sera from Myanmar. Jpn. J. Infect. Dis. 2001, 54, 244. [Google Scholar] [PubMed]
- Lau, S.; Woo, P.; Tse, H.; Fu, C.; Au, W.; Chen, X.; Tsoi, H.; Tsang, T.; Chan, J.; Tsang, D.; et al. Identification of novel porcine and bovine parvoviruses closely related to human parvovirus 4. J. Gen. Virol. 2008, 89, 1840–1848. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Qiao, C.; Han, X.; Han, T.; Kang, W.; Zi, Z.; Cao, Z.; Zhai, X.; Cai, X. Identification and genomic characterization of a novel porcine parvovirus (PPV6) in China. Virol. J. 2014, 11, 203. [Google Scholar] [CrossRef]
- Schirtzinger, E.; Suddith, A.; Hause, B.; Hesse, R.A. First identification of porcine parvovirus 6 in North America by viral metagenomic sequencing of serum from pigs infected with porcine reproductive and respiratory syndrome virus. Virol. J. 2015, 12, 170. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Giménez-Lirola, L.; Jiang, Y.; Halbur, P.; Opriessnig, T. Characterization of a novel porcine parvovirus tentatively designated PPV5. PLoS ONE 2013, 8, e65312. [Google Scholar]
- Afolabi, K.O.; Iweriebor, B.C.; Okoh, A.I.; Obi, L.C. Increasing diversity of swine parvoviruses and their epidemiology in African pigs. Infect. Genet. Evol. 2019, 73, 175–183. [Google Scholar] [CrossRef]
- Novosel, D.; Cadar, D.; Tuboly, T.; Jungic, A.; Stadejek, T.; Ait-Ali, T.; Csagola, A. Investigating porcine parvoviruses genogroup 2 infection using in situ polymerase chain reaction. BMC Vet. Res. 2018, 14, 163. [Google Scholar] [CrossRef]
- Ranz, A.I.; Manclús, J.J.; Díaz-Aroca, E.; Casal, J.I. Porcine Parvovirus: DNA Sequence and Genome Organization. J. Gen. Virol. 1989, 70, 2541–2553. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, Y. Induction of immune responses in mice after intragastric administration of Lactobacillus casei producing porcine parvovirus VP2 protein. Appl. Env. Microbiol. 2007, 73, 7041–7047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meszaros, I.; Olasz, F.; Csagola, A.; Tijssen, P.; Zadori, Z. Biology of Porcine Parvovirus (Ungulate parvovirus 1). Viruses 2017, 9, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streck, A.F.; Truyen, U. Porcine Parvovirus. Curr. Issues Mol. Biol. 2020, 37, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Antonis, A.F.; Bruschke, C.J.; Rueda, P.; Maranga, L.; Casal, J.I.; Vela, C.; Hilgers, L.A.; Belt, P.B.; Weerdmeester, K.; Carrondo, M.J.; et al. A novel recombinant virus-like particle vaccine for prevention of porcine parvovirus-induced reproductive failure. Vaccine 2006, 24, 5481–5490. [Google Scholar] [CrossRef]
- Guo, C.; Zhong, Z.; Huang, Y. Production and immunogenicity of VP2 protein of porcine parvovirus expressed in Pichia pastoris. Arch. Virol. 2013, 159, 963–970. [Google Scholar] [CrossRef]
- Yang, D.; Chen, L.; Duan, J.; Yu, Y.; Zhou, J.; Lu, H. Investigation of Kluyveromyces marxianus as a novel host for large-scale production of porcine parvovirus virus-like particles. Microb. Cell Fact. 2021, 20, 24. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Chen, Y.; Wang, A.; Wei, Q.; Liu, D.; Zhang, G. Large-scale manufacture of VP2 VLP vaccine against porcine parvovirus in Escherichia coli with high-density fermentation. Appl. Microbiol. Biotechnol. 2020, 104, 3847–3857. [Google Scholar] [CrossRef]
- Almanza, H.; Cubillos, C.; Angulo, I.; Mateos, F.; Caston, J.R.; van der Poel, W.H.; Vinje, J.; Barcena, J.; Mena, I. Self-assembly of the recombinant capsid protein of a swine norovirus into virus-like particles and evaluation of monoclonal antibodies cross-reactive with a human strain from genogroup II. J. Clin. Microbiol. 2008, 46, 3971–3979. [Google Scholar] [CrossRef] [Green Version]
- Chao, D.Y.; Whitney, M.T.; Davis, B.S.; Medina, F.A.; Munoz, J.L.; Chang, G.J. Comprehensive Evaluation of Differential Serodiagnosis between Zika and Dengue Viral Infections. J. Clin. Microbiol. 2019, 57, e01506-18. [Google Scholar] [CrossRef] [Green Version]
- Nainys, J.; Lasickiene, R.; Petraityte-Burneikiene, R.; Dabrisius, J.; Lelesius, R.; Sereika, V.; Zvirbliene, A.; Sasnauskas, K.; Gedvilaite, A. Generation in yeast of recombinant virus-like particles of porcine circovirus type 2 capsid protein and their use for a serologic assay and development of monoclonal antibodies. BMC Biotechnol. 2014, 14, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, G.; Duan, W.T.; Sun, M.X.; Wang, M.H.; Wang, S.H.; Cai, X.H.; Tu, Y.B. Self-assembly into virus-like particles of the recombinant capsid protein of porcine circovirus type 3 and its application on antibodies detection. AMB Express 2020, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, Z.; Zhan, Y.; Gong, Q.; Yu, W.; Deng, Z.; Wang, A.; Yang, Y.; Wang, N. Generation of E. coli-derived virus-like particles of porcine circovirus type 2 and their use in an indirect IgG enzyme-linked immunosorbent assay. Arch. Virol. 2016, 161, 1485–1491. [Google Scholar] [CrossRef] [PubMed]
- Santi, L.; Huang, Z.; Mason, H. Virus-like particles production in green plants. Methods 2006, 40, 66–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, J.K.; Jeevan-Raj, B.; Netter, H.J. Hepatitis B Virus (HBV) Subviral Particles as Protective Vaccines and Vaccine Platforms. Viruses 2020, 12, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, M.L.; Tiollais, P. Hepatitis B vaccines: Protective efficacy and therapeutic potential. Pathol. Biol. 2010, 58, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Shirbaghaee, Z.; Bolhassani, A. Different applications of virus-like particles in biology and medicine: Vaccination and delivery systems. Biopolymers 2016, 105, 113–132. [Google Scholar] [CrossRef]
- Aires, K.A.; Cianciarullo, A.M.; Carneiro, S.M.; Villa, L.L.; Boccardo, E.; Pérez-Martinez, G.; Perez-Arellano, I.; Oliveira, M.L.S.; Ho, P.L. Production of Human Papillomavirus Type 16 L1 Virus-Like Particles by Recombinant Lactobacillus casei Cells. Appl. Environ. Microbiol. 2006, 72, 745–752. [Google Scholar] [CrossRef] [Green Version]
- Phelps, J.P.; Dao, P.; Jin, H.; Rasochova, L. Expression and self-assembly of cowpea chlorotic mottle virus-like particles in Pseudomonas fluorescens. J. Biotechnol. 2007, 128, 290–296. [Google Scholar] [CrossRef]
- Zhao, Q.; Allen, M.J.; Wang, Y.; Wang, B.; Wang, N.; Shi, L.; Sitrin, R.D. Disassembly and reassembly improves morphology and thermal stability of human papillomavirus type 16 virus-like particles. Nanomedicine 2012, 8, 1182–1189. [Google Scholar] [CrossRef]
- Ding, X.; Liu, D.; Booth, G.; Gao, W.; Lu, Y. Virus-Like Particle Engineering: From Rational Design to Versatile Applications. Biotechnol. J. 2018, 13, e1700324. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Zhai, L.; Tumban, E. Virus-like Particle-Based L2 Vaccines against HPVs: Where Are We Today? Viruses 2019, 12, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamarre, B.; Ryadnov, M.G. Self-assembling viral mimetics: One long journey with short steps. Macromol. Biosci. 2011, 11, 503–513. [Google Scholar] [CrossRef]
- Wang, K.; Zhou, L.; Chen, T.; Li, Q.; Li, J.; Liu, L.; Li, Y.; Sun, J.; Li, T.; Wang, Y.; et al. Engineering for an HPV 9-valent vaccine candidate using genomic constitutive over-expression and low lipopolysaccharide levels in Escherichia coli cells. Microb. Cell Fact. 2021, 20, 227. [Google Scholar] [CrossRef]
- He, J.; Cao, J.; Zhou, N.; Jin, Y.; Wu, J.; Zhou, J. Identification and functional analysis of the novel ORF4 protein encoded by porcine circovirus type 2. J. Virol. 2013, 87, 1420–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.W.; Chang, M.H.; Chang, H.W.; Wu, T.Y.; Chang, Y.C. Parenterally Administered Porcine Epidemic Diarrhea Virus-Like Particle-Based Vaccine Formulated with CCL25/28 Chemokines Induces Systemic and Mucosal Immune Protectivity in Pigs. Viruses 2020, 12, 1122. [Google Scholar] [CrossRef] [PubMed]
- Brune, K.D.; Howarth, M. New Routes and Opportunities for Modular Construction of Particulate Vaccines: Stick, Click, and Glue. Front. Immunol. 2018, 9, 1432. [Google Scholar] [CrossRef]
- Lua, L.H.L.; Connors, N.K.; Sainsbury, F.; Chuan, Y.P.; Wibowo, N.; Middelberg, A.P.J. Bioengineering virus-like particles as vaccines. Biotechnol. Bioeng. 2014, 111, 425–440. [Google Scholar] [CrossRef]
- Nelsen, A.; Lin, C.-M.; Hause, B.M. Porcine Parvovirus 2 Is Predominantly Associated With Macrophages in Porcine Respiratory Disease Complex. Front. Vet. Sci. 2021, 8, 726884. [Google Scholar] [CrossRef]
- Lagan Tregaskis, P.; Staines, A.; Gordon, A.; Sheridan, P.; McMenamy, M.; Duffy, C.; Collins, P.J.; Mooney, M.H.; Lemon, K. Co-infection status of novel parvovirus’s (PPV2 to 4) with porcine circovirus 2 in porcine respiratory disease complex and porcine circovirus-associated disease from 1997 to 2012. Transbound. Emerg. Dis. 2021, 68, 1979–1994. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serum Dilution | Concentration of Coating Antigen (X ± SD, ug/mL) | |||||
|---|---|---|---|---|---|---|
| 10 | 7.5 | 5 | 2.5 | 1 | 0.5 | |
| 1:50 (+) | 1.194 ± 0.0898 | 1.524 ± 0.0757 | 1.675 ± 0.0764 | 1.199 ± 0.0396 | 1.966 ± 0.0233 | 1.225 ± 0.0452 |
| 1:50 (−) | 0.0985 ± 0.002 | 0.0965 ± 0.002 | 0.1565 ± 0.023 | 0.1085 ± 0.013 | 0.1225 ± 0.011 | 0.113 ± 0.01 |
| P/N | 13.13 | 15.78 | 10.70 | 11.05 | 17.47 | 10.84 |
| 1:100 (+) | 0.517 ± 0.021 | 0.8335 ± 0.047 | 1.1095 ± 0.046 | 0.6395 ± 0.122 | 1.52 ± 0.1075 | 0.945 ± 0.006 |
| 1:100 (−) | 0.1 ± 0.002 | 0.1185 ± 0.0021 | 0.149 ± 0.0028 | 0.0915 ± 0.009 | 0.119 ± 0.01 | 0.1105 ± 0.004 |
| P/N | 5.10 | 7.3 | 7.45 | 6.99 | 12.77 | 8.55 |
| 1:150 (+) | 0.728 ± 0.11 | 1.156 ± 0.113 | 1.1011 ± 0.016 | 0.754 ± 0.0339 | 1.268 ± 0.012 | 0.8925 ± 0.0446 |
| 1:150 (−) | 0.127 ± 0.037 | 0.087 ± 0.0212 | 0.0875 ± 0.013 | 0.174 ± 0.0368 | 0.087 ± 0.007 | 0.1185 ± 0.0629 |
| P/N | 5.73 | 13.29 | 11.55 | 5.26 | 12.22 | 6.69 |
| PPV-VLP-ELISA | Commercial Kit | ||
|---|---|---|---|
| Positive | Negative | Total | |
| Positive | 36 | 5 | 41 |
| Negative | 0 | 23 | 23 |
| Total | 36 | 28 | 64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Wang, H.; Wang, S.; Sun, M.; Fang, Z.; Liu, X.; Cai, X.; Tu, Y. Self-Assembly of Porcine Parvovirus Virus-like Particles and Their Application in Serological Assay. Viruses 2022, 14, 1828. https://doi.org/10.3390/v14081828
Gao Y, Wang H, Wang S, Sun M, Fang Z, Liu X, Cai X, Tu Y. Self-Assembly of Porcine Parvovirus Virus-like Particles and Their Application in Serological Assay. Viruses. 2022; 14(8):1828. https://doi.org/10.3390/v14081828
Chicago/Turabian StyleGao, Yanfei, Haiwei Wang, Shanghui Wang, Mingxia Sun, Zheng Fang, Xinran Liu, Xuehui Cai, and Yabin Tu. 2022. "Self-Assembly of Porcine Parvovirus Virus-like Particles and Their Application in Serological Assay" Viruses 14, no. 8: 1828. https://doi.org/10.3390/v14081828
APA StyleGao, Y., Wang, H., Wang, S., Sun, M., Fang, Z., Liu, X., Cai, X., & Tu, Y. (2022). Self-Assembly of Porcine Parvovirus Virus-like Particles and Their Application in Serological Assay. Viruses, 14(8), 1828. https://doi.org/10.3390/v14081828