
Biological Properties of 12 Newly Isolated Acinetobacter baumannii-Specific Bacteriophages
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Search, Isolation, and Amplification of Bacteriophages
2.3. Lytic Spectrum of Examined Phages
2.4. Multiplicity of Infection (MOI)
2.5. Phage Adsorption to Host Bacterial Cells
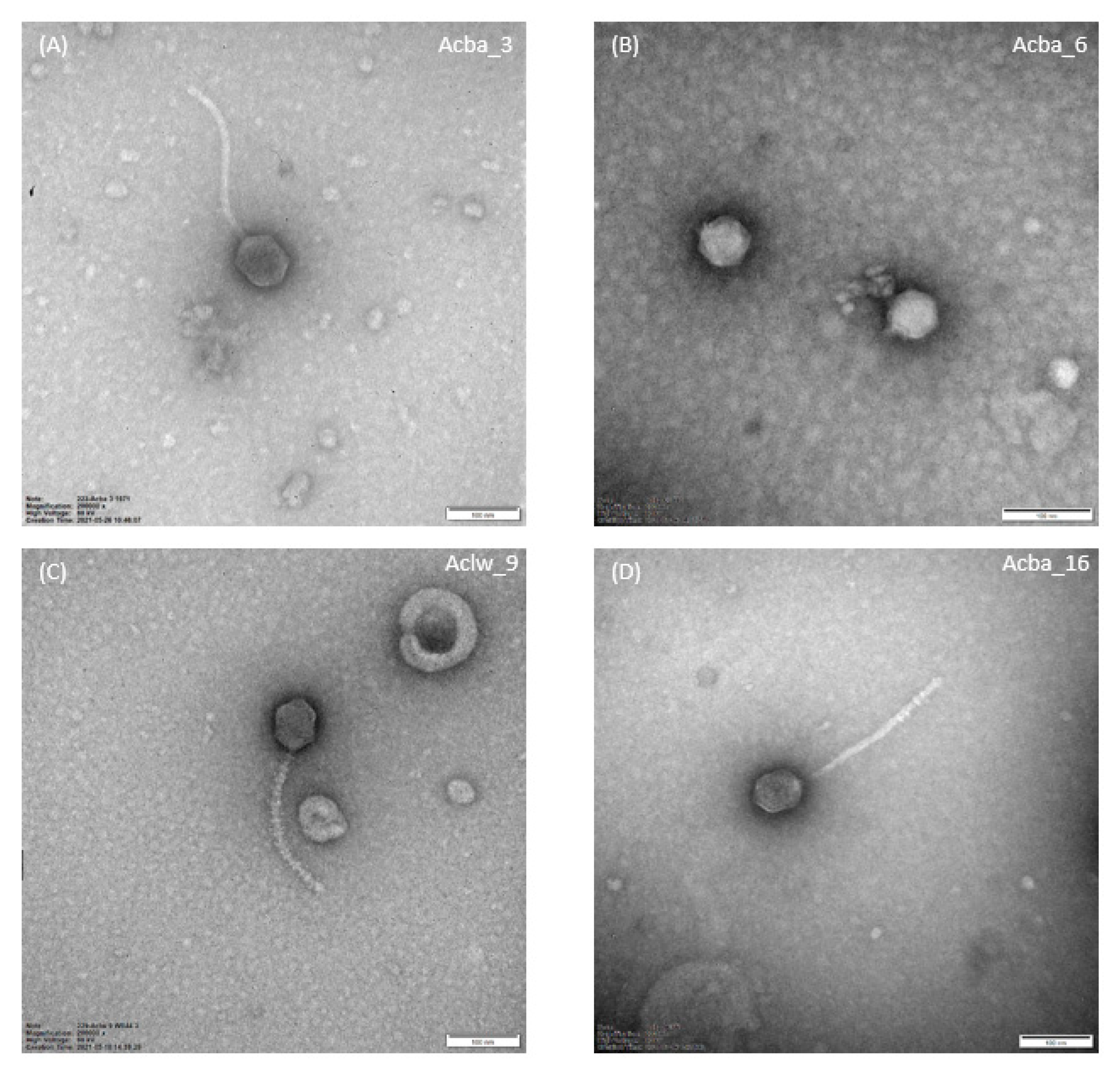
2.6. Transmission Electron Microscopy (TEM)
2.7. Isolation and Sequencing of Phage DNA
2.8. Phylogenetic Tree
3. Results
3.1. Bacterial Strains Used in the Study
3.2. Bacteriophages
3.2.1. General Characteristics
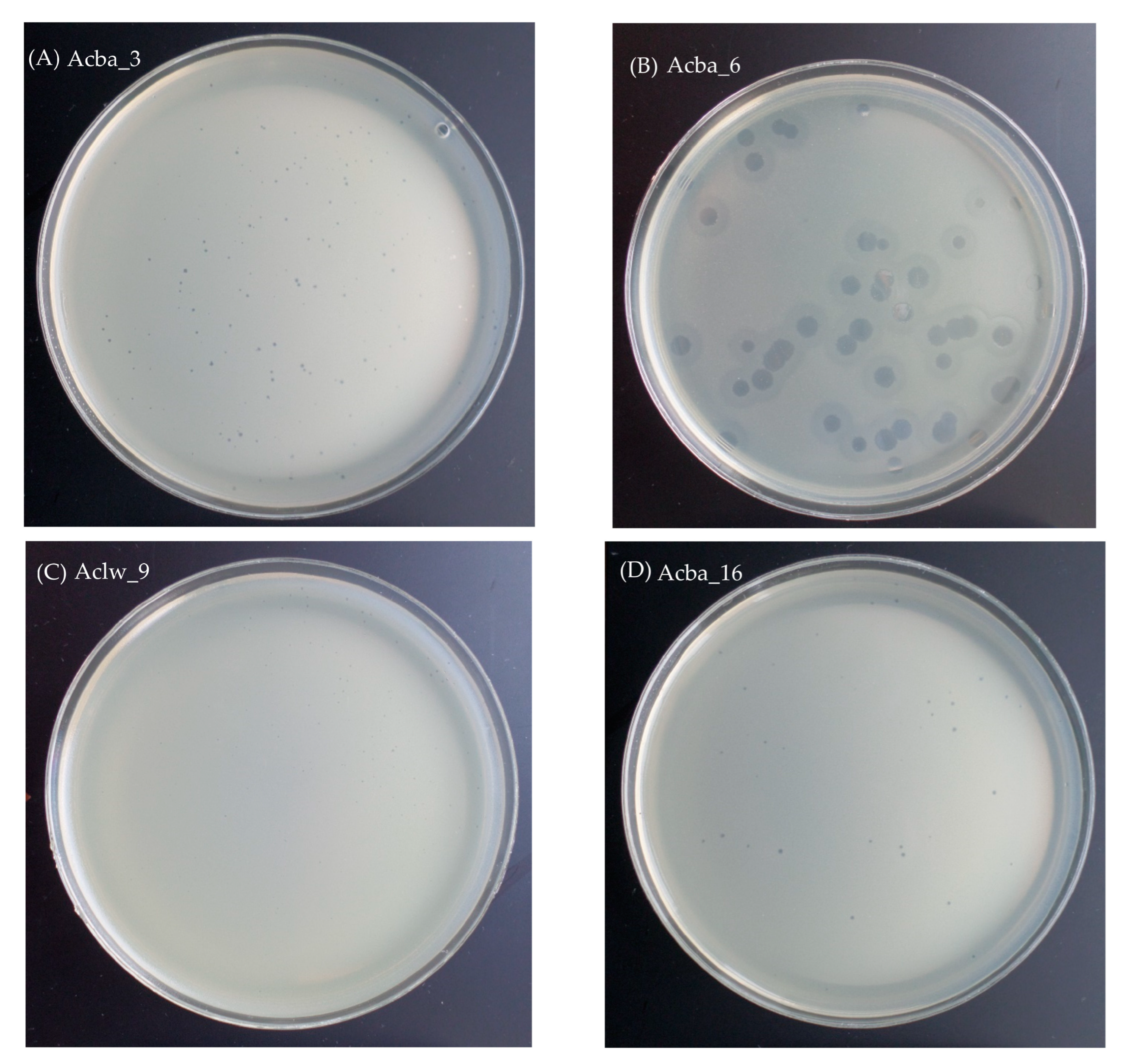
3.2.2. Morphology and Dimensions of Phages and Their Plaques
3.3. Investigating the Best MOI to Optimize Phage Amplification
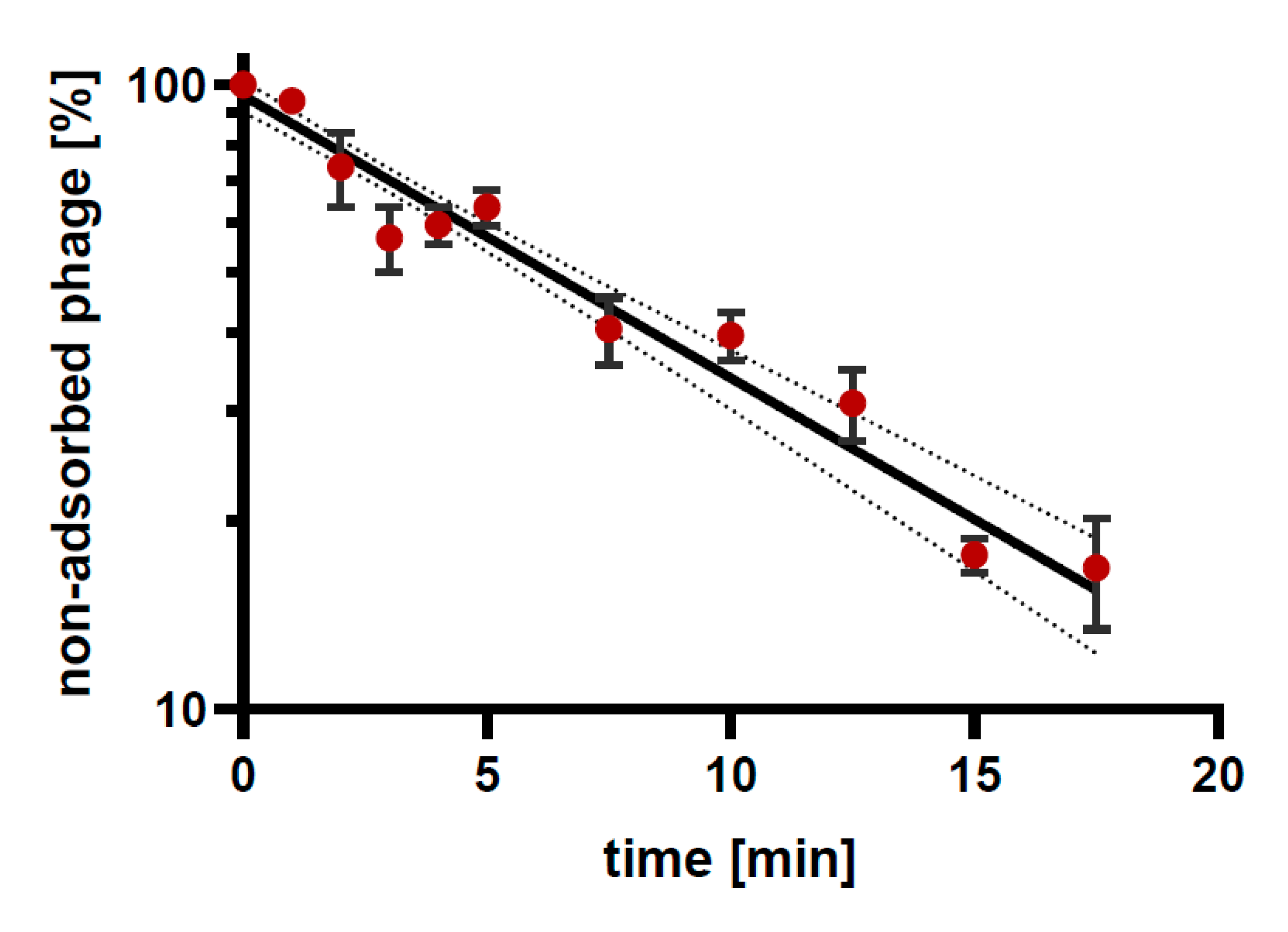
3.4. Phage Adsorption to Bacterial Cells
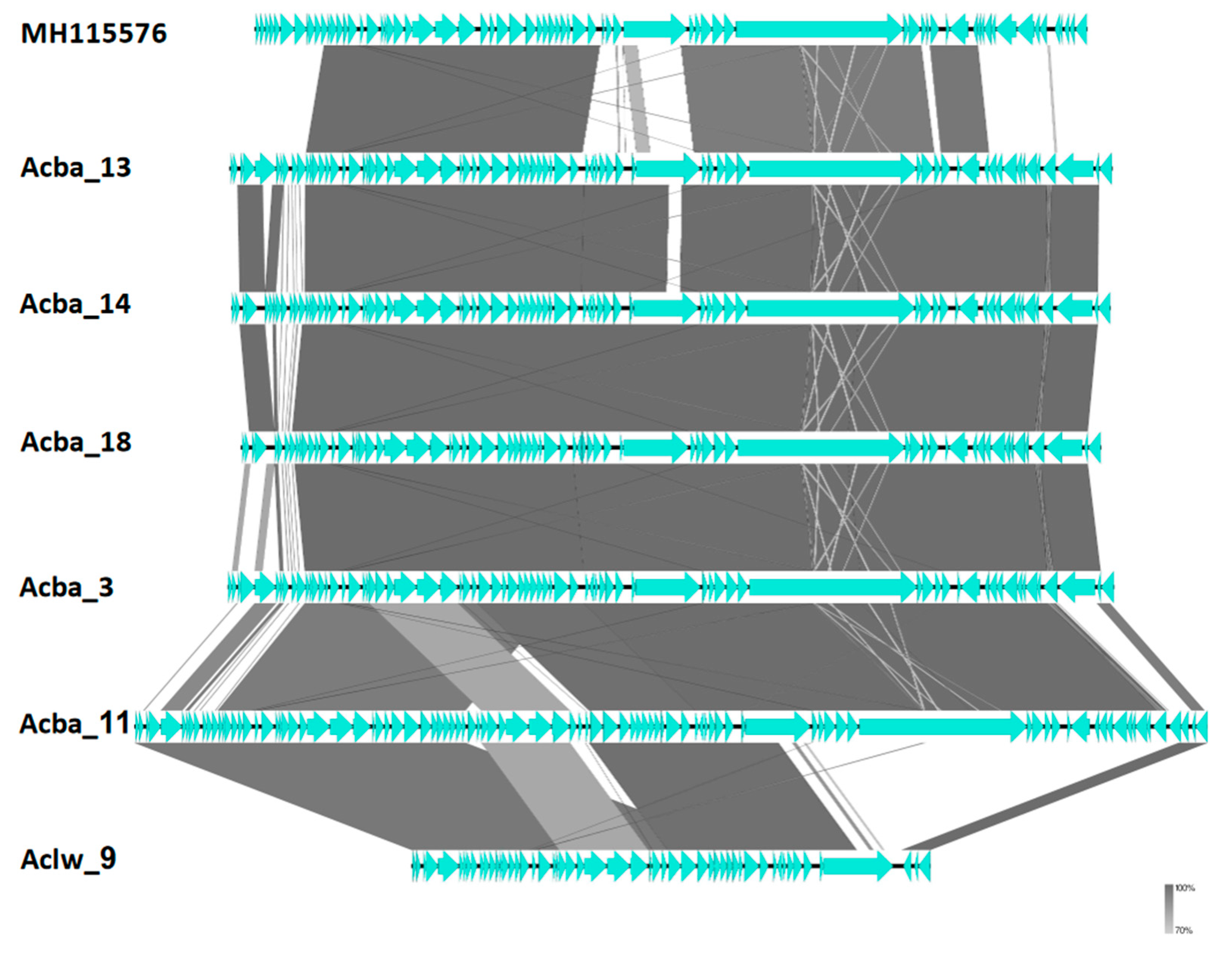
3.5. Genome Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Towner, K.J. Acinetobacter: An old friend, but a new enemy. J. Hosp. Infect. 2009, 7, 355–363. [Google Scholar] [CrossRef]
- Kuehn, B.M. Progress Against Antimicrobial Resistance Has Slipped. JAMA 2022, 328, 702. [Google Scholar] [CrossRef] [PubMed]
- Munk, P.; Brinch, C.; Møller, F.D.; Petersen, T.N.; Hendriksen, R.S.; Seyfarth, A.M.; Kjeldgaard, J.S.; Svendsen, C.A.; van Bunnik, B.; Berglund, F.; et al. Genomic analysis of sewage from 101 countries reveals global landscape of antimicrobial resistance. Nat. Commun. 2022, 13, 7251. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Available online: http://www.who.int/mediacentre/news/releases/2017/bacteria-antibiotics-needed/en (accessed on 14 November 2018).
- World Health Organization (WHO). Available online: https://www.who.int/emergencies/ten-threats-to-global-health-in-2019 (accessed on 18 February 2019).
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.H.; Wang, J.L.; Wen, F.S.; Chang, K.M.; Kuo, C.F.; Lin, C.H.; Luo, H.R.; Hung, C.H. Isolation and characterization of φkm18p, a novel lytic phage with therapeutic potential against extensively drug resistant Acinetobacter baumannii. PLoS ONE 2012, 7, e46537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, F.; Mi, Z.; Huang, Y.; Yuan, X.; Niu, W.; Wang, Y.; Hua, Y.; Fan, H.; Bai, C.; Tong, Y. Characterization, sequencing and comparative genomic analysis of vB_AbaM-IME-AB2, a novel lytic bacteriophage that infects multidrug-resistant Acinetobacter baumannii clinical isolates. BMC Microbiol. 2014, 5, 181. [Google Scholar] [CrossRef] [Green Version]
- Cisneros, J.; Rodriguez-Bano, J. Nosocomial bacteremia due to Acinetobacter baumannii: Epidemiology, clinical features and treatment. Clin. Microbiol. Infect. 2002, 8, 687–693. [Google Scholar] [CrossRef] [Green Version]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [Green Version]
- Al-Kadmy, I.M.S.; Ali, A.N.M.; Salman, I.M.A.; Khazaal, S.S. Molecular characterization of Acinetobacter baumannii isolated from Iraqi hospital environment. New Microbes New Infect. 2018, 21, 51–57. [Google Scholar] [CrossRef]
- Garnacho-Montero, J.; Ortiz-Leyba, C.; Fernández-Hinojosa, E.; Aldabó-Pallás, T.; Cayuela, A.; Marquez-Vácaro, J.A.; Garcia-Curiel, A.; Jiménez-Jiménez, F.J. Acinetobacter baumannii ventilator-associated pneumonia: Epidemiological and clinical findings. Intensive Care Med. 2005, 31, 649–655. [Google Scholar] [CrossRef]
- Khazaal, S.S.; Al-Saryi, N.; Ibrahim, S.A. Immunomodulation by Acinetobacter baumannii of endotracheal tube biofilm in ventilator-associated pneumonia. Meta Gene. 2020, 13, 100672. [Google Scholar] [CrossRef]
- Khazaal, S.S.; Al-Kadmy, I.M.; Aziz, S.N. Mechanism of pathogenesis in multidrug resistant Acinetobacter baumannii isolated from intensive care unit. Gene Rep. 2020, 18, 100557. [Google Scholar] [CrossRef]
- Hakemi Vala, M.; Hallajzadeh, M.; Hashemi, A.; Goudarzi, H.; Tarhani, M.; Sattarzadeh Tabrizi, M.; Bazmi, F. Detection of Ambler class A, B and D ß-lactamases among Pseudomonas aeruginosa and Acinetobacter baumannii clinical isolates from burn patients. Ann. Burns Fire Disasters 2014, 27, 8–13. [Google Scholar]
- Kareem, S.M.; Al-Kadmy, I.M.; Kazaal, S.S.; Ali, A.N.M.; Aziz, S.N.; Makharita, R.R.; Algammal, A.M.; Al-Rejaie, S.; Behl, T.; Batiha, G.E.S.; et al. Detection of gyra and parc mutations and prevalence of plasmid-mediated quinolone resistance genes in Klebsiella pneumoniae. Infect. Drug Resist. 2021, 14, 555. [Google Scholar] [CrossRef]
- Fernandes, R.; Amador, P.; Prudêncio, C. β-lactams: Chemical structure, mode of action and mechanisms of resistance. Rev. Med. Microbiol. 2013, 24, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Jean, S.S.; Hsueh, P.R. High burden of antimicrobial resistance in Asia. Int. J. Antimicrob. Agents. 2011, 37, 291–295. [Google Scholar] [CrossRef]
- Chopra, T.; Marchaim, D.; Awali, R.A.; Krishna, A.; Johnson, P.; Tansek, R.; Chaudary, K.; Lephart, P.; Slim, J.; Hothi, J.; et al. Epidemiology of bloodstream infections caused by Acinetobacter baumannii and impact of drug resistance to both carbapenems and ampicillin-sulbactam on clinical outcomes. Antimicrob. Agents Chemother. 2013, 12, 6270–6275. [Google Scholar] [CrossRef] [Green Version]
- Velkov, T.; Dai, C.; Ciccotosto, G.D.; Cappai, R.; Hoyer, D.; Li, J. Polymyxins for CNS infections: Pharmacology and neurotoxicity. Pharmacol. Ther. 2018, 181, 85–90. [Google Scholar] [CrossRef]
- Ibrahim, S.; Al-Saryi, N.; Al-Kadmy, I.M.S.; Aziz, S.N. Multidrug-resistant Acinetobacter baumannii as an emerging concern in hospitals. Mol. Biol. Rep. 2021, 48, 6987–6998. [Google Scholar] [CrossRef]
- Alaoui Mdarhri, H.; Benmessaoud, R.; Yacoubi, H.; Seffar, L.; Guennouni Assimi, H.; Hamam, M.; Boussettine, R.; Filali-Ansari, N.; Lahlou, F.A.; Diawara, I.; et al. Alternatives Therapeutic Approaches to Conventional Antibiotics: Advantages, Limitations and Potential Application in Medicine. Antibiotics 2022, 11, 1826. [Google Scholar] [CrossRef]
- Tan, Y.; Su, J.; Fu, M.; Zhang, H.; Zeng, H. Recent Advances in Phage-Based Therapeutics for Multi-Drug Resistant Acinetobacter baumannii. Bioengineering 2023, 10, 35. [Google Scholar] [CrossRef]
- Blasco, L.; Bleriot, I.; González de Aledo, M.; Fernández-García, L.; Pacios, O.; Oliveira, H.; López, M.; Ortiz-Cartagena, C.; Fernández-Cuenca, F.; Pascual, Á.; et al. Development of an Anti-Acinetobacter baumannii Biofilm Phage Cocktail: Genomic Adaptation to the Host. Antimicrob. Agents Chemother. 2022, 66, e0192321. [Google Scholar] [CrossRef]
- Grygorcewicz, B.; Roszak, M.; Golec, P.; Śleboda-Taront, D.; Łubowska, N.; Górska, M.; Jursa-Kulesza, J.; Rakoczy, R.; Wojciuk, B.; Dołęgowska, B. Antibiotics Act with vB_AbaP_AGC01 Phage against Acinetobacter baumannii in Human Heat-Inactivated Plasma Blood and Galleria mellonella Models. Int. J. Mol. Sci. 2020, 21, 4390. [Google Scholar] [CrossRef] [PubMed]
- Gordillo Altamirano, F.L.; Kostoulias, X.; Subedi, D.; Korneev, D.; Peleg, A.Y.; Barr, J.J. Phage-antibiotic combination is a superior treatment against Acinetobacter baumannii in a preclinical study. EBioMedicine 2022, 80, 104045. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Kuo, C.F.; Yeh, C.M.; Chen, J.R.; Cheng, M.F.; Hung, C.H. Efficacy of φkm18p phage therapy in a murine model of extensively drug-resistant Acinetobacter baumannii infection. Infect. Drug Resist. 2018, 11, 2301–2310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.; Dai, J.; Guo, M.; Li, J.; Zhou, X.; Li, F.; Gao, Y.; Qu, H.; Lu, H.; Jin, J.; et al. Pre-optimized phage therapy on secondary Acinetobacter baumannii infection in four critical COVID-19 patients. Emerg. Microbes Infect. 2021, 10, 612–618. [Google Scholar] [CrossRef]
- Wienhold, S.M.; Brack, M.C.; Nouailles, G.; Krishnamoorthy, G.; Korf, I.H.E.; Seitz, C.; Wienecke, S.; Dietert, K.; Gurtner, C.; Kershaw, O.; et al. Preclinical Assessment of Bacteriophage Therapy against Experimental Acinetobacter baumannii Lung Infection. Viruses 2021, 14, 33. [Google Scholar] [CrossRef]
- Tan, X.; Chen, H.; Zhang, M.; Zhao, Y.; Jiang, Y.; Liu, X.; Huang, W.; Ma, Y. Clinical Experience of Personalized Phage Therapy Against Carbapenem-Resistant Acinetobacter baumannii Lung Infection in a Patient with Chronic Obstructive Pulmonary Disease. Front. Cell. Infect. Microbiol. 2021, 11, 631585. [Google Scholar] [CrossRef]
- Allemailem, K.S.; Alnuqaydan, A.M.; Almatroudi, A.; Alrumaihi, F.; Aljaghwani, A.; Khalilullah, H.; Younus, H.; Khan, A.; Khan, M.A. Safety and Therapeutic Efficacy of Thymoquinone-Loaded Liposomes against Drug-Sensitive and Drug-Resistant Acinetobacter baumannii. Pharmaceutics 2021, 13, 677. [Google Scholar] [CrossRef]
- Khan, M.A.; Allemailem, K.S.; Maswadeh, H.; Younus, H. Glycosphingolipids (GSLs) from Sphingomonas paucimobilis Increase the Efficacy of Liposome-Based Nanovaccine against Acinetobacter baumannii-Associated Pneumonia in Immunocompetent and Immunocompromised Mice. Molecules 2022, 27, 7790. [Google Scholar] [CrossRef]
- Ślopek, S.; Durlakowa, I.; Weber-Dabrowska, B.; Dabrowski, M.; Kucharewicz-Krukowska, A. Results of bacteriophage treatment of suppurative bacterial infections. III. Detailed evaluation of the results obtained in further 150 cases. Arch. Immunol. Ther. Exp. 1984, 32, 317–335. [Google Scholar]
- Cieślik, M.; Harhala, M.; Orwat, F.; Dąbrowska, K.; Górski, A.; Jończyk-Matysiak, E. Two Newly Isolated Enterobacter-Specific Bacteriophages: Biological Properties and Stability Studies. Viruses 2022, 14, 1518. [Google Scholar] [CrossRef] [PubMed]
- Gratia, A. Des relations numeriques entre bacteries lysogenes et particles de bacteriophage. Ann. Inst. Pasteur. 1936, 57, 652–676. [Google Scholar]
- Adams, M.H. Enumeration of bacteriophage particles. In The Bacteriophages; Inter Science Publisher: New York, NY, USA, 1959. [Google Scholar]
- Kropinski, A.M. Measurement of the rate of attachment of bacteriophage to cells. In Bacteriophages. Methods in Molecular Biology; Clokie, M.R., Kropinski, A.M., Eds.; Humana Press: Totowa, NJ, USA, 2009; Volume 501, pp. 151–155. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Antipov, D.; Korobeynikov, A.; McLean, J.S.; Pevzner, P.A. hybrid SPAdes: An algorithm for hybrid assembly of short and long reads. Bioinformatics 2016, 32, 1009–1015. [Google Scholar] [CrossRef] [Green Version]
- Prjibelski, A.D.; Vasilinetc, I.; Bankevich, A.; Gurevich, A.; Krivosheeva, T.; Nurk, S.; Pham, S.; Korobeynikov, A.; Lapidus, A.; Pevzner, P.A. ExSPAnder: A universal repeat resolver for DNA fragment assembly. Bioinformatics 2014, 30, i293–i301. [Google Scholar] [CrossRef] [Green Version]
- Vasilinetc, I.; Prjibelski, A.D.; Gurevich, A.; Korobeynikov, A.; Pevzner, P.A. Assembling short reads from jumping libraries with large insert sizes. Bioinformatics 2015, 31, 3262–3268. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.; Korobeynikov, A.; Lapidus, A.; Prjibelsky, A.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling Genomes and Mini-metagenomes from Highly Chimeric Reads. In Lecture Notes in Computer Science; Springer: Berlin/Heidelberg, Germany, 2013; Volume 821, pp. 158–170. [Google Scholar]
- Cuccuru, G.; Orsini, M.; Pinna, A.; Sbardellati, A.; Soranzo, N.; Travaglione, A.; Uva, P.; Zanetti, G.; Fotia, G. Orione, a web-based framework for NGS analysis in microbiology. Bioinformatics 2014, 30, 1928–1929. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka. Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, H.; Domingues, R.; Evans, B.; Sutton, J.M.; Adriaenssens, E.M.; Turner, D. Genomic diversity of bacteriophages infecting the genus Acinetobacter. Viruses 2022, 14, 181. [Google Scholar] [CrossRef]
- Adriaenssens, E.; Brister, J.R. How to Name and Classify Your Phage: An Informal Guide. Viruses 2017, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Turner, D.; Ackermann, H.W.; Kropinski, A.M.; Lavigne, R.; Sutton, J.M.; Reynolds, D.M. Comparative analysis of 37 Acinetobacter bacteriophages. Viruses 2017, 10, 5. [Google Scholar] [CrossRef]
- Suh, G.A.; Lodise, T.P.; Tamma, P.D.; Knisely, J.M.; Alexander, J.; Aslam, S.; Barton, K.D.; Bizzell, E.; Totten, K.M.C.; Campbell, J.; et al. Antibacterial Resistance Leadership Group. Considerations for the Use of Phage Therapy in Clinical Practice. Antimicrob. Agents Chemother. 2022, 66, e02071-21. [Google Scholar] [CrossRef]
- Liu, M.; Hernandez-Morales, A.; Clark, J.; Le, T.; Biswas, B.; Bishop-Lilly, K.A.; Henry, M.; Quinones, J.; Voegtly, L.J.; Cer, R.Z.; et al. Comparative genomics of Acinetobacter baumannii and therapeutic bacteriophages from a patient undergoing phage therapy. Nat. Commun. 2022, 13, 3776. [Google Scholar] [CrossRef]
- Nemec, A.; Krizova, L.; Maixnerova, M.; van der Reijden, T.J.; Deschaght, P.; Passet, V.; Vaneechoutte, M.; Brisse, S.; Dijkshoorn, L. Genotypic and phenotypic characterization of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex with the proposal of Acinetobacter pittii sp. nov. (formerly Acinetobacter genomic species 3) and Acinetobacter nosocomialis sp. nov. (formerly Acinetobacter genomic species 13TU). Res. Microbiol. 2011, 162, 393–404. [Google Scholar]
- Rodrigues, D.L.N.; Morais-Rodrigues, F.; Hurtado, R.; Dos Santos, R.G.; Costa, D.C.; Barh, D.; Ghosh, P.; Alzahrani, K.J.; Soares, S.C.; Ramos, R.; et al. Pan-Resistome Insights into the Multidrug Resistance of Acinetobacter baumannii. Antibiotics 2021, 10, 596. [Google Scholar] [CrossRef]
- Álvarez, V.E.; Quiroga, M.P.; Galán, A.V.; Vilacoba, E.; Quiroga, C.; Ramírez, M.S.; Centrón, D. Crucial Role of the Accessory Genome in the Evolutionary Trajectory of Acinetobacter baumannii Global Clone 1. Front. Microbiol. 2020, 11, 342. [Google Scholar] [CrossRef] [Green Version]
- Holt, K.; Kenyon, J.J.; Hamidian, M.; Schultz, M.B.; Pickard, D.J.; Dougan, G.; Hall, R. Five decades of genome evolution in the globally distributed, extensively antibiotic-resistant Acinetobacter baumannii global clone 1. Microb. Genom. 2016, 2, e000052. [Google Scholar] [CrossRef]
- Lakshminarasimhan, A. Prophage induction therapy: Activation of the lytic phase in prophages for the elimination of pathogenic bacteria. Med. Hypotheses 2022, 169, 110980. [Google Scholar] [CrossRef]
- Jin, J.; Li, Z.J.; Wang, S.W.; Wang, S.M.; Huang, D.H.; Li, Y.H.; Ma, Y.Y.; Wang, J.; Liu, F.; Chen, X.D.; et al. Isolation and characterization of ZZ1, a novel lytic phage that infects Acinetobacter baumannii clinical isolates. BMC Microbiol. 2012, 12, 156. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Hu, K.; Xie, Y.; Liu, Y.; Mu, D.; Guo, H.; Zhang, Z.; Zhang, Y.; Chang, D.; Shi, Y.A. Novel Phage PD-6A3, and Its Endolysin Ply6A3, With Extended Lytic Activity Against Acinetobacter baumannii. Front. Microbiol. 2019, 9, 3302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wintachai, P.; Surachat, K.; Chaimaha, G.; Septama, A.W.; Smith, D.R. Isolation and Characterization of a Phapecoctavirus Infecting Multidrug-Resistant Acinetobacter baumannii in A549 Alveolar Epithelial Cells. Viruses 2022, 14, 2561. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, X.; Hua, X.; Yu, Y.; Leptihn, S.; Loh, B. Therapeutic evaluation of the Acinetobacter baumannii phage Phab24 for clinical use. Virus. Res. 2022, 320, 198889. [Google Scholar] [CrossRef] [PubMed]
- Popova, A.V.; Shneider, M.M.; Arbatsky, N.P.; Kasimova, A.A.; Senchenkova, S.N.; Shashkov, A.S.; Dmitrenok, A.S.; Chizhov, A.O.; Mikhailova, Y.V.; Shagin, D.A.; et al. Specific Interaction of Novel Friunavirus Phages Encoding Tailspike Depolymerases with Corresponding Acinetobacter baumannii Capsular Types. J. Virol. 2020, 95, e01714-20. [Google Scholar]
- Vukotic, G.; Obradovic, M.; Novovic, K.; Di Luca, M.; Jovcic, B.; Fira, D.; Neve, H.; Kojic, M.; McAuliffe, O. Characterization, Antibiofilm, and Depolymerizing Activity of Two Phages Active on Carbapenem-Resistant Acinetobacter baumannii. Front. Med. 2020, 7, 426. [Google Scholar] [CrossRef]
- Timoshina, O.Y.; Shneider, M.M.; Evseev, P.V.; Shchurova, A.S.; Shelenkov, A.A.; Mikhaylova, Y.V.; Sokolova, O.S.; Kasimova, A.A.; Arbatsky, N.P.; Dmitrenok, A.S.; et al. Novel Acinetobacter baumannii Bacteriophage Aristophanes Encoding Structural Polysaccharide Deacetylase. Viruses 2021, 13, 1688. [Google Scholar] [CrossRef]
- Omata, K.; Hibi, N.; Nakano, S.; Komoto, S.; Sato, K.; Nunokawa, K.; Amano, S.; Ueda, K. Distribution and genome structures of temperate phages in acetic acid bacteria. Sci. Rep. 2021, 11, 21567. [Google Scholar] [CrossRef]
- Badawy, S.; Pajunen, M.I.; Haiko, J.; Baka, Z.A.M.; Abou-Dobara, M.I.; El-Sayed, A.K.A.; Skurnik, M. Identification and Functional Analysis of Temperate Siphoviridae Bacteriophages of Acinetobacter baumannii. Viruses 2020, 12, 604. [Google Scholar] [CrossRef]
- Oppenheim, A.B.; Kobiler, O.; Stavans, J.; Court, D.L.; Adhya, S. Switches in Bacteriophage Lambda Development. Annu. Rev. Genet. 2005, 39, 409–429. [Google Scholar] [CrossRef]
- Baek, K.; Svenningsen, S.; Eisen, H.; Sneppen, K.; Brown, S. Single-cell analysis of lambda immunity regulation. J. Mol. Biol. 2003, 334, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Hammerl, J.A.; Jäckel, C.; Funk, E.; Pinnau, S.; Mache, C.; Hertwig, S. The diverse genetic switch of enterobacterial and marine telomere phages. Bacteriophage 2016, 6, e1148805. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | Source of Isolation | Sensitivity to Tested A. baumannii Phages |
|---|---|---|
| P. aeruginosa 6111 | swab from the fistula | no |
| P. aeruginosa 1790/1 | sputum | no |
| K. pneumoniae 8385 | urine | no |
| E. coli 9608 | vagina | no |
| E. coli 8069 | urine | no |
| E. cloacae 2084 | urine | no |
| No. | Phage Symbol | ICTV Symbol | Source and Year of Isolation | Lytic Spectrum | Host Strain |
|---|---|---|---|---|---|
| 1. | Acba_1 | Acinetobacter phage vB_AbaS-1 | city ditch water (Sroda Slaska, Lower Silesia, Poland, 2007) | 11% | A. baumannii K26905 (main) * A. baumannii: 703, 1326, 1971 (alternative) ** |
| 2. | Acba_3 | Acinetobacter phage vB_AbaS-3 | water sample (Kluczowa, Lower Silesia, Poland, 2018) | 47% | A. baumannii 1971 |
| 3. | Acba_4 | Acinetobacter phage vB_AbaS-4 | water sample (Drobnice, Lodz Province, Poland, 2018) | 30% | A. baumannii 1971 |
| 4. | Acba_6 | Acinetobacter phage vB_AbaP-7 | hospital sewage (delivery and operating block) Regional Specialist Hospital in Wroclaw (Wroclaw, Lower Silesia, Poland, 2013) | 19% | A. baumannii 3940 |
| 5. | Aclw_8 | Acinetobacter phage vB_AlwS-8 | sewage treated from a sewage treatment plant (Grodzisk Mazowiecki, Masovian, Poland, 2014) | 11% | A. lwoffi WII44.3 (main) A. baumannii: 703, 1326, 1971 (alternative) |
| 6. | Aclw_9 | Acinetobacter phage vB_AlwS-9 | water taken from the Water Treatment Plant (Grodzisk Mazowiecki, Masovian, Poland, 2015) | 72% | A. lwoffi WII44.3 (main) A. baumannii: 703, 1326, 1971 (alternative) |
| 7. | Acba_11 | Acinetobacter phage vB_AbaS-11 | water sample (Walichnowy, Lodz Province, Poland, 2018) | 62% | A. baumannii 1326 |
| 8. | Acba_13 | Acinetobacter phage vB_AbaS-13 | river water (Olawa, Lower Silesia, Poland, 2018) | 17% | A. baumannii 1326 |
| 9. | Acba_14 | Acinetobacter phage vB_AbaS-14 | water sample (Walichnowy, Lodz Province, Poland, 2018) | 21% | A. baumannii 1971 |
| 10. | Acba_15 | Acinetobacter phage vB_AbaS-15 | water sample (Lyskornia, Lodz Province, Poland, 2018) | 75% | A. baumannii 703 (main) A. baumannii: 1326, 1971 (alternative) |
| 11. | Acba_16 | Acinetobacter phage vB_AbaS-16 | water sample (Lyskornia, Lodz Province, Poland, 2018) | 13% | A. baumannii 703 (main) A. baumannii: 1326, 1971 (alternative) |
| 12. | Acba_18 | Acinetobacter phage vB_AbaS-18 | water from the drainage ditch (Olawa, Lower Silesia, Poland, 2018) | 11% | A. baumannii 703 (main) A. baumannii: 1326, 1971 (alternative) |
| Phage Symbol | Total Dimension [nm] | Capsid Length [nm] | Capsid Width [nm] | Capsid Diagonal [nm] | Tail Length [nm] | Tail Width [nm] |
|---|---|---|---|---|---|---|
| Acba_1 | 316.9 | 64.8 | 64.2 | 67.6 | 252.1 | 10.7 |
| Acba_3 | 331.5 | 82.8 | 65.4 | 77.2 | 248.7 | 10.1 |
| Acba_4 | 292 | 68.5 | 54.6 | 65.7 | 223.5 | 9.8 |
| Acba_6 | 56.9 | 56.9 | 58.6 | n/a | n/a | n/a |
| Aclw_8 | 308.8 | 71.5 | 56.3 | 67.2 | 237.3 | 9.7 |
| Aclw_9 | 307.3 | 76.1 | 59.7 | 67.1 | 231.2 | 10.3 |
| Acba_11 | 315.9 | 67.9 | 56.1 | 65.9 | 248 | 9.4 |
| Acba_13 | 329 | 76.2 | 62.8 | 75.2 | 252.8 | 10.3 |
| Acba_14 | 316 | 78.7 | 65.8 | 74.5 | 237.3 | 9.3 |
| Acba_15 | 322.5 | 78.3 | 61.6 | 73.3 | 244.2 | 10.3 |
| Acba_16 | 319.4 | 73.6 | 58.3 | 68.8 | 245.8 | 9.6 |
| Acba_18 | 297.1 | 64.6 | 52.8 | 61.1 | 232.5 | 9.4 |
| Phage Symbol | Phage Titer [PFU/mL] | MOI | Phage Titer after Incubation [PFU/mL] |
|---|---|---|---|
| Acba_1 | 102 | 0.001 | 7.89 × 106 |
| Acba_3 | 102 | 0.001 | 9.76 × 106 |
| Acba_4 | 104 | 0.1 | 4.73 × 107 |
| Acba_6 | 102 | 0.001 | 2.31 × 108 |
| Aclw_8 | 104 | 0.1 | 1.86 × 107 |
| Aclw_9 | 105 | 1 | 5.16 × 107 |
| Acba_11 | 104 | 0.1 | 4.74 × 107 |
| Acba_13 | 105 | 1 | 9.97 × 106 |
| Acba_14 | 105 | 1 | 4.73 × 107 |
| Acba_15 | 106 | 10 | 6.16 × 107 |
| Acba_16 | 105 | 1 | 4.61 × 107 |
| Acba_18 | 105 | 1 | 7.71 × 107 |
| Adsorption Rate Constant [mL/min] | |
|---|---|
| Acba_1 | 1.6 × 10−9 |
| Acba_3 | 1.2 × 10−9 |
| Acba_4 | 1.33 × 10−8 |
| Acba_6 | 5.76 × 10−9 |
| Aclw_8 | 1.04 × 10−9 |
| Aclw_9 | 2.61 × 10−9 |
| Acba_11 | 4.6 × 10−9 |
| Acba_13 | 1.11 × 10−9 |
| Acba_14 | 4.8 × 10−9 |
| Acba_15 | 2.6 × 10−9 |
| Acba_16 | 1.66 × 10−8 |
| Acba_18 | 5.6 × 10−9 |
| Phage Symbol | Genome Length [bp] | No. of ORFs | Strand [%] | Described Protein [%] | Start Codons [%] | |||
|---|---|---|---|---|---|---|---|---|
| Positive | Negative | ATG | TTG | GTG | ||||
| Acba_1 | 50,696 | 77 | 79.2 | 20.8 | 51.9 | 90.9 | 2.60 | 5.19 |
| Acba_3 | 55,181 | 72 | 80.6 | 19.4 | 52.8 | 97.2 | 1.39 | 1.39 |
| Acba_4 | 57,429 | 77 | 81.8 | 18.2 | 51.9 | 93.5 | 2.60 | 3.90 |
| Acba_6 | 42,776 | 52 | 100.0 | 0.0 | 65.4 | 98.1 | 1.92 | 0.00 |
| Aclw_8 | 47,018 | 70 | 84.3 | 15.7 | 45.7 | 94.3 | 1.43 | 4.29 |
| Aclw_9 | 32,415 | 53 | 92.5 | 7.5 | 41.5 | 94.3 | 1.89 | 3.77 |
| Acba_11 | 67,052 | 96 | 84.4 | 15.6 | 47.9 | 95.8 | 1.04 | 3.13 |
| Acba_13 | 55,385 | 73 | 80.8 | 19.2 | 49.3 | 98.6 | 1.37 | 0.00 |
| Acba_14 | 54,941 | 74 | 81.1 | 18.9 | 44.6 | 94.6 | 2.70 | 2.70 |
| Acba_15 | 60,424 | 81 | 80.2 | 19.8 | 53.1 | 95.1 | 2.47 | 2.47 |
| Acba_16 | 51,595 | 64 | 81.3 | 18.8 | 50.0 | 95.3 | 1.56 | 3.13 |
| Acba_18 | 53,751 | 70 | 80.0 | 20.0 | 48.6 | 98.6 | 1.43 | 0.00 |
| Presented Phage | Most Similar and Representative Phages | Subfamily (Proposed) | Taxonomy (Proposed) | Cluster | |
|---|---|---|---|---|---|
| Acc. no. | NCBI Description | ||||
| Acba_1 Acba_16 Acba_4 Acba_15 Aclw_8 | NC_041866 NC_019541 | Acinetobacter phage YMC11/11/R3177 Acinetobacter phage YMC/09/02/B1251_ABA_BP | Junivirinae | Vieuvirus | F3 |
| Acba_3 Aclw_9 Acba_18 Acba_14 Acba_11 Acba_13 | MH115576 | Acinetobacter phage AM106 | Junivirinae | Shemyakinvirus | F1 |
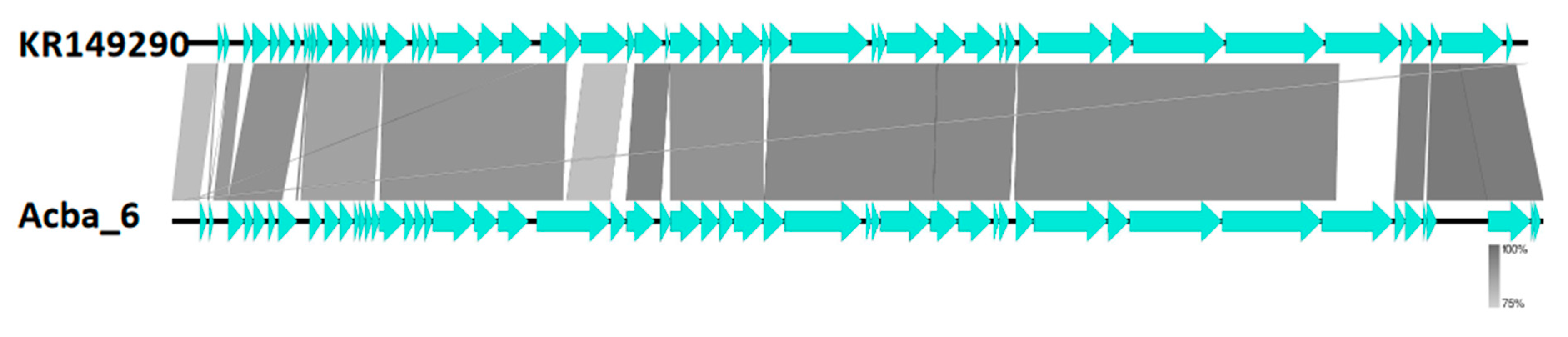
| Acba_6 | MN604239 NC_042007 KR149290 | Acinetobacter phage vB_AbaP_APK87 Acinetobacter phage vB_ApiP_P2 Acinetobacter phage Fri1 | Beijernickvirinae | Friunavirus | E2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bagińska, N.; Harhala, M.A.; Cieślik, M.; Orwat, F.; Weber-Dąbrowska, B.; Dąbrowska, K.; Górski, A.; Jończyk-Matysiak, E. Biological Properties of 12 Newly Isolated Acinetobacter baumannii-Specific Bacteriophages. Viruses 2023, 15, 231. https://doi.org/10.3390/v15010231
Bagińska N, Harhala MA, Cieślik M, Orwat F, Weber-Dąbrowska B, Dąbrowska K, Górski A, Jończyk-Matysiak E. Biological Properties of 12 Newly Isolated Acinetobacter baumannii-Specific Bacteriophages. Viruses. 2023; 15(1):231. https://doi.org/10.3390/v15010231
Chicago/Turabian StyleBagińska, Natalia, Marek Adam Harhala, Martyna Cieślik, Filip Orwat, Beata Weber-Dąbrowska, Krystyna Dąbrowska, Andrzej Górski, and Ewa Jończyk-Matysiak. 2023. "Biological Properties of 12 Newly Isolated Acinetobacter baumannii-Specific Bacteriophages" Viruses 15, no. 1: 231. https://doi.org/10.3390/v15010231
APA StyleBagińska, N., Harhala, M. A., Cieślik, M., Orwat, F., Weber-Dąbrowska, B., Dąbrowska, K., Górski, A., & Jończyk-Matysiak, E. (2023). Biological Properties of 12 Newly Isolated Acinetobacter baumannii-Specific Bacteriophages. Viruses, 15(1), 231. https://doi.org/10.3390/v15010231