


Construction of a Chikungunya Virus, Replicon, and Helper Plasmids for Transfection of Mammalian Cells
 , ,
, , 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Constructions
2.2. Cell Culture Conditions
2.3. Plasmid Transfection
Expression Kinetics of mKate2 Protein Reporter with Chikungunya Plasmid
2.4. Entry Assay
2.5. CHIKV Infection and Viral Quantification by Plaque Assay
2.6. Ultra-Structural Analysis by Thin-Section Transmission Electron Microscopy
3. Results
3.1. Generation of Viral, Replicon, and Helper CHIKV Plasmids for Expression in Mammalian Cells
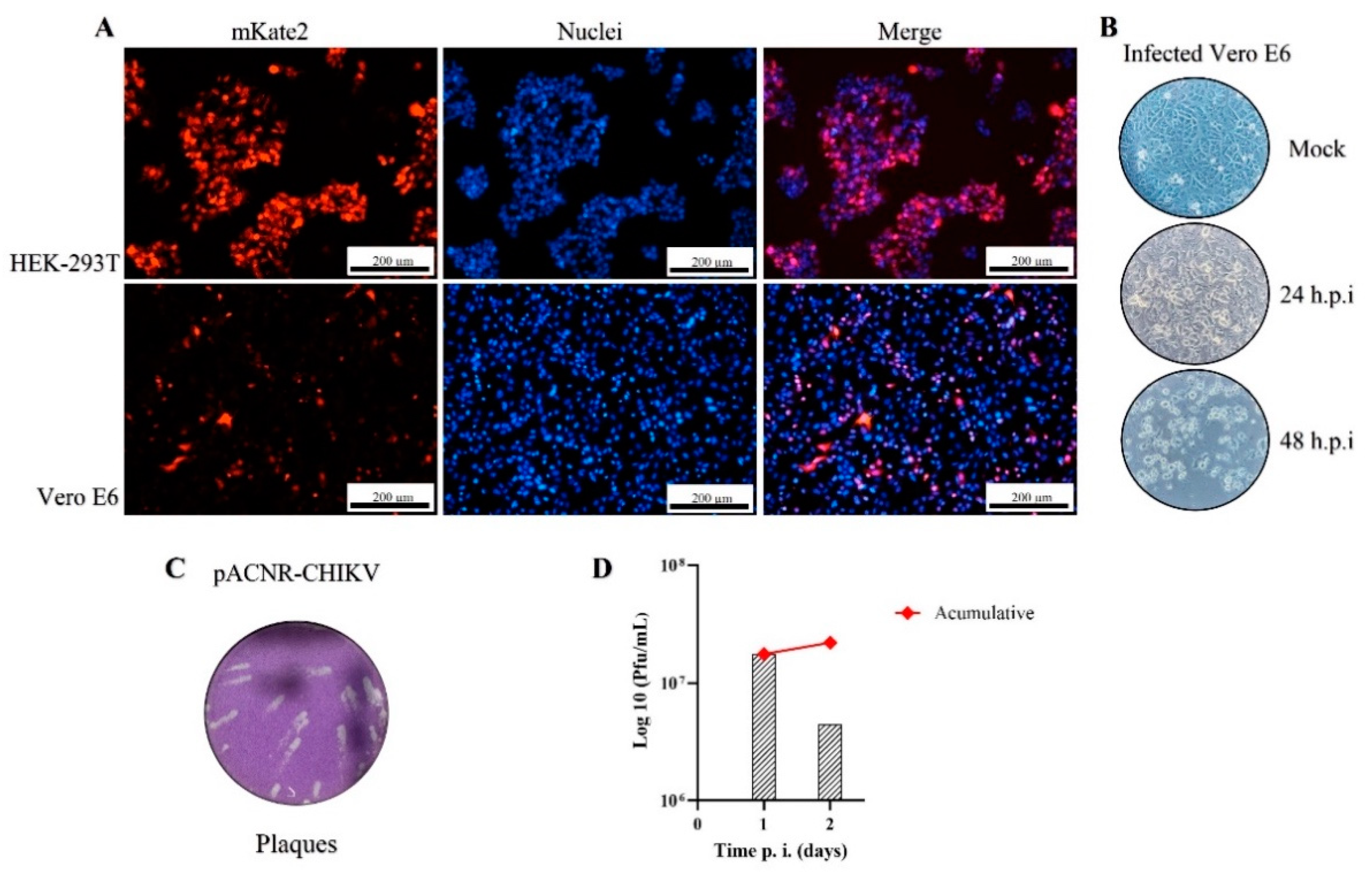
3.2. Viral Production and Analysis of Infection by Microscopy
3.3. The Replicon and Helper Vector System Generates Single-Round Infectious Particles with Gene Delivery Capability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Z.; Wang, J.; Cheng, X.; Hu, H.; Guo, C.; Huang, J.; Chen, Z.; Lu, J. The Worldwide Seroprevalence of DENV, CHIKV and ZIKV Infection: A Systematic Review and Meta-Analysis. PLoS Negl. Trop. Dis. 2021, 15, e0009337. [Google Scholar] [CrossRef] [PubMed]
- Strauss, J.H.; Strauss, E.G. The Alphaviruses: Gene Expression, Replication, and Evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, O.; Albert, M.L. Biology and Pathogenesis of Chikungunya Virus. Nat. Rev. Microbiol. 2010, 8, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Atasheva, S.; McAuley, A.J.; Plante, J.A.; Frolova, E.I.; Beasley, D.W.C.; Frolov, I. Enhancement of Protein Expression by Alphavirus Replicons by Designing Self-Replicating Subgenomic RNAs. Proc. Natl. Acad. Sci. USA 2014, 111, 10708–10713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frolov, I.; Hoffman, T.A.; Prágai, B.M.; Dryga, S.A.; Huang, H.V.; Schlesinger, S.; Rice, C.M. Alphavirus-Based Expression Vectors: Strategies and Applications. Proc. Natl. Acad. Sci. USA 1996, 93, 11371–11377. [Google Scholar] [CrossRef] [Green Version]
- Kamrud, K.I.; Alterson, K.; Custer, M.; Dudek, J.; Goodman, C.; Owens, G.; Smith, J.F. Development and Characterization of Promoterless Helper RNAs for the Production of Alphavirus Replicon Particle. J. Gen. Virol. 2010, 91, 1723–1727. [Google Scholar] [CrossRef]
- Gläsker, S.; Lulla, A.; Lulla, V.; Couderc, T.; Drexler, J.F.; Liljeström, P.; Lecuit, M.; Drosten, C.; Merits, A.; Kümmerer, B.M. Virus Replicon Particle Based Chikungunya Virus Neutralization Assay Using Gaussia Luciferase as Readout. Virol. J. 2013, 10, 235. [Google Scholar] [CrossRef] [Green Version]
- Vrba, S.M.; Kirk, N.M.; Brisse, M.E.; Liang, Y.; Ly, H. Development and Applications of Viral Vectored Vaccines to Combat Zoonotic and Emerging Public Health Threats. Vaccines 2020, 8, 680. [Google Scholar] [CrossRef]
- Frolov, I.; Frolova, E.; Schlesinger, S. Sindbis Virus Replicons and Sindbis Virus: Assembly of Chimeras and of Particles Deficient in Virus RNA. J. Virol. 1997, 71, 2819–2829. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Deng, Y.; Niu, P.; Song, J.; Wang, W.; Du, Y.; Huang, B.; Wang, W.; Zhang, L.; Zhao, P.; et al. Co-Immunization With CHIKV VLP and DNA Vaccines Induces a Promising Humoral Response in Mice. Front. Immunol. 2021, 12, 655743. [Google Scholar] [CrossRef]
- Urakami, A.; Sakurai, A.; Ishikawa, M.; Yap, M.L.; Flores-Garcia, Y.; Haseda, Y.; Aoshi, T.; Zavala, F.P.; Rossmann, M.G.; Kuno, S.; et al. Development of a Novel Virus-Like Particle Vaccine Platform That Mimics the Immature Form of Alphavirus. Clin. Vaccine Immunol. 2017, 24, e00090-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorburger, S.; Hunt, K. Adenoviral Gene Therapy. Oncologist 2002, 7, 46–59. [Google Scholar] [CrossRef]
- Bulcha, J.T.; Wang, Y.; Ma, H.; Tai, P.W.L.; Gao, G. Viral Vector Platforms within the Gene Therapy Landscape. Signal Transduct. Target. Ther. 2021, 6, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Tai, P.W.L.; Gao, G. Adeno-Associated Virus Vector as a Platform for Gene Therapy Delivery. Nat. Rev. Drug Discov. 2019, 18, 358–378. [Google Scholar] [CrossRef]
- Follenzi, A.; Sabatino, G.; Lombardo, A.; Boccaccio, C.; Naldini, L. Efficient Gene Delivery and Targeted Expression to Hepatocytes In Vivo by Improved Lentiviral Vectors. Hum. Gene Ther. 2002, 13, 243–260. [Google Scholar] [CrossRef] [PubMed]
- Phelan, A.; Elliott, G.; O’Hare, P. Intercellular Delivery of Functional P53 by the Herpesvirus Protein VP22. Nat. Biotechnol. 1998, 16, 440–443. [Google Scholar] [CrossRef]
- Gardlík, R.; Pálffy, R.; Hodosy, J.; Lukács, J.; Turna, J.; Celec, P. Vectors and Delivery Systems in Gene Therapy. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2005, 11, RA110-21. [Google Scholar]
- Mukherjee, S.; Thrasher, A.J. Gene Therapy for PIDs: Progress, Pitfalls and Prospects. Gene 2013, 525, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Michel, T.; Link, A.; Abraham, M.-K.; Schlensak, C.; Peter, K.; Wendel, H.-P.; Wang, X.; Krajewski, S. Generation of Cationic Nanoliposomes for the Efficient Delivery of In Vitro Transcribed Messenger RNA. J. Vis. Exp. 2019, 144, e58444. [Google Scholar] [CrossRef]
- Wadhwa, A.; Aljabbari, A.; Lokras, A.; Foged, C.; Thakur, A. Opportunities and Challenges in the Delivery of MRNA-Based Vaccines. Pharmaceutics 2020, 12, 102. [Google Scholar] [CrossRef] [Green Version]
- Liljeström, P.; Garoff, H. A New Generation of Animal Cell Expression Vectors Based on the Semliki Forest Virus Replicon. Bio/Technology 1991, 9, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Wahlfors, J.J.; Zullo, S.A.; Loimas, S.; Nelson, D.M.; Morgan, R.A. Evaluation of Recombinant Alphaviruses as Vectors in Gene Therapy. Gene Ther. 2000, 7, 472–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bredenbeek, P.J.; Frolov, I.; Rice, C.M.; Schlesinger, S. Sindbis Virus Expression Vectors: Packaging of RNA Replicons by Using Defective Helper RNAs. J. Virol. 1993, 67, 6439–6446. [Google Scholar] [CrossRef] [Green Version]
- Vander Veen, R.L.; Harris, D.L.H.; Kamrud, K.I. Alphavirus Replicon Vaccines. Anim. Health Res. Rev. 2012, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Atkins, G.J.; Fleeton, M.N.; Sheahan, B.J. Therapeutic and Prophylactic Applications of Alphavirus Vectors. Expert Rev. Mol. Med. 2008, 10, e33. [Google Scholar] [CrossRef]
- Rayner, J.O.; Dryga, S.A.; Kamrud, K.I. Alphavirus Vectors and Vaccination. Rev. Med. Virol. 2002, 12, 279–296. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, S.-R.; Li, L.-H.; Park, H.-J.; Park, J.-H.; Lee, K.Y.; Kim, M.-K.; Shin, B.A.; Choi, S.-Y. High Cleavage Efficiency of a 2A Peptide Derived from Porcine Teschovirus-1 in Human Cell Lines, Zebrafish and Mice. PLoS ONE 2011, 6, e18556. [Google Scholar] [CrossRef] [Green Version]
- Matusali, G.; Colavita, F.; Bordi, L.; Lalle, E.; Ippolito, G.; Capobianchi, M.; Castilletti, C. Tropism of the Chikungunya Virus. Viruses 2019, 11, 175. [Google Scholar] [CrossRef] [Green Version]
- Dudha, N.; Rana, J.; Rajasekharan, S.; Gabrani, R.; Gupta, A.; Chaudhary, V.K.; Gupta, S. Host-Pathogen Interactome Analysis of Chikungunya Virus Envelope Proteins E1 and E2. Virus Genes 2015, 50, 200–209. [Google Scholar] [CrossRef]
- Wintachai, P.; Thuaud, F.; Basmadjian, C.; Roytrakul, S.; Ubol, S.; Désaubry, L.; Smith, D.R. Assessment of Flavaglines as Potential Chikungunya Virus Entry Inhibitors. Microbiol. Immunol. 2015, 59, 129–141. [Google Scholar] [CrossRef]
- Zhang, R.; Kim, A.S.; Fox, J.M.; Nair, S.; Basore, K.; Klimstra, W.B.; Rimkunas, R.; Fong, R.H.; Lin, H.; Poddar, S.; et al. Mxra8 Is a Receptor for Multiple Arthritogenic Alphaviruses. Nature 2018, 557, 570–574. [Google Scholar] [CrossRef]
- Prado Acosta, M.; Geoghegan, E.M.; Lepenies, B.; Ruzal, S.; Kielian, M.; Martinez, M.G. Surface (S) Layer Proteins of Lactobacillus Acidophilus Block Virus Infection via DC-SIGN Interaction. Front. Microbiol. 2019, 10, 810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Duijl-Richter, M.K.S.; Hoornweg, T.E.; Rodenhuis-Zybert, I.A.; Smit, J.M. Early Events in Chikungunya Virus Infection-From Virus Cell Binding to Membrane Fusion. Viruses 2015, 7, 3647–3674. [Google Scholar] [CrossRef] [PubMed]
- Rangel, M.V.; Stapleford, K.A. Alphavirus Virulence Determinants. Pathogens 2021, 10, 981. [Google Scholar] [CrossRef]
- Bettis, A.A.; L’Azou Jackson, M.; Yoon, I.-K.; Breugelmans, J.G.; Goios, A.; Gubler, D.J.; Powers, A.M. The Global Epidemiology of Chikungunya from 1999 to 2020: A Systematic Literature Review to Inform the Development and Introduction of Vaccines. PLoS Negl. Trop. Dis. 2022, 16, e0010069. [Google Scholar] [CrossRef]
- Powers, A.M. Vaccine and Therapeutic Options To Control Chikungunya Virus. Clin. Microbiol. Rev. 2017, 31, e00104-16. [Google Scholar] [CrossRef] [Green Version]
- Bennett, S.R.; McCarty, J.M.; Ramanathan, R.; Mendy, J.; Richardson, J.S.; Smith, J.; Alexander, J.; Ledgerwood, J.E.; de Lame, P.-A.; Royalty Tredo, S.; et al. Safety and Immunogenicity of PXVX0317, an Aluminium Hydroxide-Adjuvanted Chikungunya Virus-like Particle Vaccine: A Randomised, Double-Blind, Parallel-Group, Phase 2 Trial. Lancet Infect. Dis. 2022, 22, 1343–1355. [Google Scholar] [CrossRef] [PubMed]
- Leung, J.Y.-S.; Ng, M.M.-L.; Chu, J.J.H. Replication of Alphaviruses: A Review on the Entry Process of Alphaviruses into Cells. Adv. Virol. 2011, 2011, 249640. [Google Scholar] [CrossRef] [Green Version]
- Mendes, A.; Kuhn, R.J. Alphavirus Nucleocapsid Packaging and Assembly. Viruses 2018, 10, 138. [Google Scholar] [CrossRef] [Green Version]
- Shin, G.; Yost, S.A.; Miller, M.T.; Elrod, E.J.; Grakoui, A.; Marcotrigiano, J. Structural and Functional Insights into Alphavirus Polyprotein Processing and Pathogenesis. Proc. Natl. Acad. Sci. USA 2012, 109, 16534–16539. [Google Scholar] [CrossRef] [Green Version]
- Hucke, F.I.L.; Bugert, J.J. Current and Promising Antivirals Against Chikungunya Virus. Front. Public Health 2020, 8, 618624. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros-Briones, M.C.; Silva-Pilipich, N.; Herrador-Cañete, G.; Vanrell, L.; Smerdou, C. A New Generation of Vaccines Based on Alphavirus Self-Amplifying RNA. Curr. Opin. Virol. 2020, 44, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Perri, S.; Greer, C.E.; Thudium, K.; Doe, B.; Legg, H.; Liu, H.; Romero, R.E.; Tang, Z.; Bin, Q.; Dubensky, T.W.; et al. An Alphavirus Replicon Particle Chimera Derived from Venezuelan Equine Encephalitis and Sindbis Viruses Is a Potent Gene-Based Vaccine Delivery Vector. J. Virol. 2003, 77, 10394–10403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloom, K.; van den Berg, F.; Arbuthnot, P. Self-Amplifying RNA Vaccines for Infectious Diseases. Gene Ther. 2021, 28, 117–129. [Google Scholar] [CrossRef]
- Erasmus, J.H.; Khandhar, A.P.; O’Connor, M.A.; Walls, A.C.; Hemann, E.A.; Murapa, P.; Archer, J.; Leventhal, S.; Fuller, J.T.; Lewis, T.B.; et al. An Alphavirus-Derived Replicon RNA Vaccine Induces SARS-CoV-2 Neutralizing Antibody and T Cell Responses in Mice and Nonhuman Primates. Sci. Transl. Med. 2020, 12, eabc9396. [Google Scholar] [CrossRef]
- Varnavski, A.N.; Young, P.R.; Khromykh, A.A. Stable High-Level Expression of Heterologous Genes In Vitro and In Vivo by Noncytopathic DNA-Based Kunjin Virus Replicon Vectors. J. Virol. 2000, 74, 4394–4403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ljungberg, K.; Whitmore, A.C.; Fluet, M.E.; Moran, T.P.; Shabman, R.S.; Collier, M.L.; Kraus, A.A.; Thompson, J.M.; Montefiori, D.C.; Beard, C.; et al. Increased Immunogenicity of a DNA-Launched Venezuelan Equine Encephalitis Virus-Based Replicon DNA Vaccine. J. Virol. 2007, 81, 13412–13423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Wall, S.; Ljungberg, K.; Ip, P.P.; Boerma, A.; Knudsen, M.L.; Nijman, H.W.; Liljeström, P.; Daemen, T. Potent Therapeutic Efficacy of an Alphavirus Replicon DNA Vaccine Expressing Human Papilloma Virus E6 and E7 Antigens. OncoImmunology 2018, 7, e1487913. [Google Scholar] [CrossRef] [Green Version]
- Leitner, W.W.; Hwang, L.N.; Deveer, M.J.; Zhou, A.; Silverman, R.H.; Williams, B.R.G.; Dubensky, T.W.; Ying, H.; Restifo, N.P. Alphavirus-Based DNA Vaccine Breaks Immunological Tolerance by Activating Innate Antiviral Pathways. Nat. Med. 2003, 9, 33–39. [Google Scholar] [CrossRef]
- Kümmerer, B.M.; Grywna, K.; Gläsker, S.; Wieseler, J.; Drosten, C. Construction of an Infectious Chikungunya Virus CDNA Clone and Stable Insertion of MCherry Reporter Genes at Two Different Sites. J. Gen. Virol. 2012, 93, 1991–1995. [Google Scholar] [CrossRef]
- Scholte, F.E.M.; Tas, A.; Martina, B.E.E.; Cordioli, P.; Narayanan, K.; Makino, S.; Snijder, E.J.; van Hemert, M.J. Characterization of Synthetic Chikungunya Viruses Based on the Consensus Sequence of Recent E1-226V Isolates. PLoS ONE 2013, 8, e71047. [Google Scholar] [CrossRef] [PubMed]
- Boussier, J.; Levi, L.; Weger-Lucarelli, J.; Poirier, E.Z.; Vignuzzi, M.; Albert, M.L. Chikungunya Virus Superinfection Exclusion Is Mediated by a Block in Viral Replication and Does Not Rely on Non-Structural Protein 2. PLoS ONE 2020, 15, e0241592. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.; Higgs, S.; McGee, C.E.; Lamballerie, X.D.; Charrel, R.N.; Vanlandingham, D.L. Infectious Clones of Chikungunya Virus (La Réunion Isolate) for Vector Competence Studies. Vector-Borne Zoonotic Dis. 2006, 6, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Szurgot, I.; Ljungberg, K.; Kümmerer, B.M.; Liljeström, P. Infectious RNA Vaccine Protects Mice against Chikungunya Virus Infection. Sci. Rep. 2020, 10, 21076. [Google Scholar] [CrossRef] [PubMed]
- Hidajat, R.; Nickols, B.; Forrester, N.; Tretyakova, I.; Weaver, S.; Pushko, P. Next Generation Sequencing of DNA-Launched Chikungunya Vaccine Virus. Virology 2016, 490, 83–90. [Google Scholar] [CrossRef]
- Suzuki, Y.; Tanaka, A.; Maeda, Y.; Emi, A.; Fujioka, Y.; Sakaguchi, S.; Vasudevan, S.G.; Kobayashi, T.; Lim, C.-K.; Takasaki, T.; et al. Construction and Characterization of an Infectious Clone Generated from Chikungunya Virus SL11131 Strain. Virology 2021, 552, 52–62. [Google Scholar] [CrossRef]
- Tretyakova, I.; Hearn, J.; Wang, E.; Weaver, S.; Pushko, P. DNA Vaccine Initiates Replication of Live Attenuated Chikungunya Virus In Vitro and Elicits Protective Immune Response in Mice. J. Infect. Dis. 2014, 209, 1882–1890. [Google Scholar] [CrossRef]
- Maniatis, T.; Fritsch, E.F.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1982; ISBN 978-0-87969-136-3. [Google Scholar]
- Tsetsarkin, K.A.; Kenney, H.; Chen, R.; Liu, G.; Manukyan, H.; Whitehead, S.S.; Laassri, M.; Chumakov, K.; Pletnev, A.G. A Full-Length Infectious CDNA Clone of Zika Virus from the 2015 Epidemic in Brazil as a Genetic Platform for Studies of Virus-Host Interactions and Vaccine Development. mBio 2016, 7, e01114-16. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Hernandez, E.I.; Coronado-Ipiña, M.A.; Colunga-Saucedo, M.; Gonzalez-Sanchez, H.M.; Comas-Garcia, M.; Castillo, C.G. Astrocytes Derived from Neural Progenitor Cells Are 1 Susceptible to Zika Virus Infection. PLoS ONE, 2022; submitted. [Google Scholar]
- Tobin, G.J.; Nagashima, K.; Gonda, M.A. Immunologic and Ultrastructural Characterization of HIV Pseudovirions Containing Gag and Env Precursor Proteins Engineered in Insect Cells. Methods 1996, 10, 208–218. [Google Scholar] [CrossRef]
- Utt, A.; Quirin, T.; Saul, S.; Hellström, K.; Ahola, T.; Merits, A. Versatile Trans-Replication Systems for Chikungunya Virus Allow Functional Analysis and Tagging of Every Replicase Protein. PLoS ONE 2016, 11, e0151616. [Google Scholar] [CrossRef] [Green Version]
- Hyde, J.L.; Chen, R.; Trobaugh, D.W.; Diamond, M.S.; Weaver, S.C.; Klimstra, W.B.; Wilusz, J. The 5′ and 3′ Ends of Alphavirus RNAs—Non-Coding Is Not Non-Functional. Virus Res. 2015, 206, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Steel, J.J.; Henderson, B.R.; Lama, S.B.; Olson, K.E.; Geiss, B.J. Infectious Alphavirus Production from a Simple Plasmid Transfection. Virol. J. 2011, 8, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiss, B.J.; Shimonkevitz, L.H.; Sackal, C.I.; Olson, K.E. Recombination-Ready Sindbis Replicon Expression Vectors for Transgene Expression. Virol. J. 2007, 4, 112. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, M.; Fujikura, D.; Noyori, O.; Kajihara, M.; Maruyama, J.; Miyamoto, H.; Yoshida, R.; Takada, A. A Polymorphism of the TIM-1 IgV Domain: Implications for the Susceptibility to Filovirus Infection. Biochem. Biophys. Res. Commun. 2014, 455, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Martín, C.S.-S.; Liu, C.Y.; Kielian, M. Dealing with Low PH: Entry and Exit of Alphaviruses and Flaviviruses. Trends Microbiol. 2009, 17, 514–521. [Google Scholar] [CrossRef] [Green Version]
- Kaur, P.; Lee, R.C.H.; Chu, J.J.H. Infectious Viral Quantification of Chikungunya Virus-Virus Plaque Assay. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2016; Volume 1426, pp. 93–103. [Google Scholar] [CrossRef]
- Jin, J.; Sherman, M.B.; Chafets, D.; Dinglasan, N.; Lu, K.; Lee, T.-H.; Carlson, L.-A.; Muench, M.O.; Simmons, G. An Attenuated Replication-Competent Chikungunya Virus with a Fluorescently Tagged Envelope. PLoS Negl. Trop. Dis. 2018, 12, e0006693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noranate, N.; Takeda, N.; Chetanachan, P.; Sittisaman, P.; A-nuegoonpipat, A.; Anantapreecha, S. Characterization of Chikungunya Virus-Like Particles. PLoS ONE 2014, 9, e108169. [Google Scholar] [CrossRef]
- Reis, E.V.S.; Damas, B.M.; Mendonça, D.C.; Abrahão, J.S.; Bonjardim, C.A. In-Depth Characterization of the Chikungunya Virus Replication Cycle. J. Virol. 2022, 96, e01732-21. [Google Scholar] [CrossRef]
- Lin, H.-C.; Chiao, D.-J.; Lin, C.-C.; Kuo, S.-C. Facile Method for Delivering Chikungunya Viral Replicons into Mosquitoes and Mammalian Cells. Sci. Rep. 2021, 11, 12321. [Google Scholar] [CrossRef]
- Pohjala, L.; Utt, A.; Varjak, M.; Lulla, A.; Merits, A.; Ahola, T.; Tammela, P. Inhibitors of Alphavirus Entry and Replication Identified with a Stable Chikungunya Replicon Cell Line and Virus-Based Assays. PLoS ONE 2011, 6, e28923. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colunga-Saucedo, M.; Rubio-Hernandez, E.I.; Coronado-Ipiña, M.A.; Rosales-Mendoza, S.; Castillo, C.G.; Comas-Garcia, M. Construction of a Chikungunya Virus, Replicon, and Helper Plasmids for Transfection of Mammalian Cells. Viruses 2023, 15, 132. https://doi.org/10.3390/v15010132
Colunga-Saucedo M, Rubio-Hernandez EI, Coronado-Ipiña MA, Rosales-Mendoza S, Castillo CG, Comas-Garcia M. Construction of a Chikungunya Virus, Replicon, and Helper Plasmids for Transfection of Mammalian Cells. Viruses. 2023; 15(1):132. https://doi.org/10.3390/v15010132
Chicago/Turabian StyleColunga-Saucedo, Mayra, Edson I. Rubio-Hernandez, Miguel A. Coronado-Ipiña, Sergio Rosales-Mendoza, Claudia G. Castillo, and Mauricio Comas-Garcia. 2023. "Construction of a Chikungunya Virus, Replicon, and Helper Plasmids for Transfection of Mammalian Cells" Viruses 15, no. 1: 132. https://doi.org/10.3390/v15010132
APA StyleColunga-Saucedo, M., Rubio-Hernandez, E. I., Coronado-Ipiña, M. A., Rosales-Mendoza, S., Castillo, C. G., & Comas-Garcia, M. (2023). Construction of a Chikungunya Virus, Replicon, and Helper Plasmids for Transfection of Mammalian Cells. Viruses, 15(1), 132. https://doi.org/10.3390/v15010132