
Inhibition of Cell-Free Translation and Replication of Tobacco Mosaic Virus RNA by Exogenously Added 5′-Proximal Fragments of the Genomic RNA


Abstract
:1. Introduction
2. Materials and Methods
2.1. RNA Preparation
2.2. In Vitro Translation and Replication
2.3. StreptoTag Affinity Purification
2.4. Protein and RNA Analyses
3. Results
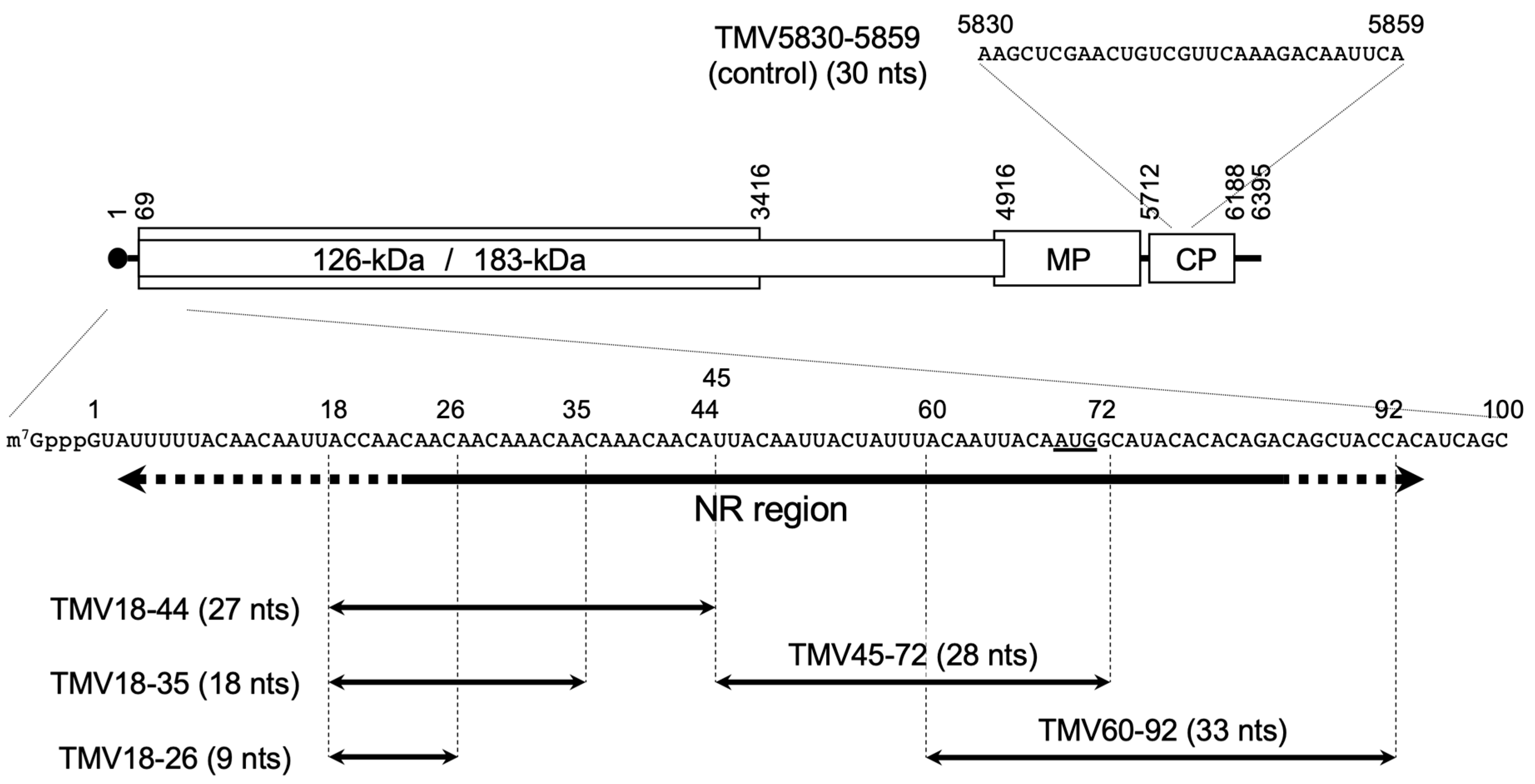
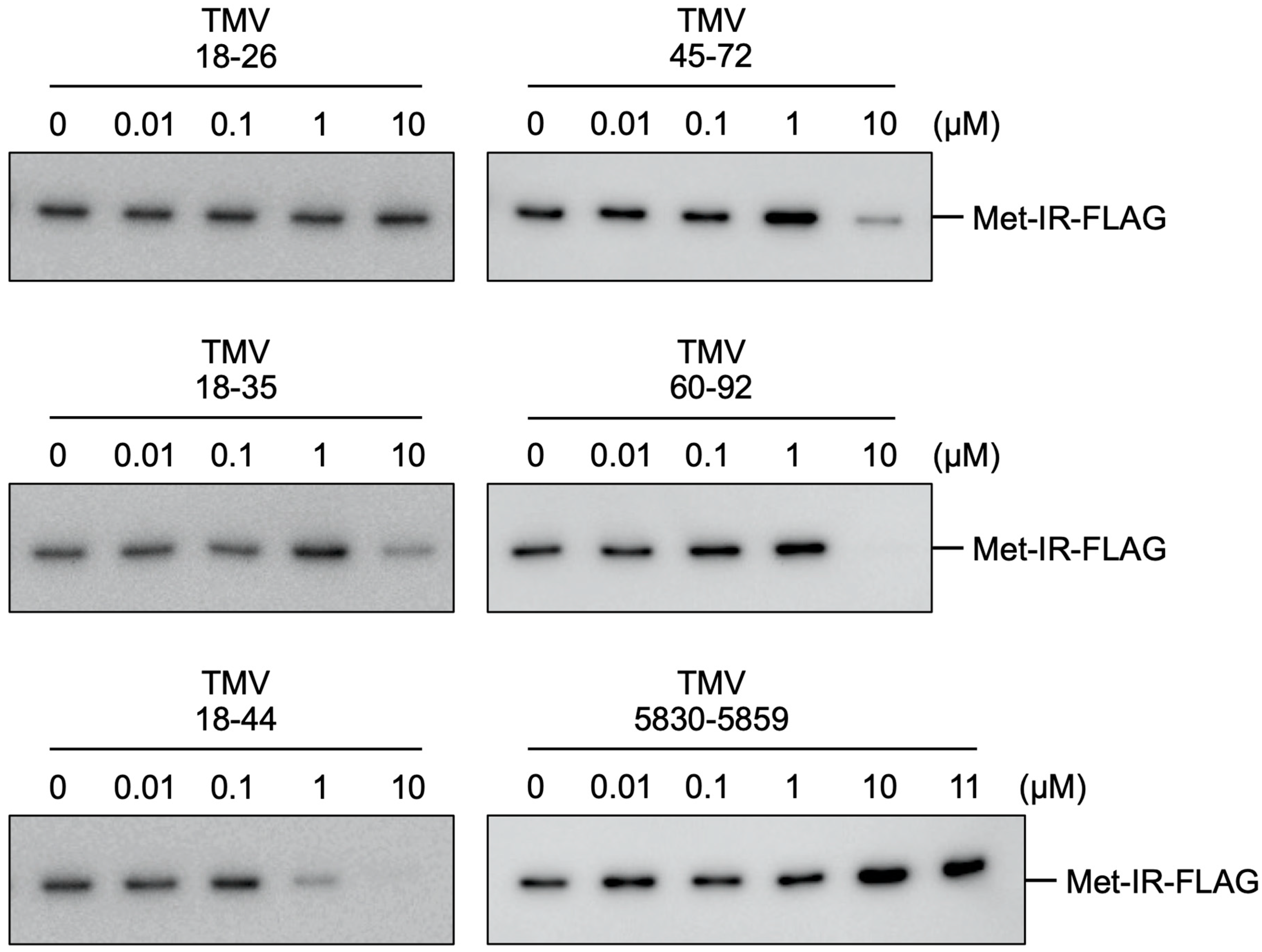
3.1. Inhibition of Binding of Met-IR to the NR Region by RNA Subfragments of This Region

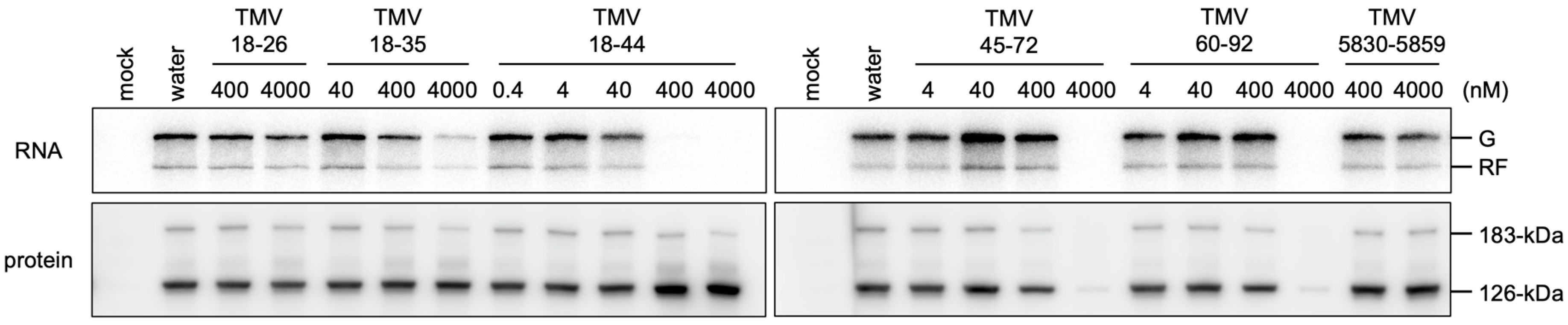
3.2. Inhibition of Cell-Free TMV RNA Replication by RNA Subfragments of the NR Region
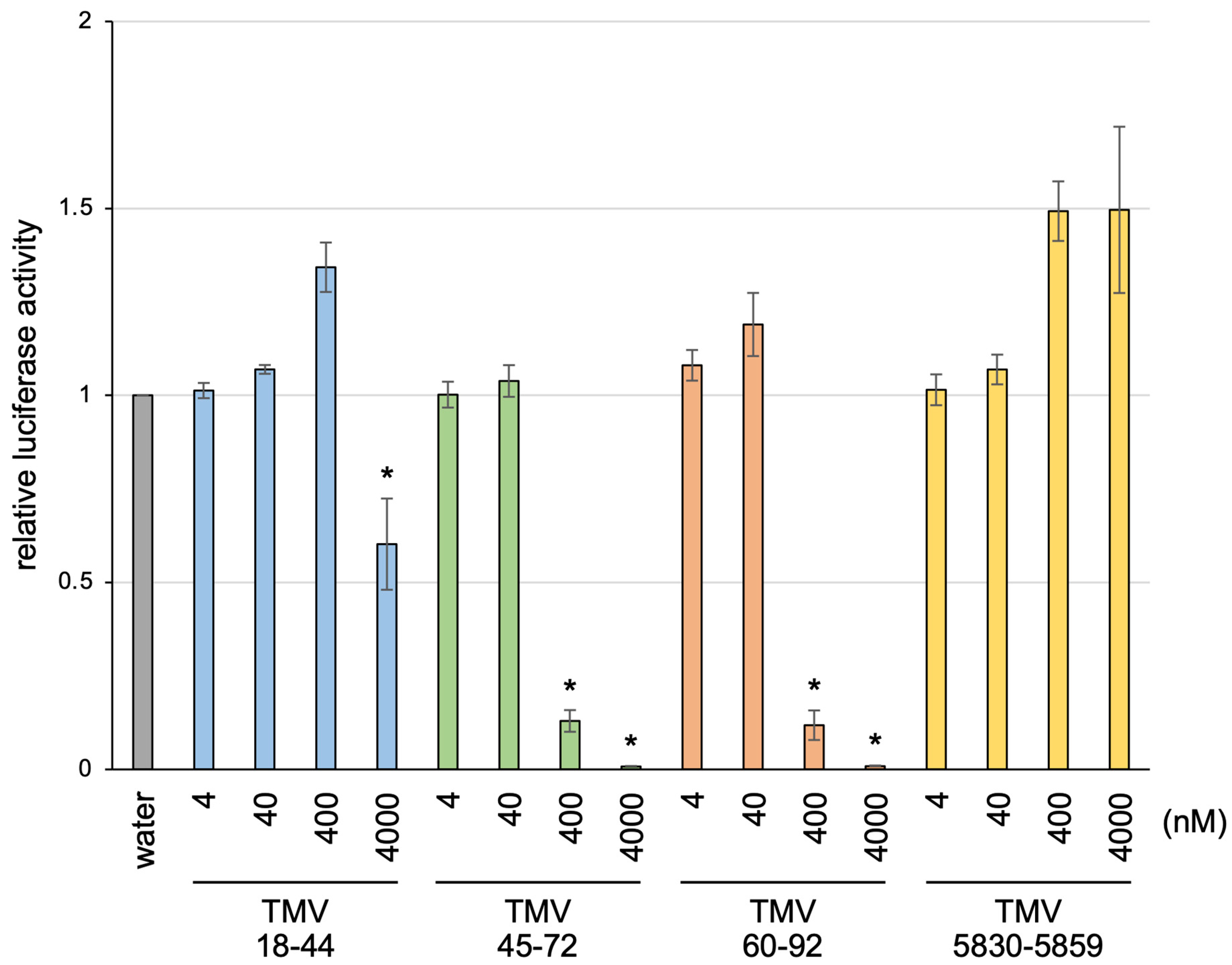
3.3. Inhibition of Translation of Nonviral mRNA by RNA Subfragments of the NR Region
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Adams, M.J.; Adkins, S.; Bragard, C.; Gilmer, D.; Li, D.; MacFarlane, S.A.; Wong, S.M.; Melcher, U.; Ratti, C.; Ryu, K.H.; et al. ICTV virus taxonomy profile: Virgaviridae. J. Gen. Virol. 2017, 98, 1999–2000. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Ishikawa, M. Replication of tobamovirus RNA. Annu. Rev. Phytopathol. 2016, 54, 55–78. [Google Scholar] [CrossRef] [PubMed]
- Chujo, T.; Ishibashi, K.; Miyashita, S.; Ishikawa, M. Functions of the 5′- and 3′-untranslated regions of tobamovirus RNA. Virus Res. 2015, 206, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Gallie, D.R.; Sleat, D.E.; Watts, J.W.; Turner, P.C.; Wilson, T.M.A. The 5′-leader sequence of tobacco mosaic virus RNA enhances the expression of foreign gene transcripts in vitro and in vivo. Nucleic Acids Res. 1987, 15, 3257–3273. [Google Scholar] [CrossRef] [PubMed]
- Sleat, D.E.; Gallie, D.R.; Jefferson, R.A.; Bevan, M.W.; Turner, P.C.; Wilson, T.M.A. Characterisation of the 5′-leader sequence of tobacco mosaic virus RNA as a general enhancer of translation in vitro. Gene 1987, 60, 217–225. [Google Scholar] [CrossRef]
- Komoda, K.; Naito, S.; Ishikawa, M. Replication of plant RNA virus genomes in a cell-free extract of evacuolated plant protoplasts. Proc. Natl. Acad. Sci. USA 2004, 101, 1863–1867. [Google Scholar] [CrossRef] [PubMed]
- Komoda, K.; Mawatari, N.; Hagiwara-Komoda, Y.; Naito, S.; Ishikawa, M. Identification of a ribonucleoprotein intermediate of tomato mosaic virus RNA replication complex formation. J. Virol. 2007, 81, 2584–2591. [Google Scholar] [CrossRef] [PubMed]
- Kawamura-Nagaya, K.; Ishibashi, K.; Huang, Y.P.; Miyashita, S.; Ishikawa, M. Replication protein of tobacco mosaic virus cotranslationally binds the 5′ untranslated region of genomic RNA to enable viral replication. Proc. Natl. Acad. Sci. USA 2014, 111, E1620–E1628. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, D.J.; Dawson, W.O. Functions of the 126- and 183-kDa proteins of tobacco mosaic virus. Virology 2000, 271, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Chiba, Y.; Sakurai, R.; Yoshino, M.; Ominato, K.; Ishikawa, M.; Onouchi, H.; Naito, S. S-adenosyl-L-methionine is an effector in the posttranscriptional autoregulation of the cystathionine γ-synthase gene in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 10225–10230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, K.; Ishikawa, M. A cell-free replication system for positive-strand RNA viruses for identification and characterization of plant resistance gene products. Methods Mol. Biol. 2019, 2028, 115–122. [Google Scholar] [PubMed]
- Bachler, M.; Schroeder, R.; von Ahsen, U. StreptoTag: A novel method for the isolation of RNA-binding proteins. RNA 1999, 5, 1509–1516. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, K.; Ishikawa, M. Identification of an Arabidopsis thaliana protein that binds to tomato mosaic virus genomic RNA and inhibits its multiplication. Virology 2008, 380, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Gallie, D.R.; Walbot, V. Identification of the motifs within the tobacco mosaic virus 5′-leader responsible for enhancing translation. Nucleic Acids Res. 1992, 20, 4631–4638. [Google Scholar] [CrossRef] [PubMed]
- Tanguay, R.L.; Gallie, D.R. Isolation and characterization of the 102-kilodalton RNA-binding protein that binds to the 5′ and 3′ translational enhancers of tobacco mosaic virus RNA. J. Biol. Chem. 1996, 271, 14316–14322. [Google Scholar] [CrossRef] [PubMed]
- Wells, D.R.; Tanguay, R.L.; Le, H.; Gallie, D.R. HSP101 functions as a specific translational regulatory protein whose activity is regulated by nutrient status. Genes Dev. 1998, 12, 3236–3251. [Google Scholar] [CrossRef] [PubMed]
- Gallie, D.R. The 5′-leader of tobacco mosaic virus promotes translation through enhanced recruitment of eIF4F. Nucleic Acids Res. 2002, 30, 3401–3411. [Google Scholar] [CrossRef] [PubMed]
- Agalarov, S.C.; Sakharov, P.A.; Fattakhova, D.K.; Sogorin, E.A.; Spirin, A.S. Internal translation initiation and eIF4F/ATP-independent scanning of mRNA by eukaryotic ribosomal particles. Sci. Rep. 2014, 4, 4438. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) |
|---|---|
| TMV18-26 | A*C*CAACAA*c |
| TMV18-35 | A*C*CAACAACAACAAACA*a |
| TMV18-44 | A*C*CAACAACAACAAACAACAAACAAC*a |
| TMV45-72 | U*U*ACAAUUACUAUUUACAAUUACAAUG*g |
| TMV60-92 | A*C*AAUUACAAUGGCAUACACACAGACAGCUAC*c |
| TMV5830-5859 | A*A*GCUCGAACUGUCGUUCAAAGACAAUUC*a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshida, T.; Ishikawa, M.; Ishibashi, K. Inhibition of Cell-Free Translation and Replication of Tobacco Mosaic Virus RNA by Exogenously Added 5′-Proximal Fragments of the Genomic RNA. Viruses 2022, 14, 1962. https://doi.org/10.3390/v14091962
Yoshida T, Ishikawa M, Ishibashi K. Inhibition of Cell-Free Translation and Replication of Tobacco Mosaic Virus RNA by Exogenously Added 5′-Proximal Fragments of the Genomic RNA. Viruses. 2022; 14(9):1962. https://doi.org/10.3390/v14091962
Chicago/Turabian StyleYoshida, Tetsuya, Masayuki Ishikawa, and Kazuhiro Ishibashi. 2022. "Inhibition of Cell-Free Translation and Replication of Tobacco Mosaic Virus RNA by Exogenously Added 5′-Proximal Fragments of the Genomic RNA" Viruses 14, no. 9: 1962. https://doi.org/10.3390/v14091962
APA StyleYoshida, T., Ishikawa, M., & Ishibashi, K. (2022). Inhibition of Cell-Free Translation and Replication of Tobacco Mosaic Virus RNA by Exogenously Added 5′-Proximal Fragments of the Genomic RNA. Viruses, 14(9), 1962. https://doi.org/10.3390/v14091962