
Therapeutic Potential of 5′-Methylschweinfurthin G in Merkel Cell Polyomavirus-Positive Merkel Cell Carcinoma

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Lentiviral Transduction
2.2. MTT Assay
2.3. Apoptosis Assay by Flow Cytometry Using Annexin V and 7-AAD Staining
2.4. Migration and Invasion Assays
2.5. Cell Viability Assay
2.6. The 3D Tumor Spheroid Assay
2.7. RNA-Seq Analysis and Protein Detection
3. Results
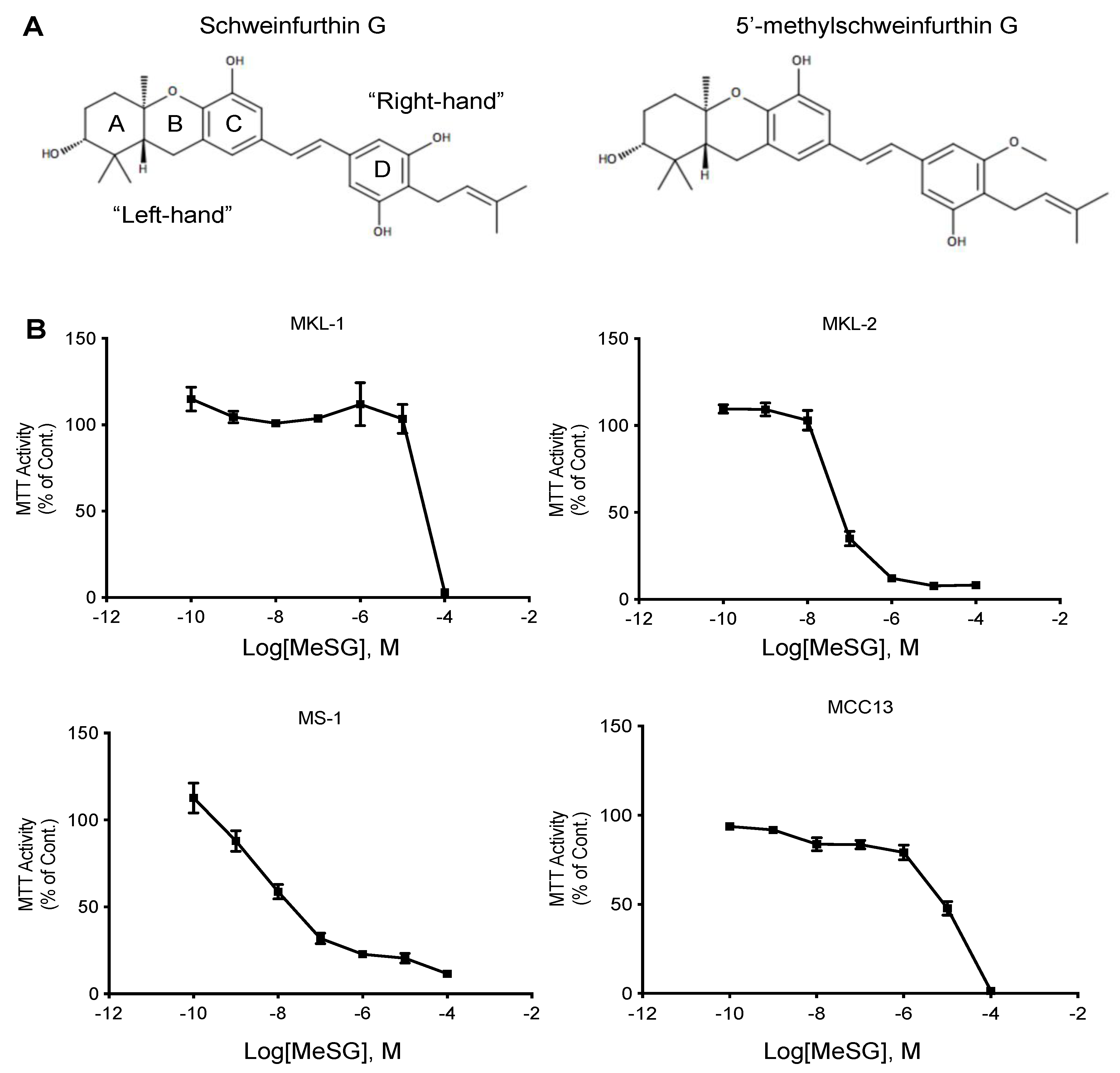
3.1. MeSG Inhibits MCC Cell Metabolic Activity
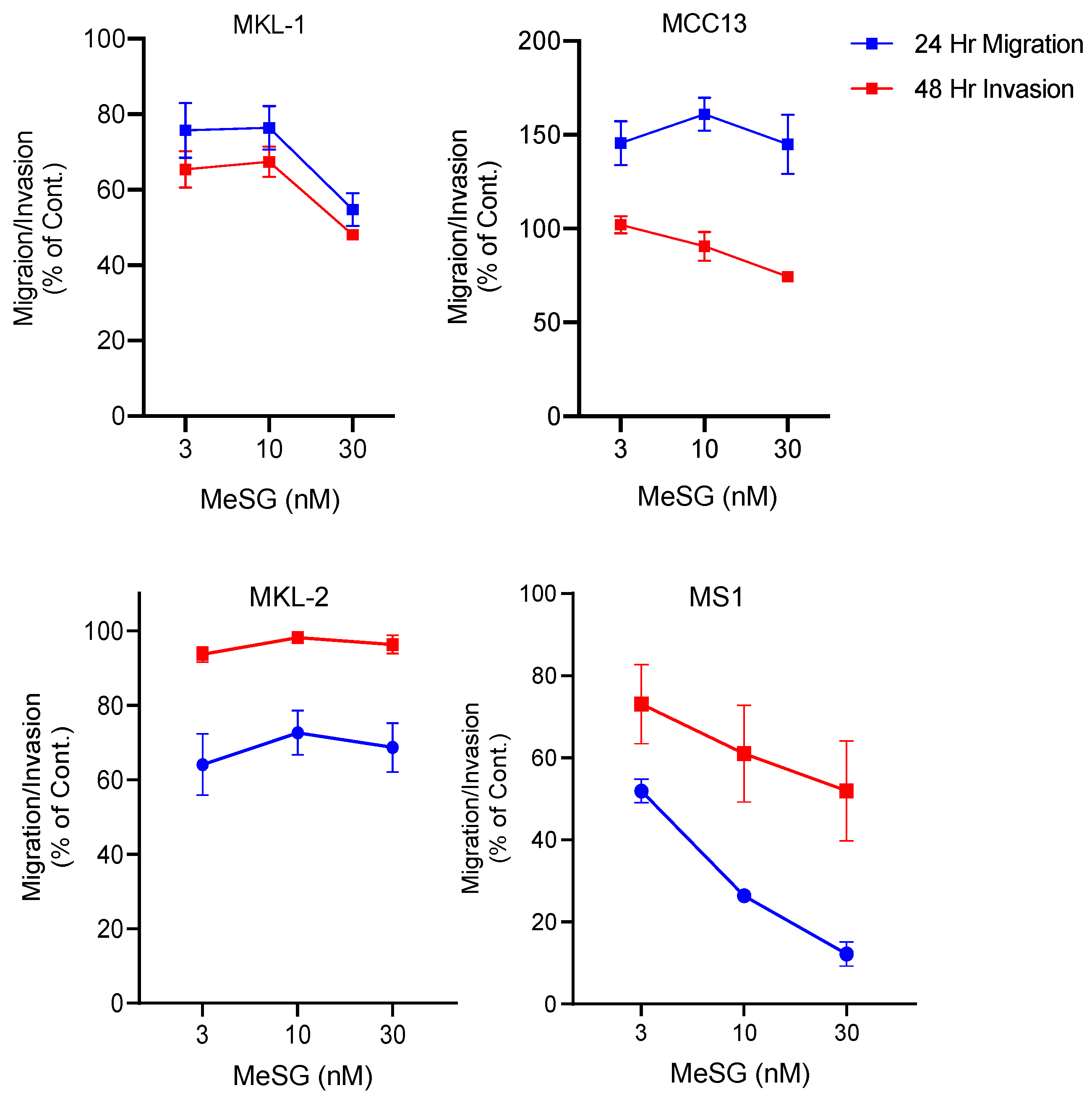
3.2. MeSG Efficiently Impairs Migration and Invasion of Virus-Positive MCC (VP-MCC) Cells
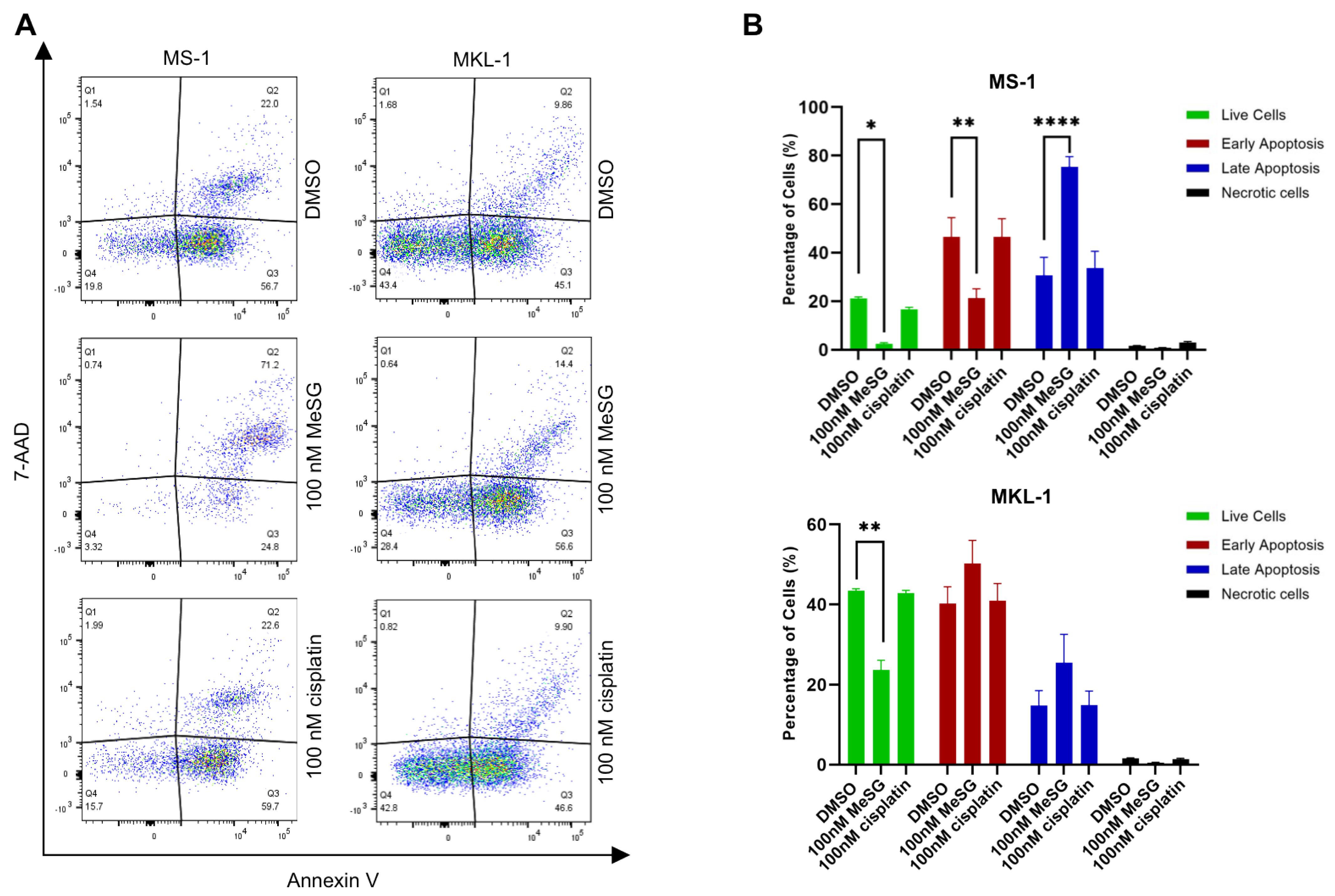
3.3. MeSG Induces Cell Death in MCC
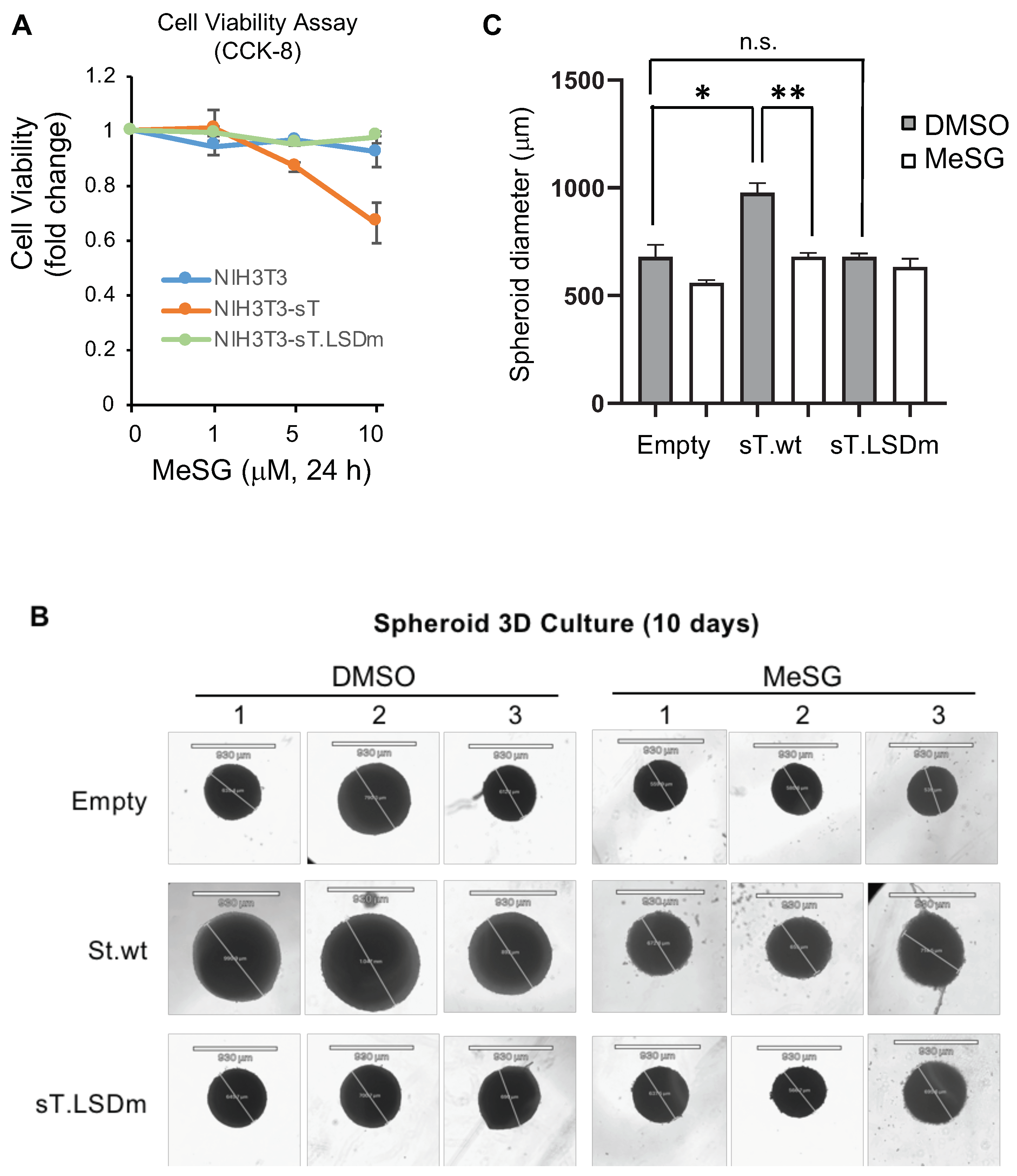
3.4. MeSG Inhibits sT Antigen-Induced Cell Growth Activity
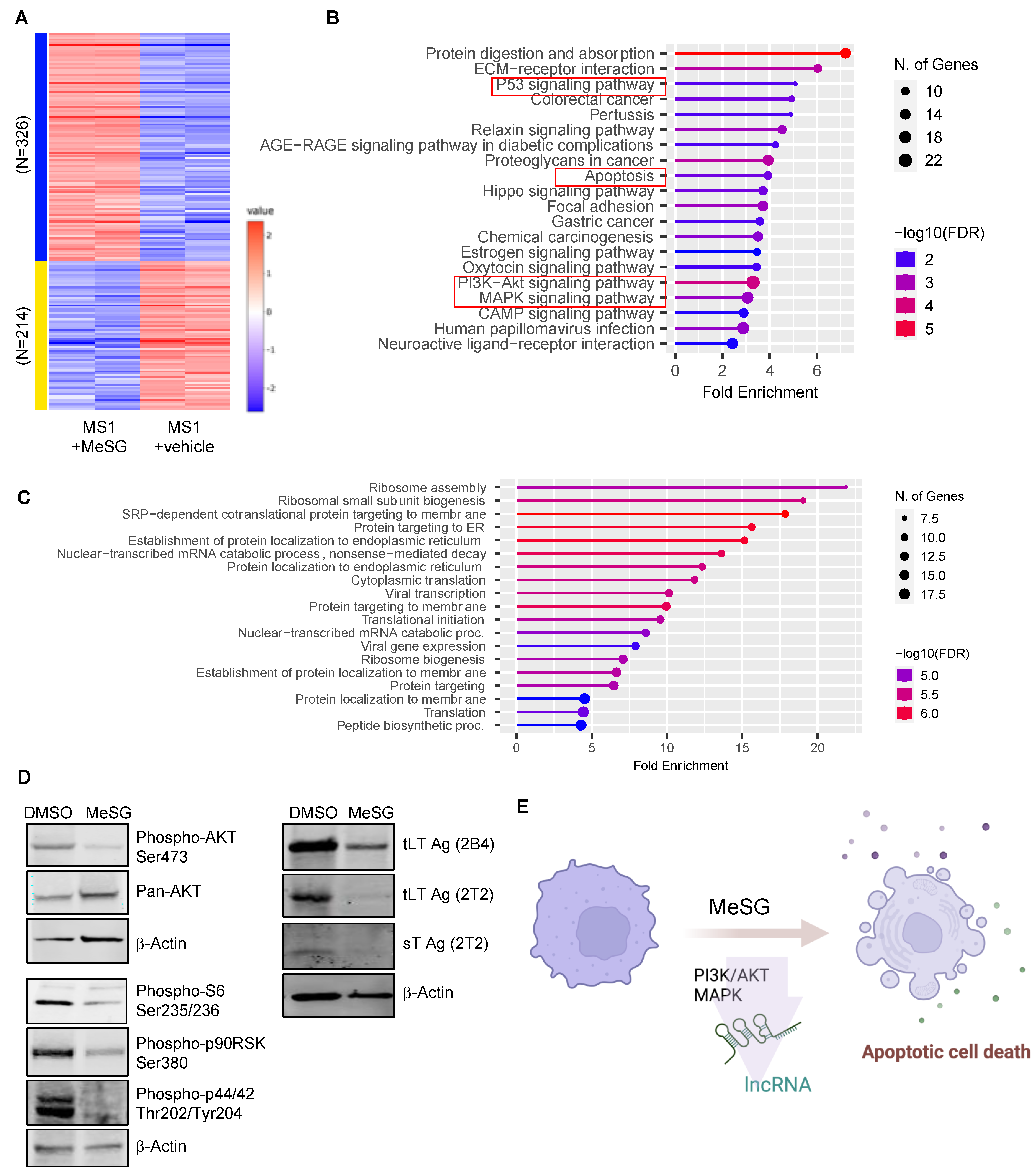
3.5. MeSG Induces Transcriptome Changes in MS1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Demain, A.L.; Vaishnav, P. Natural products for cancer chemotherapy. Microb. Biotechnol. 2011, 4, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, S.; Abbasi, B.H.; Uzair, B.; Abbasi, R. Natural products as reservoirs of novel therapeutic agents. Excli. J. 2018, 17, 420–451. [Google Scholar] [CrossRef] [PubMed]
- Beutler, J.A.; Shoemaker, R.H.; Johnson, T.; Boyd, M.R. Cytotoxic geranyl stilbenes from Macaranga schweinfurthii. J. Nat. Prod. 1998, 61, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- Koubek, E.J.; Weissenrieder, J.S.; Neighbors, J.D.; Hohl, R.J. Schweinfurthins: Lipid Modulators with Promising Anticancer Activity. Lipids 2018, 53, 767–784. [Google Scholar] [CrossRef] [PubMed]
- Harmalkar, D.S.; Mali, J.R.; Sivaraman, A.; Choi, Y.; Lee, K. Schweinfurthins A-Q: Isolation, synthesis, and biochemical properties. RSC Adv. 2018, 8, 21191–21209. [Google Scholar] [CrossRef] [PubMed]
- Treadwell, E.M.; Neighbors, J.D.; Wiemer, D.F. A cascade cyclization approach to schweinfurthin B. Org. Lett. 2002, 4, 3639–3642. [Google Scholar] [CrossRef] [PubMed]
- Neighbors, J.D.; Beutler, J.A.; Wiemer, D.F. Synthesis of nonracemic 3-deoxyschweinfurthin B. J. Org. Chem. 2005, 70, 925–931. [Google Scholar] [CrossRef]
- Mente, N.R.; Neighbors, J.D.; Wiemer, D.F. BF3 x Et2O-mediated cascade cyclizations: Synthesis of schweinfurthins F and G. J. Org. Chem. 2008, 73, 7963–7970. [Google Scholar] [CrossRef]
- Neighbors, J.D.; Salnikova, M.S.; Beutler, J.A.; Wiemer, D.F. Synthesis and structure-activity studies of schweinfurthin B analogs: Evidence for the importance of a D-ring hydrogen bond donor in expression of differential cytotoxicity. Bioorg. Med. Chem. 2006, 14, 1771–1784. [Google Scholar] [CrossRef]
- Kokolus, K.M.; Haley, J.S.; Koubek, E.J.; Gowda, R.; Dinavahi, S.S.; Sharma, A.; Claxton, D.F.; Helm, K.F.; Drabick, J.J.; Robertson, G.P.; et al. Schweinfurthin natural products induce regression of murine melanoma and pair with anti-PD-1 therapy to facilitate durable tumor immunity. Oncoimmunology 2019, 8, e1539614. [Google Scholar] [CrossRef]
- Stevens, J.W.; Meyerholz, D.K.; Neighbors, J.D.; Morcuende, J.A. 5-methylschweinfurthin G reduces chondrosarcoma tumor growth. J. Orthop. Res. 2018, 36, 1283–1293. [Google Scholar] [CrossRef]
- Heath, M.; Jaimes, N.; Lemos, B.; Mostaghimi, A.; Wang, L.C.; Penas, P.E.; Nghiem, P. Clinical characteristics of Merkel cell carcinoma at diagnosis in 195 patients: The AEIOU features. J. Am. Acad. Derm. 2008, 58, 375–381. [Google Scholar] [CrossRef]
- Paulson, K.G.; Park, S.Y.; Vandeven, N.A.; Lachance, K.; Thomas, H.; Chapuis, A.G.; Harms, K.L.; Thompson, J.A.; Bhatia, S.; Stang, A.; et al. Merkel cell carcinoma: Current US incidence and projected increases based on changing demographics. J. Am. Acad. Derm. 2018, 78, 457–463. [Google Scholar] [CrossRef]
- Nwogu, N.; Boyne, J.R.; Dobson, S.J.; Poterlowicz, K.; Blair, G.E.; Macdonald, A.; Mankouri, J.; Whitehouse, A. Cellular sheddases are induced by Merkel cell polyomavirus small tumour antigen to mediate cell dissociation and invasiveness. PLoS Pathog. 2018, 14, e1007276. [Google Scholar] [CrossRef]
- Laikova, K.V.; Oberemok, V.V.; Krasnodubets, A.M.; Gal’chinsky, N.V.; Useinov, R.Z.; Novikov, I.A.; Temirova, Z.Z.; Gorlov, M.V.; Shved, N.A.; Kumeiko, V.V.; et al. Advances in the Understanding of Skin Cancer: Ultraviolet Radiation, Mutations, and Antisense Oligonucleotides as Anticancer Drugs. Molecules 2019, 24, 1516. [Google Scholar] [CrossRef]
- Miller, R.W.; Rabkin, C.S. Merkel cell carcinoma and melanoma: Etiological similarities and differences. Cancer Epidemiol. Biomark. Prev. 1999, 8, 153–158. [Google Scholar]
- Kaae, J.; Hansen, A.V.; Biggar, R.J.; Boyd, H.A.; Moore, P.S.; Wohlfahrt, J.; Melbye, M. Merkel Cell Carcinoma: Incidence, Mortality, and Risk of Other Cancers. J. Natl. Cancer Inst. 2010, 102, 793–801. [Google Scholar] [CrossRef]
- Soltani, A.M.; Allan, B.J.; Best, M.J.; Panthaki, Z.J.; Thaller, S.R. Merkel cell carcinoma of the hand and upper extremity: Current trends and outcomes. J. Plast. Reconstr. Aesthet. Surg. 2014, 67, E71–E77. [Google Scholar] [CrossRef]
- Feng, H.C.; Shuda, M.; Chang, Y.; Moore, P.S. Clonal integration of a polyomavirus in human Merkel cell carcinoma. Science 2008, 319, 1096–1100. [Google Scholar] [CrossRef]
- Wong, S.Q.; Waldeck, K.; Vergara, I.A.; Schröder, J.; Madore, J.; Wilmott, J.S.; Colebatch, A.J.; De Paoli-Iseppi, R.; Li, J.; Lupat, R.; et al. UV-Associated Mutations Underlie the Etiology of MCV-Negative Merkel Cell Carcinomas. Cancer Res. 2015, 75, 5228–5234. [Google Scholar] [CrossRef]
- Harms, P.W.; Collie, A.M.; Hovelson, D.H.; Cani, A.K.; Verhaegen, M.E.; Patel, R.M.; Fullen, D.R.; Omata, K.; Dlugosz, A.A.; Tomlins, S.A.; et al. Next generation sequencing of Cytokeratin 20-negative Merkel cell carcinoma reveals ultraviolet-signature mutations and recurrent TP53 and RB1 inactivation. Mod. Pathol. 2016, 29, 240–248. [Google Scholar] [CrossRef]
- Goh, G.; Walradt, T.; Markarov, V.; Blom, A.; Riaz, N.; Doumani, R.; Stafstrom, K.; Moshiri, A.; Yelistratova, L.; Levinsohn, J.; et al. Mutational landscape of MCPyV-positive and MCPyV-negative Merkel cell carcinomas with implications for immunotherapy. Oncotarget 2016, 7, 3403–3415. [Google Scholar] [CrossRef]
- Chang, Y.; Moore, P.S. Merkel cell carcinoma: A virus-induced human cancer. Annu. Rev. Pathol. 2012, 7, 123–144. [Google Scholar] [CrossRef]
- Verhaegen, M.E.; Mangelberger, D.; Harms, P.W.; Vozheiko, T.D.; Weick, J.W.; Wilbert, D.M.; Saunders, T.L.; Ermilov, A.N.; Bichakjian, C.K.; Johnson, T.M.; et al. Merkel cell polyomavirus small T antigen is oncogenic in transgenic mice. J. Investig. Derm. 2015, 135, 1415–1424. [Google Scholar] [CrossRef]
- Verhaegen, M.E.; Mangelberger, D.; Harms, P.W.; Eberl, M.; Wilbert, D.M.; Meireles, J.; Bichakjian, C.K.; Saunders, T.L.; Wong, S.Y.; Dlugosz, A.A. Merkel Cell Polyomavirus Small T Antigen Initiates Merkel Cell Carcinoma-like Tumor Development in Mice. Cancer Res. 2017, 77, 3151–3157. [Google Scholar] [CrossRef]
- Shuda, M.; Kwun, H.J.; Feng, H.C.; Chang, Y.; Moore, P.S. Human Merkel cell polyomavirus small T antigen is an oncoprotein targeting the 4E-BP1 translation regulator. J. Clin. Investig. 2011, 121, 3623–3634. [Google Scholar] [CrossRef]
- Kwun, H.J.; Shuda, M.; Feng, H.; Camacho, C.J.; Moore, P.S.; Chang, Y. Merkel cell polyomavirus small T antigen controls viral replication and oncoprotein expression by targeting the cellular ubiquitin ligase SCFFbw7. Cell Host Microbe 2013, 14, 125–135. [Google Scholar] [CrossRef]
- Nwogu, N.; Ortiz, L.E.; Kwun, H.J. Surface charge of Merkel cell polyomavirus small T antigen determines cell transformation through allosteric FBW7 WD40 domain targeting. Oncogenesis 2020, 9, 53. [Google Scholar] [CrossRef]
- Kwun, H.J.; Wendzicki, J.A.; Shuda, Y.; Moore, P.S.; Chang, Y. Merkel cell polyomavirus small T antigen induces genome instability by E3 ubiquitin ligase targeting. Oncogene 2017, 36, 6784–6792. [Google Scholar] [CrossRef]
- Nwogu, N.; Ortiz, L.E.; Whitehouse, A.; Kwun, H.J. Merkel Cell Polyomavirus Small Tumor Antigen Activates Matrix Metallopeptidase-9 Gene Expression for Cell Migration and Invasion. J. Virol. 2020, 94, e00786-20. [Google Scholar] [CrossRef]
- Zhao, J.; Jia, Y.; Shen, S.; Kim, J.; Wang, X.; Lee, E.; Brownell, I.; Cho-Vega, J.H.; Lewis, C.; Homsi, J.; et al. Merkel Cell Polyomavirus Small T Antigen Activates Noncanonical NF-κB Signaling to Promote Tumorigenesis. Mol. Cancer Res. 2020, 18, 1623–1637. [Google Scholar] [CrossRef] [PubMed]
- Kwun, H.J.; Shuda, M.; Camacho, C.J.; Gamper, A.M.; Thant, M.; Chang, Y.; Moore, P.S. Restricted protein phosphatase 2A targeting by Merkel cell polyomavirus small T antigen. J. Virol. 2015, 89, 4191–4200. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Rodrik, V.; Pielak, R.M.; Knirr, S.; Zheng, Y.; Foster, D.A. mTOR-dependent suppression of protein phosphatase 2A is critical for phospholipase D survival signals in human breast cancer cells. J. Biol. Chem. 2005, 280, 35829–35835. [Google Scholar] [CrossRef] [PubMed]
- Nghiem, P.; Kaufman, H.L.; Bharmal, M.; Mahnke, L.; Phatak, H.; Becker, J.C. Systematic literature review of efficacy, safety and tolerability outcomes of chemotherapy regimens in patients with metastatic Merkel cell carcinoma. Future Oncol. 2017, 13, 1263–1279. [Google Scholar] [CrossRef] [PubMed]
- Rabinowits, G. Is this the end of cytotoxic chemotherapy in Merkel cell carcinoma? Oncotargets 2017, 10, 4803–4807. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Zheng, W.; Hata Sugi, N.; Agarwala, K.L.; Xu, Q.; Wang, Z.; Tendyke, K.; Lee, W.; Parent, L.; Li, W.; et al. Small molecule schweinfurthins selectively inhibit cancer cell proliferation and mTOR/AKT signaling by interfering with trans-Golgi-network trafficking. Cancer Biol. 2015, 16, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Hafner, C.; Houben, R.; Baeurle, A.; Ritter, C.; Schrama, D.; Landthaler, M.; Becker, J.C. Activation of the PI3K/AKT pathway in Merkel cell carcinoma. PLoS ONE 2012, 7, e31255. [Google Scholar] [CrossRef]
- Nardi, V.; Song, Y.; Santamaria-Barria, J.A.; Cosper, A.K.; Lam, Q.; Faber, A.C.; Boland, G.M.; Yeap, B.Y.; Bergethon, K.; Scialabba, V.L.; et al. Activation of PI3K signaling in Merkel cell carcinoma. Clin. Cancer Res. 2012, 18, 1227–1236. [Google Scholar] [CrossRef]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinform. 2018, 19, 534. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- Weissenrieder, J.S.; Weissenkampen, J.D.; Reed, J.L.; Green, M.V.; Zheng, C.; Neighbors, J.D.; Liu, D.J.; Hohl, R.J. RNAseq reveals extensive metabolic disruptions in the sensitive SF-295 cell line treated with schweinfurthins. Sci. Rep. 2022, 12, 359. [Google Scholar] [CrossRef]
- Mann, J. Natural products in cancer chemotherapy: Past, present and future. Nat. Rev. Cancer 2002, 2, 143–148. [Google Scholar] [CrossRef]
- Kouzmina, M.; Koljonen, V.; Leikola, J.; Böhling, T.; Lantto, E. Frequency and locations of systemic metastases in Merkel cell carcinoma by imaging. Acta Radiol. Open 2017, 6, 2058460117700449. [Google Scholar] [CrossRef]
- Stakaitytė, G.; Nwogu, N.; Dobson, S.J.; Knight, L.M.; Wasson, C.W.; Salguero, F.J.; Blackbourn, D.J.; Blair, G.E.; Mankouri, J.; Macdonald, A.; et al. Merkel Cell Polyomavirus Small T Antigen Drives Cell Motility via Rho-GTPase-Induced Filopodium Formation. J. Virol. 2018, 92, e00940-17. [Google Scholar] [CrossRef]
- Knight, L.M.; Stakaityte, G.; Wood, J.J.; Abdul-Sada, H.; Griffiths, D.A.; Howell, G.J.; Wheat, R.; Blair, G.E.; Steven, N.M.; Macdonald, A.; et al. Merkel cell polyomavirus small T antigen mediates microtubule destabilization to promote cell motility and migration. J. Virol. 2015, 89, 35–47. [Google Scholar] [CrossRef]
- Kuder, C.H.; Sheehy, R.M.; Neighbors, J.D.; Wiemer, D.F.; Hohl, R.J. Functional evaluation of a fluorescent schweinfurthin: Mechanism of cytotoxicity and intracellular quantification. Mol. Pharm. 2012, 82, 9–16. [Google Scholar] [CrossRef]
- Turbyville, T.J.; Gürsel, D.B.; Tuskan, R.G.; Walrath, J.C.; Lipschultz, C.A.; Lockett, S.J.; Wiemer, D.F.; Beutler, J.A.; Reilly, K.M. Schweinfurthin A selectively inhibits proliferation and Rho signaling in glioma and neurofibromatosis type 1 tumor cells in a NF1-GRD-dependent manner. Mol. Cancer 2010, 9, 1234–1243. [Google Scholar] [CrossRef]
- Lockett, S.; Verma, C.; Brafman, A.; Gudla, P.; Nandy, K.; Mimaki, Y.; Fuchs, P.L.; Jaja, J.; Reilly, K.M.; Beutler, J.; et al. Quantitative analysis of F-actin redistribution in astrocytoma cells treated with candidate pharmaceuticals. Cytom. A 2014, 85, 512–521. [Google Scholar] [CrossRef]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharm. 2014, 740, 364–378. [Google Scholar] [CrossRef]
- Nunes, A.S.; Barros, A.S.; Costa, E.C.; Moreira, A.F.; Correia, I.J. 3D tumor spheroids as In Vitro models to mimic In Vivo human solid tumors resistance to therapeutic drugs. Biotechnol. Bioeng. 2019, 116, 206–226. [Google Scholar] [CrossRef]
- Jeffrey, K.L.; Camps, M.; Rommel, C.; Mackay, C.R. Targeting dual-specificity phosphatases: Manipulating MAP kinase signalling and immune responses. Nat. Rev. Drug Discov. 2007, 6, 391–403. [Google Scholar] [CrossRef]
- Gao, N.; Li, Y.; Li, J.; Gao, Z.; Yang, Z.; Liu, H.; Fan, T. Long Non-Coding RNAs: The Regulatory Mechanisms, Research Strategies, and Future Directions in Cancers. Front. Oncol. 2020, 10, 598817. [Google Scholar] [CrossRef]
- Scannevin, R.H.; Alexander, R.; Haarlander, T.M.; Burke, S.L.; Singer, M.; Huo, C.F.; Zhang, Y.M.; Maguire, D.; Spurlino, J.; Deckman, I.; et al. Discovery of a highly selective chemical inhibitor of matrix metalloproteinase-9 (MMP-9) that allosterically inhibits zymogen activation. J. Biol. Chem. 2017, 292, 17963–17974. [Google Scholar] [CrossRef]
- Schrama, D.; Sarosi, E.M.; Adam, C.; Ritter, C.; Kaemmerer, U.; Klopocki, E.; König, E.M.; Utikal, J.; Becker, J.C.; Houben, R. Characterization of six Merkel cell polyomavirus-positive Merkel cell carcinoma cell lines: Integration pattern suggest that large T antigen truncating events occur before or during integration. Int. J. Cancer 2019, 145, 1020–1032. [Google Scholar] [CrossRef]
- Dobson, S.J.; Anene, A.; Boyne, J.R.; Mankouri, J.; Macdonald, A.; Whitehouse, A. Merkel cell polyomavirus small tumour antigen activates the p38 MAPK pathway to enhance cellular motility. Biochem. J. 2020, 477, 2721–2733. [Google Scholar] [CrossRef]
- Fang, B.; Kannan, A.; Zhao, S.; Nguyen, Q.H.; Ejadi, S.; Yamamoto, M.; Camilo Barreto, J.; Zhao, H.; Gao, L. Inhibition of PI3K by copanlisib exerts potent antitumor effects on Merkel cell carcinoma cell lines and mouse xenografts. Sci. Rep. 2020, 10, 8867. [Google Scholar] [CrossRef]
- Lin, Z.; McDermott, A.; Shao, L.; Kannan, A.; Morgan, M.; Stack, B.C.; Moreno, M.; Davis, D.A.; Cornelius, L.A.; Gao, L. Chronic mTOR activation promotes cell survival in Merkel cell carcinoma. Cancer Lett. 2014, 344, 272–281. [Google Scholar] [CrossRef]
- Lin, Z.; Mei, H.; Fan, J.; Yin, Z.; Wu, G. Effect of the dual phosphatidylinositol 3-kinase/mammalian target of rapamycin inhibitor NVP-BEZ235 against human Merkel cell carcinoma MKL-1 cells. Oncol. Lett. 2015, 10, 3663–3667. [Google Scholar] [CrossRef]
- Wang, J.; Lv, X.; Guo, X.; Dong, Y.; Peng, P.; Huang, F.; Wang, P.; Zhang, H.; Zhou, J.; Wang, Y.; et al. Feedback activation of STAT3 limits the response to PI3K/AKT/mTOR inhibitors in PTEN-deficient cancer cells. Oncogenesis 2021, 10, 8. [Google Scholar] [CrossRef]
- Erstad, D.J.; Cusack, J.C. Mutational analysis of merkel cell carcinoma. Cancers 2014, 6, 2116–2136. [Google Scholar] [CrossRef]
- Starrett, G.J.; Thakuria, M.; Chen, T.; Marcelus, C.; Cheng, J.; Nomburg, J.; Thorner, A.R.; Slevin, M.K.; Powers, W.; Burns, R.T.; et al. Clinical and molecular characterization of virus-positive and virus-negative Merkel cell carcinoma. Genome Med. 2020, 12, 30. [Google Scholar] [CrossRef] [PubMed]
- Chteinberg, E.; Rennspiess, D.; Sambo, R.; Tauchmann, S.; Kelleners-Smeets, N.W.J.; Winnepenninckx, V.; Speel, E.J.; Kurz, A.K.; Zenke, M.; Zur Hausen, A. Phosphatidylinositol 3-kinase p110δ expression in Merkel cell carcinoma. Oncotarget 2018, 9, 29565–29573. [Google Scholar] [CrossRef] [PubMed]
- Huang, S. mTOR Signaling in Metabolism and Cancer. Cells 2020, 9, 2278. [Google Scholar] [CrossRef] [PubMed]
- Aboudehen, K. Regulation of mTOR signaling by long non-coding RNA. Biochim. Biophys. Acta Gene Regul. Mech. 2020, 1863, 194449. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.C.; Ni, J.J.; Cui, W.Y.; Wang, B.Y.; Zhuo, W. Emerging roles of lncRNA in cancer and therapeutic opportunities. Am. J. Cancer Res. 2019, 9, 1354–1366. [Google Scholar] [PubMed]
- Wu, B.; Zhang, Y.; Yu, Y.; Zhong, C.; Lang, Q.; Liang, Z.; Lv, C.; Xu, F.; Tian, Y. Long Noncoding RNA H19: A Novel Therapeutic Target Emerging in Oncology. Front. Cell Dev. Biol. 2021, 9, 796740. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, Y.; Chen, K.; Zhang, L. Long Noncoding RNA LINC01134 Promotes Hepatocellular Carcinoma Metastasis via Activating AKT1S1 and NF-κB Signaling. Front. Cell Dev. Biol. 2020, 8, 429. [Google Scholar] [CrossRef]
- Verhaegen, M.E.; Mangelberger, D.; Weick, J.W.; Vozheiko, T.D.; Harms, P.W.; Nash, K.T.; Quintana, E.; Baciu, P.; Johnson, T.M.; Bichakjian, C.K.; et al. Merkel cell carcinoma dependence on bcl-2 family members for survival. J. Investig. Derm. 2014, 134, 2241–2250. [Google Scholar] [CrossRef]
- Kannan, A.; Lin, Z.; Shao, Q.; Zhao, S.; Fang, B.; Moreno, M.A.; Vural, E.; Stack, B.C.; Suen, J.Y.; Kannan, K.; et al. Dual mTOR inhibitor MLN0128 suppresses Merkel cell carcinoma (MCC) xenograft tumor growth. Oncotarget 2016, 7, 6576–6592. [Google Scholar] [CrossRef]
- Chteinberg, E.; Wetzels, S.; Gerritsen, W.; Temmerman, L.; van den Oord, J.; Biessen, E.; Kurz, A.K.; Winnepenninckx, V.; Zenke, M.; Speel, E.J.; et al. Navitoclax combined with Alpelisib effectively inhibits Merkel cell carcinoma cell growth. Adv. Med. Oncol. 2020, 12, 1758835920975621. [Google Scholar] [CrossRef]
- Pham, A.M.; Ortiz, L.E.; Lukacher, A.E.; Kwun, H.J. Cellular senescence preserves viral genome maintenance. BioRxiv 2022. [Google Scholar] [CrossRef]
- Choi, J.E.; Verhaegen, M.E.; Yazdani, S.; Malik, R.; Harms, P.W.; Mangelberger, D.; Tien, J.; Cao, X.; Wang, Y.; Cieślik, M.; et al. Characterizing the Therapeutic Potential of a Potent BET Degrader in Merkel Cell Carcinoma. Neoplasia 2019, 21, 322–330. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koubek, E.J.; Weissenrieder, J.S.; Ortiz, L.E.; Nwogu, N.; Pham, A.M.; Weissenkampen, J.D.; Reed, J.L.; Neighbors, J.D.; Hohl, R.J.; Kwun, H.J. Therapeutic Potential of 5′-Methylschweinfurthin G in Merkel Cell Polyomavirus-Positive Merkel Cell Carcinoma. Viruses 2022, 14, 1848. https://doi.org/10.3390/v14091848
Koubek EJ, Weissenrieder JS, Ortiz LE, Nwogu N, Pham AM, Weissenkampen JD, Reed JL, Neighbors JD, Hohl RJ, Kwun HJ. Therapeutic Potential of 5′-Methylschweinfurthin G in Merkel Cell Polyomavirus-Positive Merkel Cell Carcinoma. Viruses. 2022; 14(9):1848. https://doi.org/10.3390/v14091848
Chicago/Turabian StyleKoubek, Emily J., Jillian S. Weissenrieder, Luz E. Ortiz, Nnenna Nwogu, Alexander M. Pham, J. Dylan Weissenkampen, Jessie L. Reed, Jeffrey D. Neighbors, Raymond J. Hohl, and Hyun Jin Kwun. 2022. "Therapeutic Potential of 5′-Methylschweinfurthin G in Merkel Cell Polyomavirus-Positive Merkel Cell Carcinoma" Viruses 14, no. 9: 1848. https://doi.org/10.3390/v14091848
APA StyleKoubek, E. J., Weissenrieder, J. S., Ortiz, L. E., Nwogu, N., Pham, A. M., Weissenkampen, J. D., Reed, J. L., Neighbors, J. D., Hohl, R. J., & Kwun, H. J. (2022). Therapeutic Potential of 5′-Methylschweinfurthin G in Merkel Cell Polyomavirus-Positive Merkel Cell Carcinoma. Viruses, 14(9), 1848. https://doi.org/10.3390/v14091848