


Production and Evaluation of Chicken Egg Yolk Immunoglobulin (IgY) against Human and Simian Rotaviruses

 , , , and
, , , and 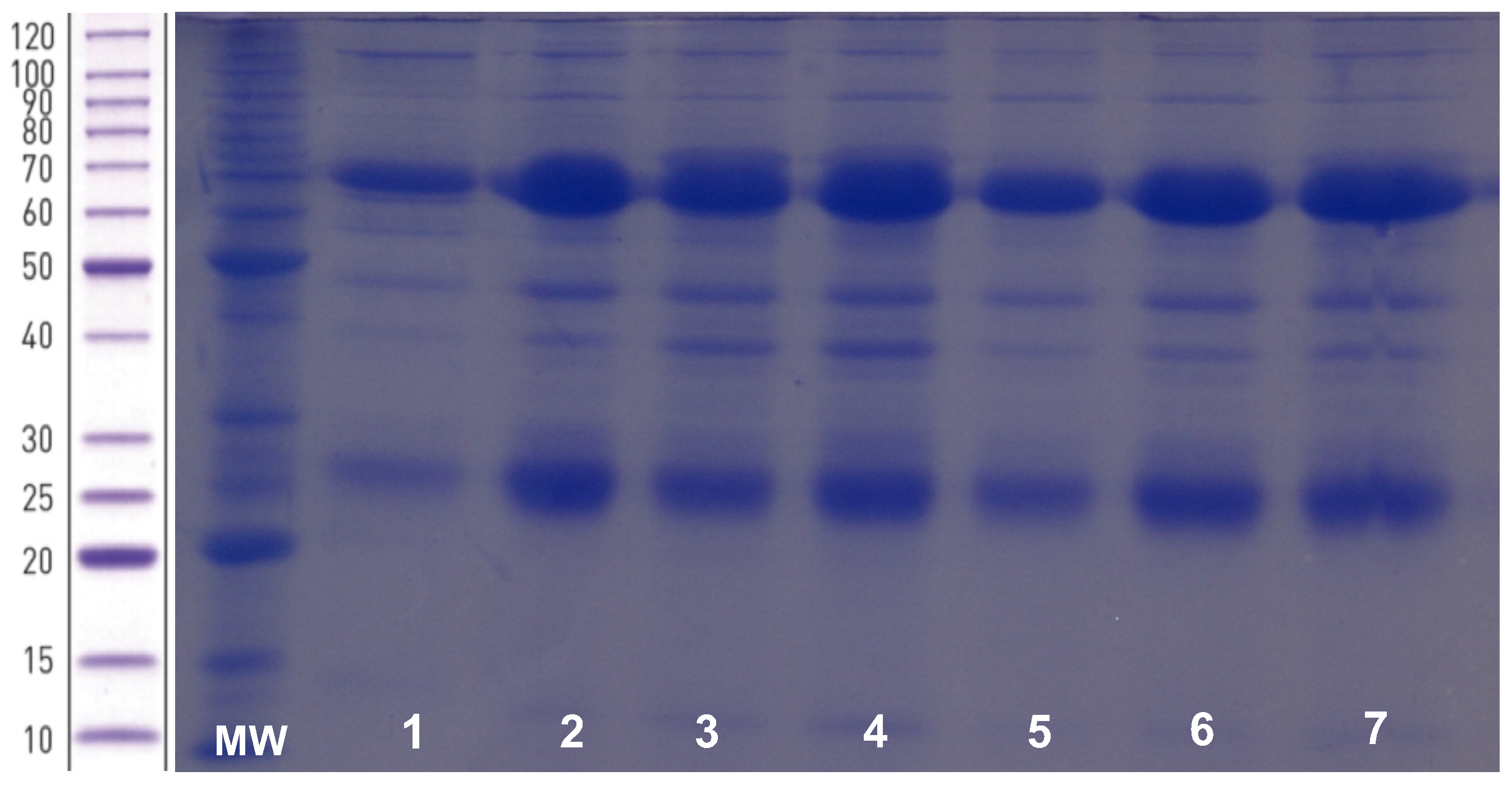{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Inoculum and Immunisation
2.3. Egg Yolk Immunoglobulin (IgY) Purification and Characterisation
2.4. Conjugation of IgY to Carboxyl-Latex Beads
3. Results
3.1. Clinical Monitoring and Immunisation Effects on Hen’s Biology
3.2. Characterisation of the Purified Anti-Rotavirus IgY
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Klemperer, F. Ueber natürliche Immunität und ihre Verwerthung für die Immunisirungstherapie. Arch. Exp. Pathol. Pharmacol. 1893, 31, 356–382. [Google Scholar] [CrossRef]
- Schade, R.; Calzado, E.G.; Sarmiento, R.; Chacana, P.A.; Porankiewicz-Asplund, J.; Terzolo, H.R. Chicken Egg Yolk Antibodies (IgY-Technology): A Review of Progress in Production and Use in Research and Human and Veterinary Medicine. Altern. Lab. Anim. 2005, 33, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Spillner, E.; Braren, I.; Greunke, K.; Seismann, H.; Blank, S.; du Plessis, D. Avian IgY Antibodies and Their Recombinant Equivalents in Research, Diagnostics and Therapy. Biol. J. Int. Assoc. Biol. Stand. 2012, 40, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Parashar, U.D.; World Health Organization–Coordinated Global Rotavirus Surveillance Network. Global, Regional, and National Estimates of Rotavirus Mortality in Children <5 Years of Age, 2000–2013. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2016, 62, S96–S105. [Google Scholar] [CrossRef]
- Estes, M.K.; Greenberg, H.B. Rotaviruses. In Fields Virology; Knipe, D.M., Howley, P.M., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E., Eds.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 1347–1401. [Google Scholar]
- Desselberger, U. Rotaviruses. Virus Res. 2014, 190, 75–96. [Google Scholar] [CrossRef]
- McDonald, S.M.; Patton, J.T. Assortment and Packaging of the Segmented Rotavirus Genome. Trends Microbiol. 2011, 19, 136–144. [Google Scholar] [CrossRef]
- Butz, A.M.; Fosarelli, P.; Dick, J.; Cusack, T.; Yolken, R. Prevalence of Rotavirus on High-Risk Fomites in Day-Care Facilities. Pediatrics 1993, 92, 202–205. [Google Scholar] [CrossRef]
- Kiulia, N.M.; Netshikweta, R.; Page, N.A.; Van Zyl, W.B.; Kiraithe, M.M.; Nyachieo, A.; Mwenda, J.M.; Taylor, M.B. The Detection of Enteric Viruses in Selected Urban and Rural River Water and Sewage in Kenya, with Special Reference to Rotaviruses. J. Appl. Microbiol. 2010, 109, 818–828. [Google Scholar] [CrossRef]
- van Zyl, W.B.; Page, N.A.; Grabow, W.O.K.; Steele, A.D.; Taylor, M.B. Molecular Epidemiology of Group A Rotaviruses in Water Sources and Selected Raw Vegetables in Southern Africa. Appl. Environ. Microbiol. 2006, 72, 4554–4560. [Google Scholar] [CrossRef]
- Bishop, R.F. Natural History of Human Rotavirus Infection. In Viral Gastroenteritis; Chiba, S., Estes, M.K., Nakata, S., Calisher, C.H., Eds.; Springer: Vienna, Austria, 1996; pp. 119–128. [Google Scholar]
- Ray, P.; Fenaux, M.; Sharma, S.; Malik, J.; Subodh, S.; Bhatnagar, S.; Greenberg, H.; Glass, R.I.; Gentsch, J.; Bhan, M.K. Quantitative Evaluation of Rotaviral Antigenemia in Children with Acute Rotaviral Diarrhea. J. Infect. Dis. 2006, 194, 588–593. [Google Scholar] [CrossRef]
- Bernstein, D.I. Rotavirus Overview. Pediatr. Infect. Dis. J. 2009, 28, S50–S53. [Google Scholar] [CrossRef] [PubMed]
- de Paula, V.S.; da Silva, A.D.S.; de Vasconcelos, G.A.L.B.M.; Iff, E.T.; Silva, M.E.M.; Kappel, L.A.; Cruz, P.B.; Pinto, M.A. Applied Biotechnology for Production of Immunoglobulin Y Specific to Hepatitis A Virus. J. Virol. Methods 2011, 171, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Bentes, G.A.; Guimarães, J.R.; Volotão, E.D.M.; Fialho, A.M.; Hooper, C.; Ganime, A.C.; Gardinali, N.R.; Lanzarini, N.M.; da Silva, A.D.S.; Pitcovski, J.; et al. Cynomolgus Monkeys (Macaca Fascicularis) as an Experimental Infection Model for Human Group A Rotavirus. Viruses 2018, 10, 355. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic Transfer of Proteins from Polyacrylamide Gels to Nitrocellulose Sheets: Procedure and Some Applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef]
- Zeng, S.Q.; Halkosalo, A.; Salminen, M.; Szakal, E.D.; Puustinen, L.; Vesikari, T. One-Step Quantitative RT-PCR for the Detection of Rotavirus in Acute Gastroenteritis. J. Virol. Methods 2008, 153, 238–240. [Google Scholar] [CrossRef]
- Bastos-González, D.; Ortega-Vinuesa, J.L.; Lasnieves, D.F.J.; Hidalgo-Alvarez, R. Carboxylated Latexes for Covalent Coupling Antibodies, I. Colloid Interface Sci. 1995, 176, 232–239. [Google Scholar] [CrossRef]
- Ma, H.; O’Kennedy, R. The Structure of Natural and Recombinant Antibodies; Methods in Molecular Biology: Clifton, NJ, USA, 2015; Volume 1348, pp. 7–11. [Google Scholar] [CrossRef]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Methuen Publishing: London, UK, 1959. [Google Scholar]
- Gassmann, M.; Thömmes, P.; Weiser, T.; Hübscher, U. Efficient Production of Chicken Egg Yolk Antibodies against a Conserved Mammalian Protein. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1990, 4, 2528–2532. [Google Scholar] [CrossRef]
- Lévesque, S.; Martinez, G.; Fairbrother, J.M. Improvement of Adjuvant Systems to Obtain a Cost-Effective Production of High Levels of Specific IgY. Poult. Sci. 2007, 86, 630–635. [Google Scholar] [CrossRef]
- Ciriaco, E.; Píñera, P.P.; Díaz-Esnal, B.; Laurà, R. Age-Related Changes in the Avian Primary Lymphoid Organs (Thymus and Bursa of Fabricius). Microsc. Res. Tech. 2003, 62, 482–487. [Google Scholar] [CrossRef]
- Hu, S.L.; Klaniecki, J.; Dykers, T.; Sridhar, P.; Travis, B.M. Neutralizing Antibodies against HIV-1 BRU and SF2 Isolates Generated in Mice Immunized with Recombinant Vaccinia Virus Expressing HIV-1 (BRU) Envelope Glycoproteins and Boosted with Homologous Gp160. AIDS Res. Hum. Retrovir. 1991, 7, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, I.; Sizemore, D.R.; Ahmed, R.K.; Mueller, S.; Wehlin, L.; Scanga, C.; Weichold, F.; Schirru, G.; Pau, M.G.; Goudsmit, J.; et al. RBCG Induces Strong Antigen-Specific T Cell Responses in Rhesus Macaques in a Prime-Boost Setting with an Adenovirus 35 Tuberculosis Vaccine Vector. PLoS ONE 2008, 3, e3790. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Yu, D.H.; Hu, X.D.; Li, S.X.; Zhu, Y.X. A Combined DNA Vaccine-Prime, BCG-Boost Strategy Results in Better Protection against Mycobacterium Bovis Challenge. DNA Cell Biol. 2006, 25, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Lu, S. Heterologous Prime-Boost Vaccination. Curr. Opin. Immunol. 2009, 21, 346–351. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, X.; Song, J.; Wu, J.; Zhu, Y.; Li, M.; Cui, Y.; Chen, Y.; Yang, L.; Liu, J.; et al. Homologous or Heterologous Booster of Inactivated Vaccine Reduces SARS-CoV-2 Omicron Variant Escape from Neutralizing Antibodies. Emerg. Microbes Infect. 2022, 11, 477–481. [Google Scholar] [CrossRef]
- Ayala, I.; Castillo, A.M.; Adánez, G.; Fernández-Rufete, A.; Pérez, B.G.; Castells, M.T. Hyperlipidemic Chicken as a Model of Non-Alcoholic Steatohepatitis. Exp. Biol. Med. 2009, 234, 10–16. [Google Scholar] [CrossRef]
- Loeken, M.R.; Roth, T.F. Analysis of Maternal IgG Subpopulations Which Are Transported into the Chicken Oocyte. Immunology 1983, 49, 21–28. [Google Scholar]
- Schade, R.; Bürger, W.; Schöneberg, T.; Schniering, A.; Schwarzkopf, C.; Hlinak, A.; Kobilke, H. Avian Egg Yolk Antibodies. The Egg Laying Capacity of Hens Following Immunisation with Antigens of Different Kind and Origin and the Efficiency of Egg Yolk Antibodies in Comparison to Mammalian Antibodies. ALTEX 1994, 11, 75–84. [Google Scholar]
- Klimentzou, P.; Paravatou-Petsotas, M.; Zikos, C.; Beck, A.; Skopeliti, M.; Czarnecki, J.; Tsitsilonis, O.; Voelter, W.; Livaniou, E.; Evangelatos, G.P. Development and Immunochemical Evaluation of Antibodies Y for the Poorly Immunogenic Polypeptide Prothymosin Alpha. Peptides 2006, 27, 183–193. [Google Scholar] [CrossRef]
- Nilsson, E.; Hanrieder, J.; Bergquist, J.; Larsson, A. Proteomic Characterization of IgY Preparations Purified with a Water Dilution Method. J. Agric. Food Chem. 2008, 56, 11638–11642. [Google Scholar] [CrossRef]
- Svensson, L.; Sheshberadaran, H.; Vene, S.; Norrby, E.; Grandien, M.; Wadell, G. Serum Antibody Responses to Individual Viral Polypeptides in Human Rotavirus Infections. J. Gen. Virol. 1987, 68, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.L.; McNeal, M.M. VP6: A Candidate Rotavirus Vaccine. J. Infect. Dis. 2010, 202, S101–S107. [Google Scholar] [CrossRef] [PubMed]
- Vleugels, B.; Ververken, C.; Goddeeris, B.M. Stimulatory Effect of CpG Sequences on Humoral Response in Chickens. Poult. Sci. 2002, 81, 1317–1321. [Google Scholar] [CrossRef] [PubMed]
- Vincent, I.E.; Lucifora, J.; Durantel, D.; Hantz, O.; Chemin, I.; Zoulim, F.; Trepo, C. Inhibitory Effect of the Combination of CpG-Induced Cytokines with Lamivudine against Hepatitis B Virus Replication in Vitro. Antivir. Ther. 2009, 14, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.L.; Weeratna, R.; Waldschmidt, T.J.; Tygrett, L.; Schorr, J.; Krieg, A.M.; Weeranta, R. CpG DNA Is a Potent Enhancer of Specific Immunity in Mice Immunized with Recombinant Hepatitis B Surface Antigen. J. Immunol. 1998, 160, 870–876. [Google Scholar]
- Moldoveanu, Z.; Love-Homan, L.; Huang, W.Q.; Krieg, A.M. CpG DNA, a Novel Immune Enhancer for Systemic and Mucosal Immunization with Influenza Virus. Vaccine 1998, 16, 1216–1224. [Google Scholar] [CrossRef]
- Eastcott, J.W.; Holmberg, C.J.; Dewhirst, F.E.; Esch, T.R.; Smith, D.J.; Taubman, M.A. Oligonucleotide Containing CpG Motifs Enhances Immune Response to Mucosally or Systemically Administered Tetanus Toxoid. Vaccine 2001, 19, 1636–1642. [Google Scholar] [CrossRef]
- Mahmood, M.S.; Siddique, M.; Hussain, I.; Khan, A.; Mansoor, M.K. Protection Capability of Recombinant Plasmid DNA Vaccine Containing VP2 Gene of Very Virulent Infectious Bursal Disease Virus in Chickens Adjuvanted with CpG Oligodeoxynucleotide. Vaccine 2006, 24, 4838–4846. [Google Scholar] [CrossRef]
- Wu, F.; Yuan, X.-Y.; Li, J.; Chen, Y.-H. The Co-Administration of CpG-ODN Influenced Protective Activity of Influenza M2e Vaccine. Vaccine 2009, 27, 4320–4324. [Google Scholar] [CrossRef]
- Wang, Y.; Shan, C.; Ming, S.; Liu, Y.; Du, Y.; Jiang, G. Immunoadjuvant Effects of Bacterial Genomic DNA and CpG Oligodeoxynucleotides on Avian Influenza Virus Subtype H5N1 Inactivated Oil Emulsion Vaccine in Chicken. Res. Vet. Sci. 2009, 86, 399–405. [Google Scholar] [CrossRef]
- Mine, Y.; Kovacs-Nolan, J. Chicken Egg Yolk Antibodies as Therapeutics in Enteric Infectious Disease: A Review. J. Med. Food 2002, 5, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Pauly, D.; Dorner, M.; Zhang, X.; Hlinak, A.; Dorner, B.; Schade, R. Monitoring of Laying Capacity, Immunoglobulin Y Concentration, and Antibody Titer Development in Chickens Immunized with Ricin and Botulinum Toxins over a Two-Year Period. Poult. Sci. 2009, 88, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.-C.; Zhang, X.-F.; Tan, M.; Huang, P.; Lei, W.; Fang, H.; Zhong, W.; Jiang, X. A Dual Chicken IgY against Rotavirus and Norovirus. Antivir. Res. 2013, 97, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Vega, C.G.; Bok, M.; Vlasova, A.N.; Chattha, K.S.; Fernández, F.M.; Wigdorovitz, A.; Parreño, V.G.; Saif, L.J. IgY Antibodies Protect against Human Rotavirus Induced Diarrhea in the Neonatal Gnotobiotic Piglet Disease Model. PLoS ONE 2012, 7, e42788. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A.; Sjöquist, J. Novel Latex Agglutination Method with Chicken Anti-Protein A for Detection of Staphylococcus Aureus Infections. J. Clin. Microbiol. 1989, 27, 2856–2857. [Google Scholar] [CrossRef]
- Budama-Kilinc, Y.; Cakir-Koc, R.; Ozdemir, B.; Kaya, Z.; Badur, S. Production and Characterization of a Conserved M2e Peptide-Based Specific IgY Antibody: Evaluation of the Diagnostic Potential via Conjugation with Latex Nanoparticles. Prep. Biochem. Biotechnol. 2018, 48, 930–939. [Google Scholar] [CrossRef]
- Cakir-Koc, R.; Budama-Kilinc, Y.; Ustun, E.; Babur, C. Conjugation and Characterization of Latex Particles with Toxoplasma Gondii-Specific Immunoglobulin Y Antibodies for Diagnostic Aim and Evaluation Efficiency in In Vitro Culture. J. Equine Vet. Sci. 2020, 92, 103145. [Google Scholar] [CrossRef]
- Rieger, A.; Bürger, W.; Hiepe, F.; Schade, R. Determination of Human Serum CRP Using a Chicken Egg Yolk Antibody (IgY) to avoid interferences with rheumatoid factors. Comparison with mammalian antibodies. ALTEX 1996, 13, 57–61. [Google Scholar]
- Larsson, A.; Sjöquist, J. Chicken Antibodies: A Tool to Avoid False Positive Results by Rheumatoid Factor in Latex Fixation Tests. J. Immunol. Methods 1988, 108, 205–208. [Google Scholar] [CrossRef]
- Thu, H.M.; Myat, T.W.; Win, M.M.; Thant, K.Z.; Rahman, S.; Umeda, K.; Nguyen, S.V.; Icatlo, F.C.; Higo-Moriguchi, K.; Taniguchi, K.; et al. Chicken Egg Yolk Antibodies (IgY) for Prophylaxis and Treatment of Rotavirus Diarrhea in Human and Animal Neonates: A Concise Review. Korean J. Food Sci. Anim. Resour. 2017, 37, 1–9. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bentes, G.A.; Lanzarini, N.M.; Guimarães, J.R.; Heinemann, M.B.; Volotão, E.d.M.; da Silva, A.d.S.; Heneine, L.G.D.; de Oliveira, J.M.; Pinto, M.A. Production and Evaluation of Chicken Egg Yolk Immunoglobulin (IgY) against Human and Simian Rotaviruses. Viruses 2022, 14, 1995. https://doi.org/10.3390/v14091995
Bentes GA, Lanzarini NM, Guimarães JR, Heinemann MB, Volotão EdM, da Silva AdS, Heneine LGD, de Oliveira JM, Pinto MA. Production and Evaluation of Chicken Egg Yolk Immunoglobulin (IgY) against Human and Simian Rotaviruses. Viruses. 2022; 14(9):1995. https://doi.org/10.3390/v14091995
Chicago/Turabian StyleBentes, Gentil Arthur, Natália Maria Lanzarini, Juliana Rodrigues Guimarães, Marcos Bryan Heinemann, Eduardo de Mello Volotão, Alexandre dos Santos da Silva, Luiz Guilherme Dias Heneine, Jaqueline Mendes de Oliveira, and Marcelo Alves Pinto. 2022. "Production and Evaluation of Chicken Egg Yolk Immunoglobulin (IgY) against Human and Simian Rotaviruses" Viruses 14, no. 9: 1995. https://doi.org/10.3390/v14091995
APA StyleBentes, G. A., Lanzarini, N. M., Guimarães, J. R., Heinemann, M. B., Volotão, E. d. M., da Silva, A. d. S., Heneine, L. G. D., de Oliveira, J. M., & Pinto, M. A. (2022). Production and Evaluation of Chicken Egg Yolk Immunoglobulin (IgY) against Human and Simian Rotaviruses. Viruses, 14(9), 1995. https://doi.org/10.3390/v14091995