
Unique Structure and Distinctive Properties of the Ancient and Ubiquitous Gamma-Type Envelope Glycoprotein

Abstract
:1. Introduction
1.1. Retroviruses with Gamma-Type Env Proteins
1.2. Variations on the Gamma-Type Env Are Also Found in Some Non-Retroviruses
1.3. Gamma-Type Env Encoded by Endogenous Retroviruses
1.4. Exaptation of ERV-Encoded Gamma-Type Env
1.4.1. Syncytins
1.4.2. Retrovirus Envs as Host-Encoded Resistance Genes
1.5. Significance
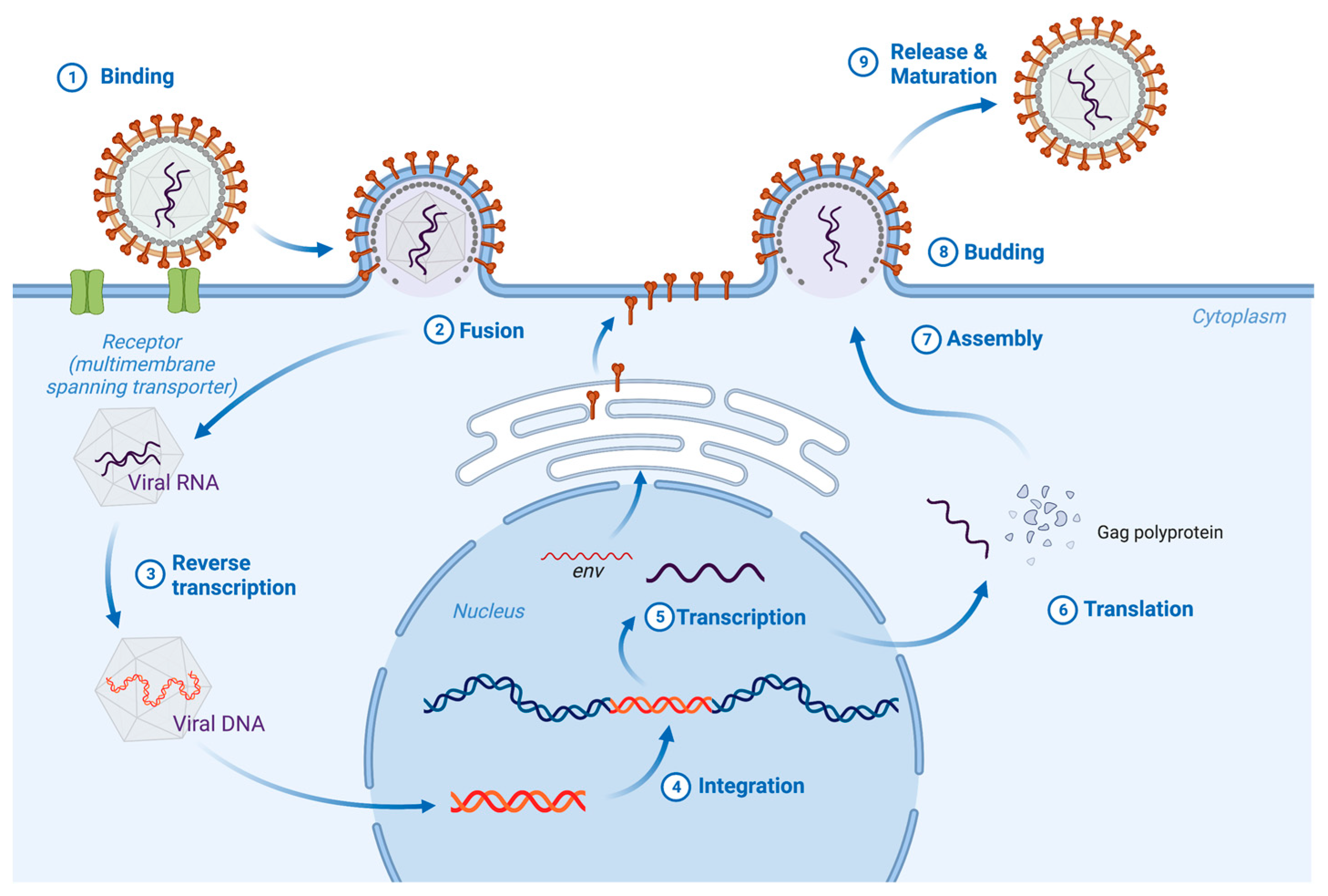
2. Biogenesis
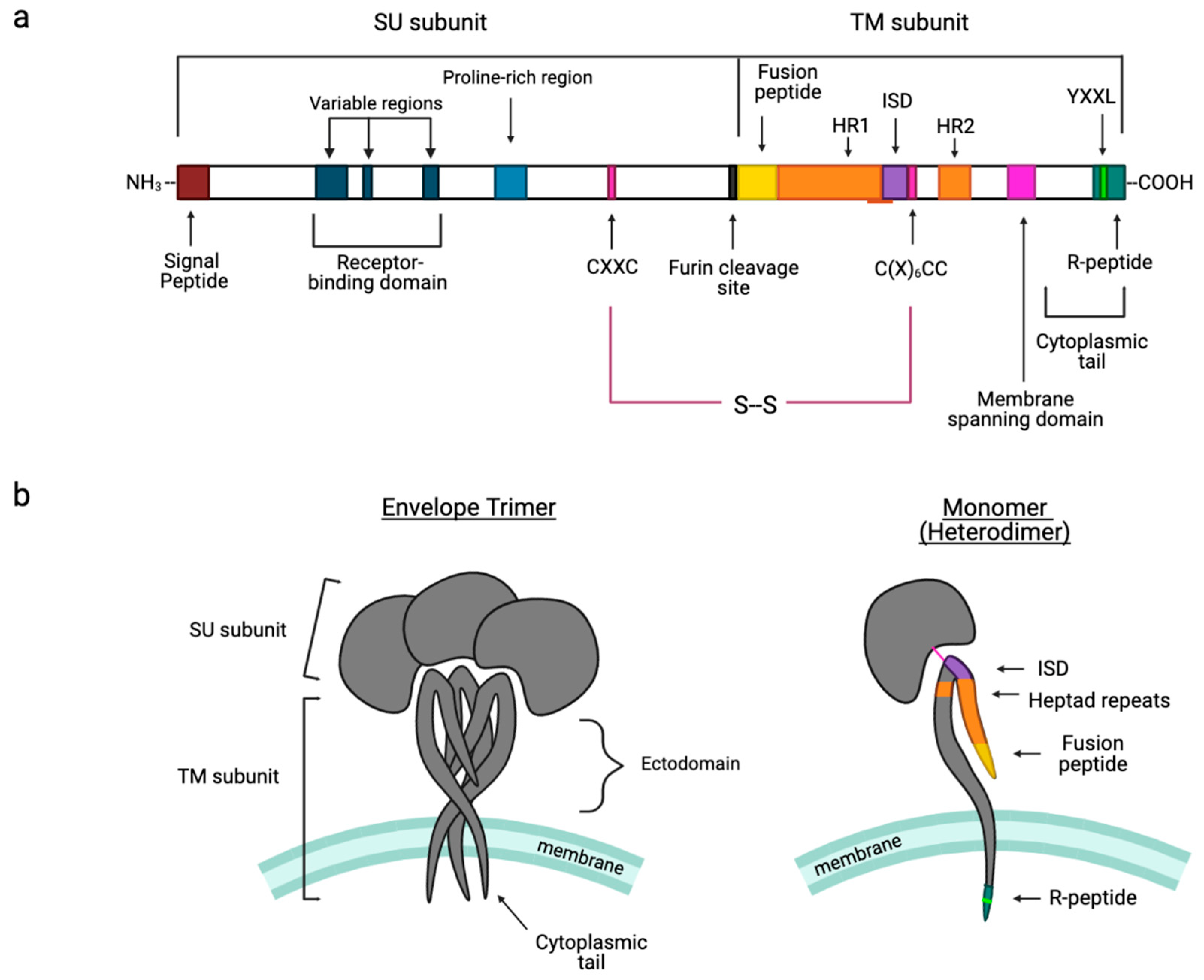
3. Hallmark Features of Gamma-Type Envs
3.1. Signal Peptide
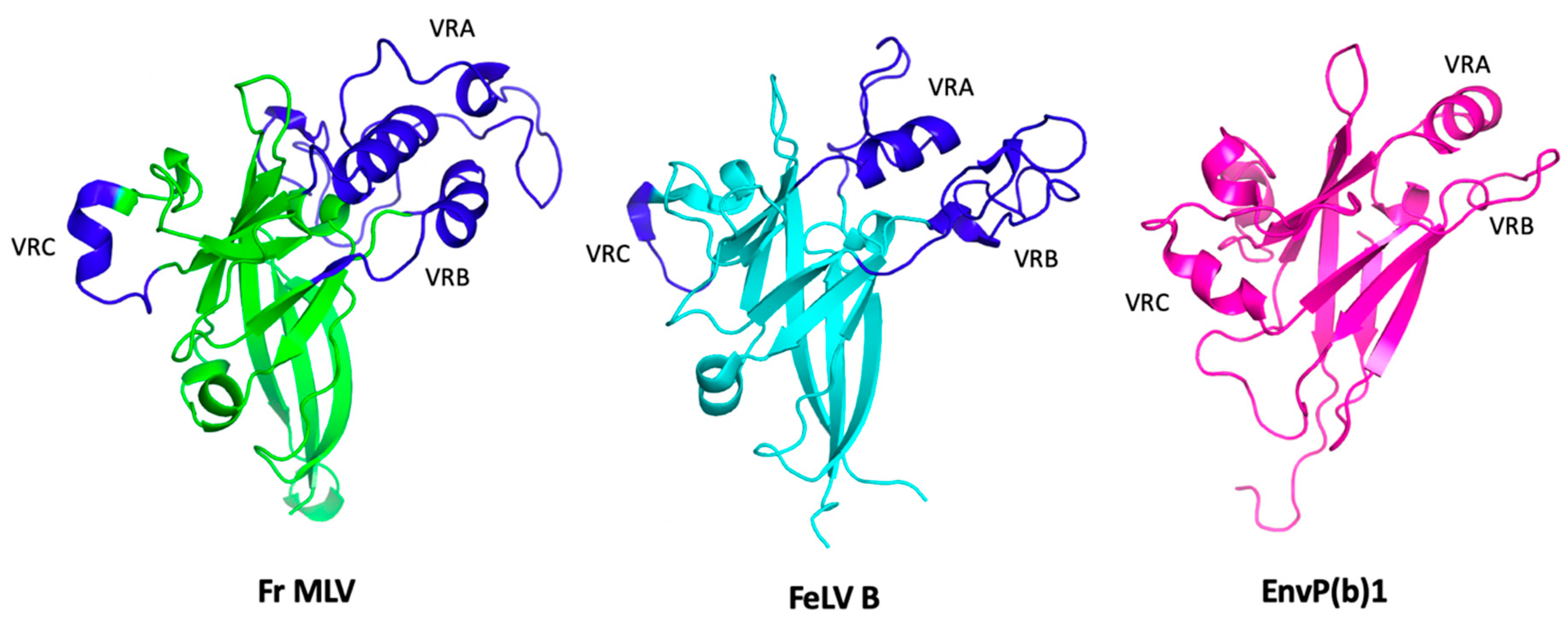
3.2. Receptor Binding Domain (RBD)
3.3. Proline-Rich Region (PRR)
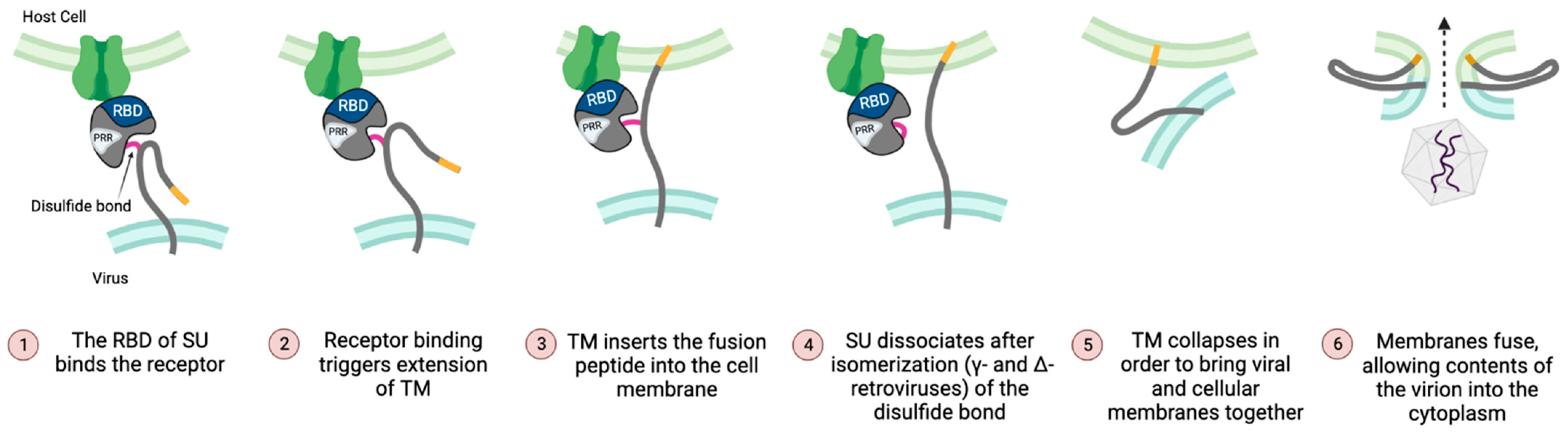
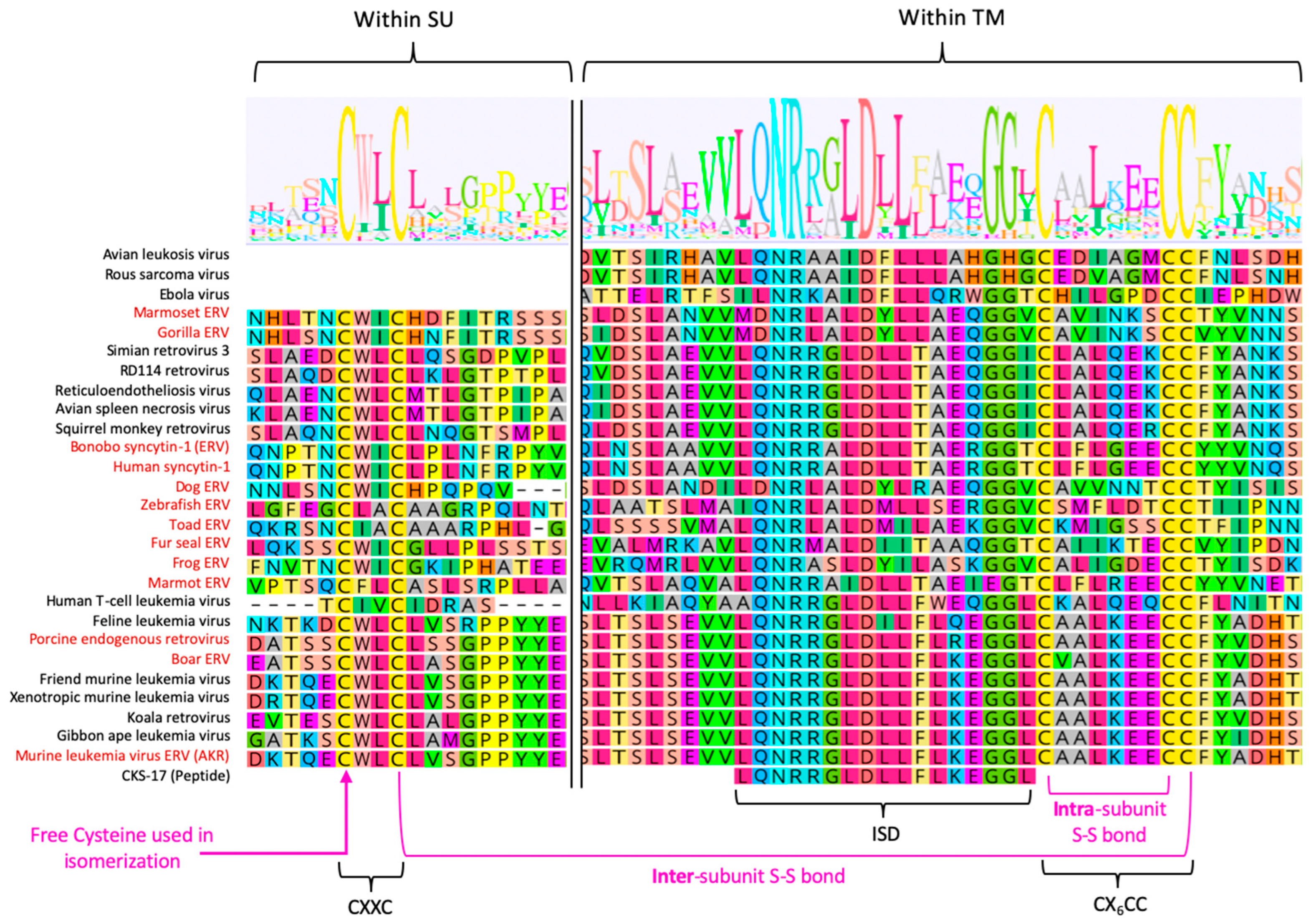
3.4. Intersubunit Disulfide Bond and the CXXC(SU) Motifs
3.5. Fusion Peptide
3.6. Heptad Repeats
3.7. Immunosuppressive Domain (ISD) and the CX6CC Motif
3.8. Cytoplasmic Tail
3.9. R-Peptide
4. Receptor Interactions of Gamma-Type Envs
4.1. The RBD Regulates Both Initiation and Completion of Fusion
4.2. Gamma-Type Env Preferentially Use SLC Superfamily Transporters as Entry Receptors
4.3. Recent Structural Insights into the SU-Receptor Interaction
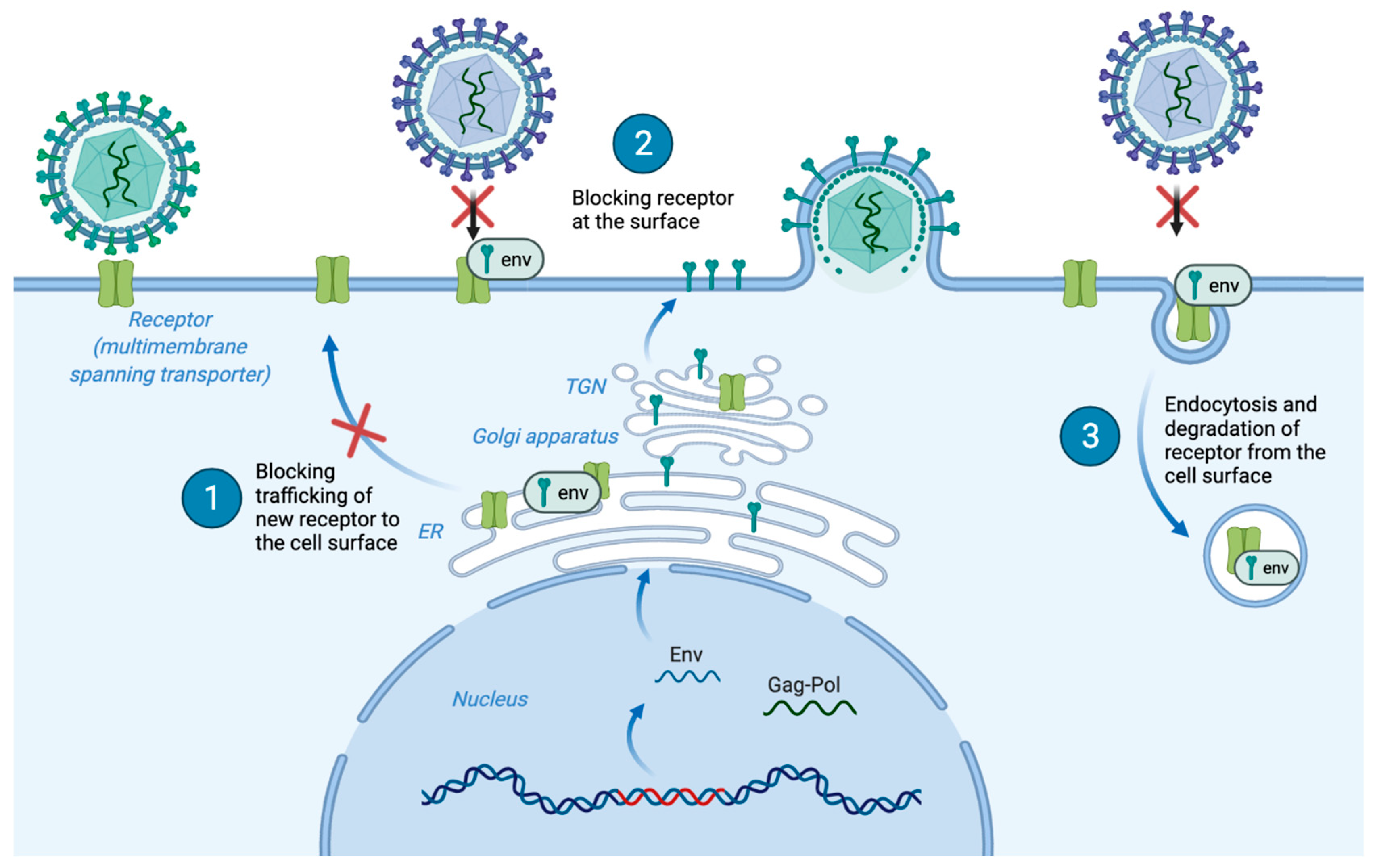
4.4. Receptor Interference
5. Summary and Outlook
5.1. Structural Insights into the Distinctive Entry and Fusion Mechanisms of Gamma-Type Env
5.2. Conservation of the ISD Motif
5.3. Receptor Bias
5.4. Virus-Host “Arms Races” and the Evolution of Individual SLC Receptors
5.5. The Link between Entry and Post-Entry
5.6. Env Trafficking and Receptor-Interference
5.7. Evolutionary Cooption of ERV-Encoded Gamma-Type Env Proteins
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coffin, J.; Blomberg, J.; Fan, H.; Gifford, R.; Hatziioannou, T.; Lindemann, D.; Mayer, J.; Stoye, J.; Tristem, M.; Johnson, W. ICTV Virus Taxonomy Profile: Retroviridae 2021. J. Gen. Virol. 2021, 102, 1712. [Google Scholar] [CrossRef] [PubMed]
- Sommerfelt, M.A.; Weiss, R.A. Receptor interference groups of 20 retroviruses plating on human cells. Virology 1990, 176, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Henzy, J.E.; Johnson, W. Pushing the endogenous envelope. Philos Trans. R Soc. Lond B Biol. Sci. 2013, 368, 20120506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wang, X.; Liao, M.-E.; Song, Y.; Zhang, Y.-Y.; Cui, J. Evolution and Genetic Diversity of the Retroviral Envelope in Anamniotes. J. Virol. 2022, 96, e0207221. [Google Scholar] [CrossRef] [PubMed]
- Sonigo, P.; Barker, C.; Hunter, E.; Wain-Hobson, S. Nucleotide sequence of Mason-Pfizer monkey virus: An immunosuppressive D-type retrovirus. Cell 1986, 45, 375–385. [Google Scholar] [CrossRef]
- Sinha, A.; Johnson, W. Retroviruses of the RDR superinfection interference group: Ancient origins and broad host distribution of a promiscuous Env gene. Curr. Opin. Virol. 2017, 25, 105–112. [Google Scholar] [CrossRef]
- Greenwood, A.D.; Ishida, Y.; O’Brien, S.P.; Roca, A.; Eiden, M.V. Transmission, Evolution, and Endogenization: Lessons Learned from Recent Retroviral Invasions. Microbiol. Mol. Biol. Rev. 2018, 82, e00044-17. [Google Scholar] [CrossRef] [Green Version]
- Overbaugh, J.; Miller, A.; Eiden, M. Receptors and entry cofactors for retroviruses include single and multiple transmembrane-spanning proteins as well as newly described glycophosphatidylinositol-anchored and secreted proteins. Microbiol. Mol. Biol. Rev. 2001, 65, 371–389, table of contents. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Zhang, S.; Kronqvist, M.; Wallin, M.; Ekström, M.; Derse, D.; Garoff, H. Intersubunit disulfide isomerization controls membrane fusion of human T-cell leukemia virus Env. J. Virol. 2008, 82, 7135–7143. [Google Scholar] [CrossRef] [Green Version]
- Opstelten, D.J.; Wallin, M.; Garoff, H. Moloney murine leukemia virus envelope protein subunits, gp70 and Pr15E, form a stable disulfide-linked complex. J. Virol. 1998, 72, 6537–6545. [Google Scholar] [CrossRef]
- Pinter, A.; Kopelman, R.; Li, Z.; Kayman, S.C.; A Sanders, D. Localization of the labile disulfide bond between SU and TM of the murine leukemia virus envelope protein complex to a highly conserved CWLC motif in SU that resembles the active-site sequence of thiol-disulfide exchange enzymes. J. Virol. 1997, 71, 8073–8077. [Google Scholar] [CrossRef] [Green Version]
- Tailor, C.S.; Lavillette, D.; Marin, M.; Kabat, D. Cell surface receptors for gammaretroviruses. Curr. Top. Microbiol. Immunol. 2003, 281, 29–106. [Google Scholar]
- Harrison, S.C. Viral membrane fusion. Nat. Struct Mol. Biol 2008, 15, 690–698. [Google Scholar] [CrossRef]
- Gallaher, W.R. Similar structural models of the transmembrane proteins of Ebola and avian sarcoma viruses. Cell 1996, 85, 477–478. [Google Scholar] [CrossRef] [Green Version]
- Volchkov, V.E.; Blinov, V.; Netesov, S. The envelope glycoprotein of Ebola virus contains an immunosuppressive-like domain similar to oncogenic retroviruses. FEBS Lett. 1992, 305, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Benit, L.; Dessen, P.; Heidmann, T. Identification, phylogeny, and evolution of retroviral elements based on their envelope genes. J. Virol. 2001, 75, 11709–11719. [Google Scholar] [CrossRef] [Green Version]
- Stenglein, M.D.; Sanders, C.; Kistler, A.L.; Ruby, J.G.; Franco, J.Y.; Reavill, D.R.; Dunker, F.; Derisi, J.L. Identification, characterization, and in vitro culture of highly divergent arenaviruses from boa constrictors and annulated tree boas: Candidate etiological agents for snake inclusion body disease. mBio 2012, 3, e00180-12. [Google Scholar] [CrossRef] [Green Version]
- Morrison, C.L.; Iwanowicz, L.; Work, T.M.; Fahsbender, E.; Breitbart, M.; Adams, C.; Iwanowicz, D.; Sanders, L.; Ackermann, M.; Cornman, R.S.; et al. Genomic evolution, recombination, and inter-strain diversity of chelonid alphaherpesvirus 5 from Florida and Hawaii green sea turtles with fibropapillomatosis. PeerJ 2018, 6, e4386. [Google Scholar] [CrossRef] [Green Version]
- Malfavon-Borja, R.; Feschotte, C. Fighting fire with fire: Endogenous retrovirus envelopes as restriction factors. J. Virol. 2015, 89, 4047–4050. [Google Scholar] [CrossRef] [Green Version]
- Johnson, W.E. Origins and evolutionary consequences of ancient endogenous retroviruses. Nat. Rev. Microbiol. 2019, 17, 355–370. [Google Scholar] [CrossRef]
- Dupressoir, A.; Lavialle, C.; Heidmann, T. From ancestral infectious retroviruses to bona fide cellular genes: Role of the captured syncytins in placentation. Placenta 2012, 33, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Heidmann, O.; Béguin, A.; Paternina, J.; Berthier, R.; Deloger, M.; Bawa, O.; Heidmann, T. HEMO, an ancestral endogenous retroviral envelope protein shed in the blood of pregnant women and expressed in pluripotent stem cells and tumors. Proc. Natl. Acad. Sci. USA 2017, 114, E6642–E6651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, K.R.; Timpona, J.L.; Jenni, S.; Bloyet, L.-M.; Brusic, V.; Johnson, W.E.; Whelan, S.P.J.; Robinson-McCarthy, L.R. Structure of the Receptor Binding Domain of EnvP(b)1, an Endogenous Retroviral Envelope Protein Expressed in Human Tissues. mBio 2020, 11, e02772-20. [Google Scholar] [CrossRef] [PubMed]
- Henzy, J.E.; Gifford, R.J.; Kenaley, C.P.; Johnson, W.E. An Intact Retroviral Gene Conserved in Spiny-Rayed Fishes for over 100 My. Mol. Biol. Evol. 2017, 34, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.A.; Singh, M.; Cullen, H.B.; Kirou, R.A.; Benkaddour-Boumzaouad, M.; Cortes, J.L.; Pérez, J.G.; Coyne, C.B.; Feschotte, C. Evolution and antiviral activity of a human protein of retroviral origin. Science 2022, 378, 422–428. [Google Scholar] [CrossRef]
- Blaise, S.; de Parseval, N.; Bénit, L.; Heidmann, T. Genomewide screening for fusogenic human endogenous retrovirus envelopes identifies syncytin 2, a gene conserved on primate evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 13013–13018. [Google Scholar] [CrossRef] [Green Version]
- de Parseval, N.; Lazar, V.; Casella, J.F.; Benit, L.; Heidmann, T. Survey of human genes of retroviral origin: Identification and transcriptome of the genes with coding capacity for complete envelope proteins. J. Virol. 2003, 77, 10414–10422. [Google Scholar] [CrossRef] [Green Version]
- Hayward, A.; Grabherr, M.; Jern, P. Broad-scale phylogenomics provides insights into retrovirus-host evolution. Proc. Natl. Acad. Sci. USA 2013, 110, 20146–20151. [Google Scholar] [CrossRef] [Green Version]
- Weiss, R.A. The discovery of endogenous retroviruses. Retrovirology 2006, 3, 67. [Google Scholar] [CrossRef] [Green Version]
- Gifford, R.; Kabat, P.; Martin, J.; Lynch, C.; Tristem, M. Evolution and distribution of class II-related endogenous retroviruses. J. Virol. 2005, 79, 6478–6486. [Google Scholar] [CrossRef] [Green Version]
- Gifford, R.; Tristem, M. The evolution, distribution and diversity of endogenous retroviruses. Virus Genes 2003, 26, 291–315. [Google Scholar] [CrossRef]
- Gifford, R.J.; Blomberg, J.; Coffin, J.M.; Fan, H.; Heidmann, T.; Mayer, J.; Stoye, J.; Tristem, M.; Johnson, W.E. Nomenclature for endogenous retrovirus (ERV) loci. Retrovirology 2018, 15, 59. [Google Scholar] [CrossRef] [Green Version]
- Yedavalli, V.R.K.; Patil, A.; Parrish, J.; Kozak, C.A. A novel class III endogenous retrovirus with a class I envelope gene in African frogs with an intact genome and developmentally regulated transcripts in Xenopus tropicalis. Retrovirology 2021, 18, 20. [Google Scholar] [CrossRef]
- Henzy, J.E.; Gifford, R.J.; Johnson, W.E.; Coffin, J.M. A novel recombinant retrovirus in the genomes of modern birds combines features of avian and mammalian retroviruses. J. Virol. 2014, 88, 2398–2405. [Google Scholar] [CrossRef] [Green Version]
- Farkasova, H.; Hron, T.; Pačes, J.; Hulva, P.; Benda, P.; Gifford, R.J.; Elleder, D. Discovery of an endogenous Deltaretrovirus in the genome of long-fingered bats (Chiroptera: Miniopteridae). Proc. Natl. Acad. Sci. USA 2017, 114, 3145–3150. [Google Scholar] [CrossRef] [Green Version]
- Hron, T.; Farkašová, H.; Gifford, R.J.; Benda, P.; Hulva, P.; Görföl, T.; Pačes, J.; Elleder, D. Remnants of an Ancient Deltaretrovirus in the Genomes of Horseshoe Bats (Rhinolophidae). Viruses 2018, 10, 185. [Google Scholar] [CrossRef] [Green Version]
- Hron, T.; Elleder, D.; Gifford, R. Deltaretroviruses have circulated since at least the Paleogene and infected a broad range of mammalian species. Retrovirology 2019, 16, 33. [Google Scholar] [CrossRef]
- Hayward, A.; Cornwallis, C.; Jern, P. Pan-vertebrate comparative genomics unmasks retrovirus macroevolution. Proc. Natl. Acad. Sci. USA 2015, 112, 464–469. [Google Scholar] [CrossRef] [Green Version]
- Basta, H.A.; Cleveland, S.B.; Clinton, R.A.; Dimitrov, A.G.; McClure, M.A. Evolution of teleost fish retroviruses: Characterization of new retroviruses with cellular genes. J. Virol. 2009, 83, 10152–10162. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Yang, Z.-Z.; Li, H.; Hong, Y.; Fan, D.-D.; Lin, A.-F.; Xiang, L.-X.; Shao, J.-Z. Genome-Wide Characterization of Zebrafish Endogenous Retroviruses Reveals Unexpected Diversity in Genetic Organizations and Functional Potentials. Microbiol. Spectr. 2021, 9, e0225421. [Google Scholar] [CrossRef]
- Shen, C.H.; Steiner, L. Genome structure and thymic expression of an endogenous retrovirus in zebrafish. J. Virol. 2004, 78, 899–911. [Google Scholar] [CrossRef]
- Frank, J.A.; Feschotte, C. Co-option of endogenous viral sequences for host cell function. Curr. Opin. Virol. 2017, 25, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Lavialle, C.; Cornelis, G.; Dupressoir, A.; Esnault, C.; Heidmann, O.; Vernochet, C.; Heidmann, T. Paleovirology of ‘syncytins’, retroviral env genes exapted for a role in placentation. Philos Trans. R Soc. Lond B Biol. Sci. 2013, 368, 20120507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Best, S.; Le Tissier, P.; Stoye, J. Endogenous retroviruses and the evolution of resistance to retroviral infection. Trends Microbiol. 1997, 5, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.; Kozak, C.; Boso, G. Cross-species transmission of an ancient endogenous retrovirus and convergent co-option of its envelope gene in two mammalian orders. PLoS Genet. 2022, 18, e1010458. [Google Scholar] [CrossRef]
- Diehl, W.E.; Patel, N.; Halm, K.; Johnson, W.E. Tracking interspecies transmission and long-term evolution of an ancient retrovirus using the genomes of modern mammals. Elife 2016, 5, e12704. [Google Scholar] [CrossRef]
- Ueda, M.T.; Kryukov, K.; Mitsuhashi, S.; Mitsuhashi, H.; Imanishi, T.; Nakagawa, S. Comprehensive genomic analysis reveals dynamic evolution of endogenous retroviruses that code for retroviral-like protein domains. Mob. DNA 2020, 11, 29. [Google Scholar] [CrossRef]
- Kasperek, A.; Béguin, A.; Bawa, O.; De Azevedo, K.; Job, B.; Massard, C.; Scoazec, J.; Heidmann, T.; Heidmann, O. Therapeutic potential of the human endogenous retroviral envelope protein HEMO: A pan-cancer analysis. Mol. Oncol. 2022, 16, 1451–1473. [Google Scholar] [CrossRef]
- Mi, S.; Lee, X.; Li, X.P.; Veldman, G.M.; Finnerty, H.; Racie, L.; LaVallie, E.; Tang, X.Y.; Edouard, P.; Howes, S.; et al. Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis. Nature 2000, 403, 785–789. [Google Scholar] [CrossRef]
- Blond, J.L.; Lavillette, D.; Cheynet, V.; Bouton, O.; Oriol, G.; Chapel-Fernandes, S.; Mandrand, B.; Mallet, F.; Cosset, F.-L. An envelope glycoprotein of the human endogenous retrovirus HERV-W is expressed in the human placenta and fuses cells expressing the type D mammalian retrovirus receptor. J. Virol. 2000, 74, 3321–3329. [Google Scholar] [CrossRef] [Green Version]
- Cornelis, G.; Heidmann, O.; Bernard-Stoecklin, S.; Reynaud, K.; Véron, G.; Mulot, B.; Dupressoir, A.; Heidmann, T. Ancestral capture of syncytin-Car1, a fusogenic endogenous retroviral envelope gene involved in placentation and conserved in Carnivora. Proc. Natl. Acad. Sci. USA 2012, 109, E432–E441. [Google Scholar] [CrossRef]
- Imakawa, K.; Nakagawa, S. The Phylogeny of Placental Evolution Through Dynamic Integrations of Retrotransposons. Prog. Mol. Biol. Transl. Sci. 2017, 145, 89–109. [Google Scholar]
- Cornelis, G.; Funk, M.; Vernochet, C.; Leal, F.; Tarazona, O.A.; Meurice, G.; Heidmann, O.; Dupressoir, A.; Miralles, A.; Ramirez-Pinilla, M.P.; et al. An endogenous retroviral envelope syncytin and its cognate receptor identified in the viviparous placental Mabuya lizard. Proc. Natl. Acad. Sci. USA 2017, 114, E10991–E11000. [Google Scholar] [CrossRef] [Green Version]
- Funk, M.; Cornelis, G.; Vernochet, C.; Heidmann, O.; Dupressoir, A.; Conley, A.; Glickman, S.; Heidmann, T. Capture of a Hyena-Specific Retroviral Envelope Gene with Placental Expression Associated in Evolution with the Unique Emergence among Carnivorans of Hemochorial Placentation in Hyaenidae. J. Virol. 2019, 93, e01811-18. [Google Scholar] [CrossRef] [Green Version]
- Gardner, M.B.; Kozak, C.; O’Brien, S.J. The Lake Casitas wild mouse: Evolving genetic resistance to retroviral disease. Trends Genet. 1991, 7, 22–27. [Google Scholar] [CrossRef]
- Ikeda, H.; Sugimura, H. Fv-4 resistance gene: A truncated endogenous murine leukemia virus with ecotropic interference properties. J. Virol. 1989, 63, 5405–5412. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S. FV-4: A new gene affecting the splenomegaly induction by Friend leukemia virus. Jpn J. Exp. Med. 1975, 45, 473–478. [Google Scholar]
- Inaguma, Y.; Miyashita, N.; Moriwaki, K.; Huai, W.C.; Jin, M.L.; He, X.Q.; Ikeda, H. Acquisition of two endogenous ecotropic murine leukemia viruses in distinct Asian wild mouse populations. J. Virol. 1991, 65, 1796–1802. [Google Scholar] [CrossRef] [Green Version]
- Limjoco, T.I.; Dickie, P.; Ikeda, H.; Silver, J. Transgenic Fv-4 mice resistant to Friend virus. J. Virol. 1993, 67, 4163–4168. [Google Scholar] [CrossRef] [Green Version]
- Gardner, M.B.; Rasheed, S.; Pal, B.K.; Estes, J.D.; O’Brien, S.J. Akvr-1, a dominant murine leukemia virus restriction gene, is polymorphic in leukemia-prone wild mice. Proc. Natl. Acad. Sci. USA 1980, 77, 531–535. [Google Scholar] [CrossRef] [Green Version]
- Kozak, C.A.; Gromet, N.J.; Ikeda, H.; E Buckler, C. A unique sequence related to the ecotropic murine leukemia virus is associated with the Fv-4 resistance gene. Proc. Natl. Acad. Sci. USA 1984, 81, 834–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dandekar, S.; Rossitto, P.; Pickett, S.; Mockli, G.; Bradshaw, H.; Cardiff, R.; Gardner, M. Molecular characterization of the Akvr-1 restriction gene: A defective endogenous retrovirus-borne gene identical to Fv-4r. J. Virol. 1987, 61, 308–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buller, R.S.; Ahmed, A.; Portis, J. Identification of two forms of an endogenous murine retroviral env gene linked to the Rmcf locus. J. Virol. 1987, 61, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Buller, R.S.; Sitbon, M.; Portis, J. The endogenous mink cell focus-forming (MCF) gp70 linked to the Rmcf gene restricts MCF virus replication in vivo and provides partial resistance to erythroleukemia induced by Friend murine leukemia virus. J. Exp. Med. 1988, 167, 1535–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.T.; Lyu, M.S.; Buckler-White, A.; Kozak, C.A. Characterization of a polytropic murine leukemia virus proviral sequence associated with the virus resistance gene Rmcf of DBA/2 mice. J. Virol. 2002, 76, 8218–8224. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Yan, Y.; Kozak, C. Rmcf2, a xenotropic provirus in the Asian mouse species Mus castaneus, blocks infection by polytropic mouse gammaretroviruses. J. Virol. 2005, 79, 9677–9684. [Google Scholar] [CrossRef] [Green Version]
- Takeda, A.; Matano, T. Inhibition of infectious murine leukemia virus production by Fv-4 env gene products exerting dominant negative effect on viral envelope glycoprotein. Microbes Infect. 2007, 9, 1590–1596. [Google Scholar] [CrossRef]
- Dewannieux, M.; Collins, M.K. Spontaneous heteromerization of gammaretrovirus envelope proteins: A possible novel mechanism of retrovirus restriction. J. Virol. 2008, 82, 9789–9794. [Google Scholar] [CrossRef] [Green Version]
- Taylor, G.M.; Gao, Y.; Sanders, D. Fv-4: Identification of the defect in Env and the mechanism of resistance to ecotropic murine leukemia virus. J. Virol. 2001, 75, 11244–11248. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, S.; Takahashi, M.U. gEVE: A genome-based endogenous viral element database provides comprehensive viral protein-coding sequences in mammalian genomes. Database (Oxford) 2016, 2016, baw087. [Google Scholar] [CrossRef] [Green Version]
- Ponferrada, V.G.; Mauck, B.; Wooley, D. The envelope glycoprotein of human endogenous retrovirus HERV-W induces cellular resistance to spleen necrosis virus. Arch. Virol. 2003, 148, 659–675. [Google Scholar] [CrossRef]
- Blanco-Melo, D.; Gifford, R.; Bieniasz, P. Co-option of an endogenous retrovirus envelope for host defense in hominid ancestors. Elife 2017, 6, e22519. [Google Scholar] [CrossRef]
- Murakami, T. Retroviral env glycoprotein trafficking and incorporation into virions. Mol. Biol. Int. 2012, 2012, 682850. [Google Scholar] [CrossRef]
- Tedbury, P.R.; Freed, E.O. The cytoplasmic tail of retroviral envelope glycoproteins. Prog. Mol. Biol. Transl. Sci. 2015, 129, 253–284. [Google Scholar]
- Sandrin, V.; Muriaux, D.; Darlix, J.-L.; Cosset, F.-L. Intracellular trafficking of Gag and Env proteins and their interactions modulate pseudotyping of retroviruses. J. Virol. 2004, 78, 7153–7164. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.Y.; Dubay, J.W.; Perez, L.G.; Hunter, E. Mutations within the proteolytic cleavage site of the Rous sarcoma virus glycoprotein define a requirement for dibasic residues for intracellular cleavage. J. Virol. 1992, 66, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Sjoberg, M.; Wu, S.-R.; Löving, R.; Rantalainen, K.; Lindqvist, B.; Garoff, H.; Shaltiel, I.A.; Aprelia, M.; Saurin, A.T.; Chowdhury, D.; et al. Furin cleavage of the Moloney murine leukemia virus Env precursor reorganizes the spike structure. Proc. Natl. Acad. Sci. USA 2014, 111, 6034–6039. [Google Scholar] [CrossRef] [Green Version]
- Zavorotinskaya, T.; Albritton, L.M. Failure To cleave murine leukemia virus envelope protein does not preclude its incorporation in virions and productive virus-receptor interaction. J. Virol. 1999, 73, 5621–5629. [Google Scholar] [CrossRef] [Green Version]
- Freed, E.O.; Risser, R. The role of envelope glycoprotein processing in murine leukemia virus infection. J. Virol. 1987, 61, 2852–2856. [Google Scholar] [CrossRef] [Green Version]
- Owji, H.; Nezafat, N.; Negahdaripour, M.; Hajiebrahimi, A.; Ghasemi, Y. A comprehensive review of signal peptides: Structure, roles, and applications. Eur. J. Cell Biol. 2018, 97, 422–441. [Google Scholar] [CrossRef]
- Hoch-Marchaim, H.; Weiss, A.M.; Bar-Sinai, A.; Fromer, M.; Adermann, K.; Hochman, J. The leader peptide of MMTV Env precursor localizes to the nucleoli in MMTV-derived T cell lymphomas and interacts with nucleolar protein B23. Virology 2003, 313, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Indik, S.; Günzburg, W.H.; Salmons, B.; Rouault, F. A novel, mouse mammary tumor virus encoded protein with Rev-like properties. Virology 2005, 337, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mertz, J.A.; Simper, M.S.; Lozano, M.M.; Payne, S.M.; Dudley, J.P. Mouse mammary tumor virus encodes a self-regulatory RNA export protein and is a complex retrovirus. J. Virol. 2005, 79, 14737–14747. [Google Scholar] [CrossRef] [Green Version]
- Caporale, M.; Arnaud, F.; Mura, M.; Golder, M.; Murgia, C.; Palmarini, M. The signal peptide of a simple retrovirus envelope functions as a posttranscriptional regulator of viral gene expression. J. Virol. 2009, 83, 4591–4604. [Google Scholar] [CrossRef] [PubMed]
- Hofacre, A.; Nitta, T.; Fan, H. Jaagsiekte sheep retrovirus encodes a regulatory factor, Rej, required for synthesis of Gag protein. J. Virol. 2009, 83, 12483–12498. [Google Scholar] [CrossRef] [Green Version]
- Byun, H.; Halani, N.; Gou, Y.; Nash, A.K.; Lozano, M.M.; Dudley, J.P. Requirements for mouse mammary tumor virus Rem signal peptide processing and function. J. Virol. 2012, 86, 214–225. [Google Scholar] [CrossRef] [Green Version]
- McCaul, N.; Quandte, M.; Bontjer, I.; van Zadelhoff, G.; Land, A.; Crooks, E.T.; Binley, J.M.; Sanders, R.W.; Braakman, I. Intramolecular quality control: HIV-1 envelope gp160 signal-peptide cleavage as a functional folding checkpoint. Cell Rep. 2021, 36, 109646. [Google Scholar] [CrossRef]
- Liu, H.; Wu, R.; Yuan, L.; Tian, G.; Huang, X.; Wen, Y.; Ma, X.; Huang, Y.; Yan, Q.; Zhao, Q.; et al. Introducing a cleavable signal peptide enhances the packaging efficiency of lentiviral vectors pseudotyped with Japanese encephalitis virus envelope proteins. Virus Res. 2017, 229, 9–16. [Google Scholar] [CrossRef]
- Yolitz, J.; Schwing, C.; Chang, J.; Van Ryk, D.; Nawaz, F.; Wei, D.; Cicala, C.; Arthos, J.; Fauci, A.S. Signal peptide of HIV envelope protein impacts glycosylation and antigenicity of gp120. Proc. Natl. Acad. Sci. USA 2018, 115, 2443–2448. [Google Scholar] [CrossRef] [Green Version]
- Marzi, A.; Akhavan, A.; Simmons, G.; Gramberg, T.; Hofmann, H.; Bates, P.; Lingappa, V.R.; Pöhlmann, S. The signal peptide of the ebolavirus glycoprotein influences interaction with the cellular lectins DC-SIGN and DC-SIGNR. J. Virol. 2006, 80, 6305–6317. [Google Scholar] [CrossRef] [Green Version]
- Kim, F.J.; Manel, N.; Garrido, E.N.; Valle, C.; Sitbon, M.; Battini, J.-L. HTLV-1 and -2 envelope SU subdomains and critical determinants in receptor binding. Retrovirology 2004, 1, 41. [Google Scholar] [CrossRef] [Green Version]
- Davey, R.A.; A Hamson, C.; Healey, J.J.; Cunningham, J.M. In vitro binding of purified murine ecotropic retrovirus envelope surface protein to its receptor, MCAT-1. J. Virol. 1997, 71, 8096–8102. [Google Scholar] [CrossRef] [Green Version]
- Davey, R.A.; Zuo, Y.; Cunningham, J. Identification of a receptor-binding pocket on the envelope protein of friend murine leukemia virus. J. Virol. 1999, 73, 3758–3763. [Google Scholar] [CrossRef] [Green Version]
- Bae, Y.; Kingsman, S.; Kingsman, A. Functional dissection of the Moloney murine leukemia virus envelope protein gp70. J. Virol. 1997, 71, 2092–2099. [Google Scholar] [CrossRef]
- Battini, J.L.; Danos, O.; Heard, J. Receptor-binding domain of murine leukemia virus envelope glycoproteins. J. Virol. 1995, 69, 713–719. [Google Scholar] [CrossRef] [Green Version]
- Martinez, I.; Dornburg, R. Mutational analysis of the envelope protein of spleen necrosis virus. J. Virol. 1996, 70, 6036–6043. [Google Scholar] [CrossRef] [Green Version]
- Ott, D.; Friedrich, R.; Rein, A. Sequence analysis of amphotropic and 10A1 murine leukemia viruses: Close relationship to mink cell focus-inducing viruses. J. Virol. 1990, 64, 757–766. [Google Scholar] [CrossRef] [Green Version]
- Ott, D.; Rein, A. Basis for receptor specificity of nonecotropic murine leukemia virus surface glycoprotein gp70SU. J. Virol. 1992, 66, 4632–4638. [Google Scholar] [CrossRef] [Green Version]
- Battini, J.L.; Heard, J.; Danos, O. Receptor choice determinants in the envelope glycoproteins of amphotropic, xenotropic, and polytropic murine leukemia viruses. J. Virol. 1992, 66, 1468–1475. [Google Scholar] [CrossRef] [Green Version]
- Federspiel, M.J. Reverse Engineering Provides Insights on the Evolution of Subgroups A to E Avian Sarcoma and Leukosis Virus Receptor Specificity. Viruses 2019, 11, 497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorner, A.J.; Stoye, J.; Coffin, J. Molecular basis of host range variation in avian retroviruses. J. Virol. 1985, 53, 32–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kan, N.C.; Baluda, M.; Papas, T. Sites of recombination between the transforming gene of avian myeloblastosis virus and its helper virus. Virology 1985, 145, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Bova, C.A.; Manfredi, J.; Swanstrom, R. Env genes of avian retroviruses: Nucleotide sequence and molecular recombinants define host range determinants. Virology 1986, 152, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Dorner, A.J.; Coffin, J.M. Determinants for receptor interaction and cell killing on the avian retrovirus glycoprotein gp85. Cell 1986, 45, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Bova, C.A.; Olsen, J.; Swanstrom, R. The avian retrovirus env gene family: Molecular analysis of host range and antigenic variants. J. Virol. 1988, 62, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Johnston, E.R.; Albritton, L.; Radke, K. Envelope proteins containing single amino acid substitutions support a structural model of the receptor-binding domain of bovine leukemia virus surface protein. J. Virol. 2002, 76, 10861–10872. [Google Scholar] [CrossRef] [Green Version]
- Kim, F.J.; Seiliez, I.; Denesvre, C.; Lavillette, D.; Cosset, F.-L.; Sitbon, M. Definition of an amino-terminal domain of the human T-cell leukemia virus type 1 envelope surface unit that extends the fusogenic range of an ecotropic murine leukemia virus. J. Biol. Chem. 2000, 275, 23417–23420. [Google Scholar] [CrossRef] [Green Version]
- Barnett, A.L.; Wensel, D.L.; Li, W.; Fass, D.; Cunningham, J.M. Structure and mechanism of a coreceptor for infection by a pathogenic feline retrovirus. J. Virol. 2003, 77, 2717–2729. [Google Scholar] [CrossRef] [Green Version]
- Fass, D.; Davey, R.A.; Hamson, C.A.; Kim, P.S.; Cunningham, J.M.; Berger, J.M. Structure of a murine leukemia virus receptor-binding glycoprotein at 2.0 angstrom resolution. Science 1997, 277, 1662–1666. [Google Scholar] [CrossRef]
- Koch, W.; Hunsmann, G.; Friedrich, R. Nucleotide sequence of the envelope gene of Friend murine leukemia virus. J. Virol. 1983, 45, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Burkhart, M.D.; Kayman, S.C.; He, Y.; Pinter, A. Distinct mechanisms of neutralization by monoclonal antibodies specific for sites in the N-terminal or C-terminal domain of murine leukemia virus SU. J. Virol. 2003, 77, 3993–4003. [Google Scholar] [CrossRef] [Green Version]
- Fontenot, J.D.; Tjandra, N.; Ho, C.; Andrews, P.C.; Montelaro, R.C. Structure and self assembly of a retrovirus (FeLV) proline rich neutralization domain. J. Biomol. Struct Dyn. 1994, 11, 821–836. [Google Scholar] [CrossRef]
- Lavillette, D.; Maurice, M.; Roche, C.; Russell, S.J.; Sitbon, M.; Cosset, F.-L. A proline-rich motif downstream of the receptor binding domain modulates conformation and fusogenicity of murine retroviral envelopes. J. Virol. 1998, 72, 9955–9965. [Google Scholar] [CrossRef] [Green Version]
- Machida, C.A.; Bestwick, R.K.; Boswell, B.A.; Kabat, D. Role of a membrane glycoprotein in Friend virus-induced erythroleukemia: Studies of mutant and revertant viruses. Virology 1985, 144, 158–172. [Google Scholar] [CrossRef]
- Wu, B.W.; Lu, J.; Gallaher, T.K.; Anderson, W.; Cannon, P.M. Identification of regions in the Moloney murine leukemia virus SU protein that tolerate the insertion of an integrin-binding peptide. Virology 2000, 269, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Kayman, S.C.; Park, H.; Saxon, M.; Pinter, A. The hypervariable domain of the murine leukemia virus surface protein tolerates large insertions and deletions, enabling development of a retroviral particle display system. J. Virol. 1999, 73, 1802–1808. [Google Scholar] [CrossRef] [Green Version]
- Riedel, C.; Vasishtan, D.; Siebert, C.A.; Whittle, C.; Lehmann, M.J.; Mothes, W.; Grünewald, K. Native structure of a retroviral envelope protein and its conformational change upon interaction with the target cell. J. Struct Biol. 2017, 197, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Wang, H.; Lu, X.; Wang, R.; Zheng, R.; Li, Y.; Yang, X.; Jia, W.-T.; Zhao, Y.; Wang, Y.; et al. Effects of individually silenced N-glycosylation sites and non-synonymous single-nucleotide polymorphisms on the fusogenic function of human syncytin-2. Cell Adh. Migr. 2016, 10, 39–55. [Google Scholar] [CrossRef]
- Henzy, J.E.; Coffin, J.M. Betaretroviral envelope subunits are noncovalently associated and restricted to the mammalian class. J. Virol. 2013, 87, 1937–1946. [Google Scholar] [CrossRef] [Green Version]
- Wallin, M.; Ekstrom, M.; Garoff, H. Isomerization of the intersubunit disulphide-bond in Env controls retrovirus fusion. EMBO J. 2004, 23, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Wallin, M.; Ekstrom, M.; Garoff, H. Receptor-triggered but alkylation-arrested env of murine leukemia virus reveals the transmembrane subunit in a prehairpin conformation. J. Virol. 2006, 80, 9921–9925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitbon, M.; D’Auriol, L.; Ellerbrok, H.; André, C.; Nishio, J.; Perryman, S.; Pozo, F.; Hayes, S.F.; Wehrly, K.; Tambourin, P. Substitution of leucine for isoleucine in a sequence highly conserved among retroviral envelope surface glycoproteins attenuates the lytic effect of the Friend murine leukemia virus. Proc. Natl. Acad. Sci. USA 1991, 88, 5932–5936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.G.; Cunningham, J.M. Receptor-induced thiolate couples Env activation to retrovirus fusion and infection. PLoS Pathog 2007, 3, e198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mothes, W.; Boerger, A.L.; Narayan, S.; Cunningham, J.M.; Young, J.A. Retroviral entry mediated by receptor priming and low pH triggering of an envelope glycoprotein. Cell 2000, 103, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.G.; Mothes, W.; Blacklow, S.C.; Cunningham, J.M. The mature avian leukosis virus subgroup A envelope glycoprotein is metastable, and refolding induced by the synergistic effects of receptor binding and low pH is coupled to infection. J. Virol. 2004, 78, 1403–1410. [Google Scholar] [CrossRef] [Green Version]
- Harrison, S.C. Viral membrane fusion. Virology 2015, 479–480, 498–507. [Google Scholar] [CrossRef] [Green Version]
- Lozada, C.; Barlow, T.M.A.; Gonzalez, S.; Lubin-Germain, N.; Ballet, S. Identification and Characteristics of Fusion Peptides Derived From Enveloped Viruses. Front. Chem. 2021, 9, 689006. [Google Scholar] [CrossRef]
- Zhu, N.L.; Cannon, P.M.; Chen, D.; Anderson, W.F. Mutational analysis of the fusion peptide of Moloney murine leukemia virus transmembrane protein p15E. J. Virol. 1998, 72, 1632–1639. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, L.D.; White, J.M. Mutational analysis of the candidate internal fusion peptide of the avian leukosis and sarcoma virus subgroup A envelope glycoprotein. J. Virol. 1998, 72, 3259–3267. [Google Scholar] [CrossRef] [Green Version]
- Davies, S.M.; Kelly, S.M.; Price, N.C.; Bradshaw, J.P. Structural plasticity of the feline leukaemia virus fusion peptide: A circular dichroism study. FEBS Lett. 1998, 425, 415–418. [Google Scholar] [CrossRef] [Green Version]
- Delos, S.E.; Gilbert, J.; White, J. The central proline of an internal viral fusion peptide serves two important roles. J. Virol. 2000, 74, 1686–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, P.; Pringle, C.; Easton, A. Heptad repeat sequences are located adjacent to hydrophobic regions in several types of virus fusion glycoproteins. J. Gen. Virol. 1990, 71 (Pt 12), 3075–3080. [Google Scholar] [CrossRef]
- Netter, R.C.; Amberg, S.M.; Balliet, J.W.; Biscone, M.J.; Vermeulen, A.; Earp, L.J.; White, J.M.; Bates, P. Heptad repeat 2-based peptides inhibit avian sarcoma and leukosis virus subgroup a infection and identify a fusion intermediate. J. Virol. 2004, 78, 13430–13439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fass, D.; Harrison, S.; Kim, P. Retrovirus envelope domain at 1.7 angstrom resolution. Nat. Struct. Biol. 1996, 3, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Ou, W.; Silver, J. Inhibition of murine leukemia virus envelope protein (env) processing by intracellular expression of the env N-terminal heptad repeat region. J. Virol. 2005, 79, 4782–4792. [Google Scholar] [CrossRef] [Green Version]
- Harrison, J.S.; Koellhoffer, J.F.; Chandran, K.; Lai, J.R. Marburg virus glycoprotein GP2: pH-dependent stability of the ectodomain alpha-helical bundle. Biochemistry 2012, 51, 2515–2525. [Google Scholar] [CrossRef] [Green Version]
- Hetzel, U.; Sironen, T.; Laurinmäki, P.; Liljeroos, L.; Patjas, A.; Henttonen, H.; Vaheri, A.; Artelt, A.; Kipar, A.; Butcher, S.; et al. Isolation, identification, and characterization of novel arenaviruses, the etiological agents of boid inclusion body disease. J. Virol. 2013, 87, 10918–10935. [Google Scholar] [CrossRef] [Green Version]
- Koellhoffer, J.F.; Malashkevich, V.N.; Harrison, J.S.; Toro, R.; Bhosle, R.C.; Chandran, K.; Almo, S.C.; Lai, J.R. Crystal structure of the Marburg virus GP2 core domain in its postfusion conformation. Biochemistry 2012, 51, 7665–7675. [Google Scholar] [CrossRef] [Green Version]
- Cianciolo, G.J.; Bogerd, H.P.; Kipnis, R.J.; Copeland, T.D.; Oroszlan, S.; Snyderman, R. Inhibition of lymphocyte proliferation by a synthetic peptide homologous to envelope proteins of human and animal retroviruses. Trans. Assoc. Am. Physicians 1985, 98, 30–41. [Google Scholar]
- Cianciolo, G.J.; Copeland, T.D.; Oroszlan, S.; Snyderman, R. Inhibition of lymphocyte proliferation by a synthetic peptide homologous to retroviral envelope proteins. Science 1985, 230, 453–455. [Google Scholar] [CrossRef]
- Kadota, J.; Cianciolo, G.; Snyderman, R. A synthetic peptide homologous to retroviral transmembrane envelope proteins depresses protein kinase C mediated lymphocyte proliferation and directly inactivated protein kinase C: A potential mechanism for immunosuppression. Microbiol. Immunol. 1991, 35, 443–459. [Google Scholar] [CrossRef]
- Kleinerman, E.S.; Lachman, L.B.; Knowles, R.D.; Snyderman, R.; Cianciolo, G.J. A synthetic peptide homologous to the envelope proteins of retroviruses inhibits monocyte-mediated killing by inactivating interleukin 1. J. Immunol. 1987, 139, 2329–2337. [Google Scholar] [CrossRef]
- Blaise, S.; Mangeney, M.; Heidmann, T. The envelope of Mason-Pfizer monkey virus has immunosuppressive properties. J. Gen. Virol. 2001, 82 (Pt 7), 1597–1600. [Google Scholar] [CrossRef] [Green Version]
- Mangeney, M.; Heidmann, T. Tumor cells expressing a retroviral envelope escape immune rejection in vivo. Proc. Natl. Acad. Sci. USA 1998, 95, 14920–14925. [Google Scholar] [CrossRef] [Green Version]
- Mangeney, M.; Renard, M.; Schlecht-Louf, G.; Bouallaga, I.; Heidmann, O.; Letzelter, C.; Richaud, A.; Ducos, B.; Heidmann, T. Placental syncytins: Genetic disjunction between the fusogenic and immunosuppressive activity of retroviral envelope proteins. Proc. Natl. Acad. Sci. USA 2007, 104, 20534–20539. [Google Scholar] [CrossRef]
- Schlecht-Louf, G.; Renard, M.; Mangeney, M.; Letzelter, C.; Richaud, A.; Ducos, B.; Bouallaga, I.; Heidmann, T. Retroviral infection in vivo requires an immune escape virulence factor encrypted in the envelope protein of oncoretroviruses. Proc. Natl. Acad. Sci. USA 2010, 107, 3782–3787. [Google Scholar] [CrossRef] [Green Version]
- Eksmond, U.; Jenkins, B.; Merkenschlager, J.; Mothes, W.; Stoye, J.P.; Kassiotis, G. Mutation of the Putative Immunosuppressive Domain of the Retroviral Envelope Glycoprotein Compromises Infectivity. J. Virol. 2017, 91, e01152-17. [Google Scholar] [CrossRef] [Green Version]
- Brody, B.A.; Hunter, E. Mutations within the env gene of Mason-Pfizer monkey virus: Effects on protein transport and SU-TM association. J. Virol. 1992, 66, 3466–3475. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Micoli, K.; Hunter, E. Activity of the Mason-Pfizer monkey virus fusion protein is modulated by single amino acids in the cytoplasmic tail. J. Virol. 2005, 79, 11569–11579. [Google Scholar] [CrossRef] [Green Version]
- Blot, V.; Lopez-Vergès, S.; Breton, M.; Pique, C.; Berlioz-Torrent, C.; Grange, M.-P. The conserved dileucine- and tyrosine-based motifs in MLV and MPMV envelope glycoproteins are both important to regulate a common Env intracellular trafficking. Retrovirology 2006, 3, 62. [Google Scholar] [CrossRef] [Green Version]
- Bouard, D.; Sandrin, V.; Boson, B.; Nègre, D.; Thomas, G.; Granier, C.; Cosset, F.-L. An acidic cluster of the cytoplasmic tail of the RD114 virus glycoprotein controls assembly of retroviral envelopes. Traffic 2007, 8, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Micoli, K.; Bauerova, H.; Pichova, I.; Hunter, E. Amino acid residues in the cytoplasmic domain of the Mason-Pfizer monkey virus glycoprotein critical for its incorporation into virions. J. Virol. 2005, 79, 11559–11568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.C. Mechanisms for Env glycoprotein acquisition by retroviruses. AIDS Res. Hum. Retrovir. 2011, 27, 239–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgenson, R.L.; Vogt, V.; Johnson, M. Foreign glycoproteins can be actively recruited to virus assembly sites during pseudotyping. J. Virol. 2009, 83, 4060–4067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rein, A.; Mirro, J.; Haynes, J.G.; Ernst, S.M.; Nagashima, K. Function of the cytoplasmic domain of a retroviral transmembrane protein: p15E-p2E cleavage activates the membrane fusion capability of the murine leukemia virus Env protein. J. Virol. 1994, 68, 1773–1781. [Google Scholar] [CrossRef]
- Green, N.; Shinnick, T.M.; Witte, O.; Ponticelli, A.; Sutcliffe, J.G.; A Lerner, R. Sequence-specific antibodies show that maturation of Moloney leukemia virus envelope polyprotein involves removal of a COOH-terminal peptide. Proc. Natl. Acad. Sci. USA 1981, 78, 6023–6027. [Google Scholar] [CrossRef] [Green Version]
- Sutcliffe, J.G.; Shinnick, T.M.; Green, N.; Liu, F.-T.; Niman, H.L.; Lerner, R.A. Chemical synthesis of a polypeptide predicted from nucleotide sequence allows detection of a new retroviral gene product. Nature 1980, 287, 801–805. [Google Scholar] [CrossRef]
- Bobkova, M.; Stitz, J.; Engelstädter, M.; Cichutek, K.; Buchholz, C.J. Identification of R-peptides in envelope proteins of C-type retroviruses. J. Gen. Virol. 2002, 83 (Pt 9), 2241–2246. [Google Scholar] [CrossRef]
- Ragheb, J.A.; Anderson, W.F. pH-independent murine leukemia virus ecotropic envelope-mediated cell fusion: Implications for the role of the R peptide and p12E TM in viral entry. J. Virol. 1994, 68, 3220–3231. [Google Scholar] [CrossRef] [Green Version]
- Kim, F.J.; Manel, N.; Boublik, Y.; Battini, J.-L.; Sitbon, M. Human T-cell leukemia virus type 1 envelope-mediated syncytium formation can be activated in resistant Mammalian cell lines by a carboxy-terminal truncation of the envelope cytoplasmic domain. J. Virol. 2003, 77, 963–969. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Li, Z.-N.; Yao, Q.; Yang, C.; Steinhauer, D.A.; Compans, R.W. Murine leukemia virus R Peptide inhibits influenza virus hemagglutinin-induced membrane fusion. J. Virol. 2006, 80, 6106–6114. [Google Scholar] [CrossRef] [Green Version]
- Lucas, T.M.; Lyddon, T.D.; Grosse, S.A.; Johnson, M.C. Two distinct mechanisms regulate recruitment of murine leukemia virus envelope protein to retroviral assembly sites. Virology 2010, 405, 548–555. [Google Scholar] [CrossRef]
- Rein, A. Across the Hall from Pioneers. Viruses 2021, 13, 491. [Google Scholar] [CrossRef]
- Henderson, L.E.; Sowder, R.; Copeland, T.D.; Smythers, G.; Oroszlan, S. Quantitative separation of murine leukemia virus proteins by reversed-phase high-pressure liquid chromatography reveals newly described gag and env cleavage products. J. Virol. 1984, 52, 492–500. [Google Scholar] [CrossRef] [Green Version]
- Kubo, Y.; Amanuma, H. Mutational analysis of the R peptide cleavage site of Moloney murine leukaemia virus envelope protein. J. Gen. Virol. 2003, 84 (Pt 8), 2253–2257. [Google Scholar] [CrossRef]
- Loving, R.; Li, K.; Wallin, M.; Sjöberg, M.; Garoff, H. R-Peptide cleavage potentiates fusion-controlling isomerization of the intersubunit disulfide in Moloney murine leukemia virus Env. J. Virol. 2008, 82, 2594–2597. [Google Scholar] [CrossRef] [Green Version]
- Loving, R.; Wu, S.-R.; Sjöberg, M.; Lindqvist, B.; Garoff, H. Maturation cleavage of the murine leukemia virus Env precursor separates the transmembrane subunits to prime it for receptor triggering. Proc. Natl. Acad. Sci. USA 2012, 109, 7735–7740. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Yang, C.; Compans, R. Mutations in the cytoplasmic tail of murine leukemia virus envelope protein suppress fusion inhibition by R peptide. J. Virol. 2001, 75, 2337–2344. [Google Scholar] [CrossRef] [Green Version]
- Kubo, Y.; Izumida, M.; Togawa, K.; Zhang, F.; Hayashi, H. Cytoplasmic R-peptide of murine leukemia virus envelope protein negatively regulates its interaction with the cell surface receptor. Virology 2019, 532, 82–87. [Google Scholar] [CrossRef]
- Schneider, I.C.; Eckhardt, M.; Brynza, J.; Collins, M.K.; Cichutek, K.; Buchholz, C.J. Escape from R-peptide deletion in a gamma-retrovirus. Virology 2011, 418, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Cardone, G.; Brecher, M.; Fontana, J.; Winkler, D.C.; Butan, C.; White, J.M.; Steven, A.C. Visualization of the two-step fusion process of the retrovirus avian sarcoma/leukosis virus by cryo-electron tomography. J. Virol. 2012, 86, 12129–12137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melikyan, G.B.; Barnard, R.J.O.; Markosyan, R.M.; Young, J.A.T.; Cohen, F.S. Low pH is required for avian sarcoma and leukosis virus Env-induced hemifusion and fusion pore formation but not for pore growth. J. Virol. 2004, 78, 3753–3762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nussbaum, O.; Roop, A.; Anderson, W. Sequences determining the pH dependence of viral entry are distinct from the host range-determining region of the murine ecotropic and amphotropic retrovirus envelope proteins. J. Virol. 1993, 67, 7402–7405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, I.; Vilhardt, F. Macropinocytosis is the entry mechanism of amphotropic murine leukemia virus. J. Virol. 2015, 89, 1851–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavillette, D.; Ruggieri, A.; Russell, S.J.; Cosset, F.-L. Activation of a cell entry pathway common to type C mammalian retroviruses by soluble envelope fragments. J. Virol. 2000, 74, 295–304. [Google Scholar] [CrossRef]
- Barnett, A.L.; Davey, R.; Cunningham, J. Modular organization of the Friend murine leukemia virus envelope protein underlies the mechanism of infection. Proc. Natl. Acad. Sci. USA 2001, 98, 4113–4118. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, L.; Roth, M.J. Second-site changes affect viability of amphotropic/ecotropic chimeric enveloped murine leukemia viruses. J. Virol. 2000, 74, 899–913. [Google Scholar] [CrossRef] [Green Version]
- Farrell, K.B.; Ting, Y.; Eiden, M. Fusion-defective gibbon ape leukemia virus vectors can be rescued by homologous but not heterologous soluble envelope proteins. J. Virol. 2002, 76, 4267–4274. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.M.; Lauring, A.S.; Burns, C.C.; Overbaugh, J. Identification of a cellular cofactor required for infection by feline leukemia virus. Science 2000, 287, 1828–1830. [Google Scholar] [CrossRef]
- Lauring, A.S.; Anderson, M.; Overbaugh, J. Specificity in receptor usage by T-cell-tropic feline leukemia viruses: Implications for the in vivo tropism of immunodeficiency-inducing variants. J. Virol. 2001, 75, 8888–8898. [Google Scholar] [CrossRef] [Green Version]
- Lauring, A.S.; Cheng, H.H.; Eiden, M.V.; Overbaugh, J. Genetic and biochemical analyses of receptor and cofactor determinants for T-cell-tropic feline leukemia virus infection. J. Virol. 2002, 76, 8069–8078. [Google Scholar] [CrossRef] [Green Version]
- Lavillette, D.; Boson, B.; Russell, S.J.; Cosset, F.-L. Activation of membrane fusion by murine leukemia viruses is controlled in cis or in trans by interactions between the receptor-binding domain and a conserved disulfide loop of the carboxy terminus of the surface glycoprotein. J. Virol. 2001, 75, 3685–3695. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Lee, S.; Anderson, W. Functional interactions between monomers of the retroviral envelope protein complex. J. Virol. 1997, 71, 6967–6972. [Google Scholar] [CrossRef] [Green Version]
- Rein, A.; Yang, C.; Haynes, J.A.; Mirro, J.; Compans, R.W. Evidence for cooperation between murine leukemia virus Env molecules in mixed oligomers. J. Virol. 1998, 72, 3432–3435. [Google Scholar] [CrossRef] [Green Version]
- Koo, H.M.; Gu, J.; Varela-Echavarria, A.; Ron, Y.; Dougherty, J.P. Reticuloendotheliosis type C and primate type D oncoretroviruses are members of the same receptor interference group. J. Virol. 1992, 66, 3448–3454. [Google Scholar] [CrossRef]
- Rasko, J.E.; Battini, J.L.; Gottschalk, R.J.; Mazo, I.; Miller, A.D. The RD114/simian type D retrovirus receptor is a neutral amino acid transporter. Proc. Natl. Acad. Sci. USA 1999, 96, 2129–2134. [Google Scholar] [CrossRef] [Green Version]
- Kewalramani, V.N.; Panganiban, A.; Emerman, M. Spleen necrosis virus, an avian immunosuppressive retrovirus, shares a receptor with the type D simian retroviruses. J. Virol. 1992, 66, 3026–3031. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, J.; Sugimoto, M.; Bernstein, H.; Jinno, Y.; Schust, D. A novel human endogenous retroviral protein inhibits cell-cell fusion. Sci Rep. 2013, 3, 1462. [Google Scholar] [CrossRef] [Green Version]
- Tailor, C.S.; Nouri, A.; Zhao, Y.; Takeuchi, Y.; Kabat, D. A sodium-dependent neutral-amino-acid transporter mediates infections of feline and baboon endogenous retroviruses and simian type D retroviruses. J. Virol. 1999, 73, 4470–4474. [Google Scholar] [CrossRef] [Green Version]
- Marin, M.; Tailor, C.S.; Nouri, A.; Kabat, D. Sodium-dependent neutral amino acid transporter type 1 is an auxiliary receptor for baboon endogenous retrovirus. J. Virol. 2000, 74, 8085–8093. [Google Scholar] [CrossRef] [Green Version]
- Manel, N.; Kinet, S.; Battini, J.-L.; Kim, F.J.; Taylor, N.; Sitbon, M. The HTLV receptor is an early T-cell activation marker whose expression requires de novo protein synthesis. Blood 2003, 101, 1913–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hein, S.; Prassolov, V.; Zhang, Y.; Ivanov, D.; Löhler, J.; Ross, S.R.; Stocking, C. Sodium-dependent myo-inositol transporter 1 is a cellular receptor for Mus cervicolor M813 murine leukemia virus. J. Virol. 2003, 77, 5926–5932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tipper, C.H.; Cingoz, O.; Coffin, J. Mus spicilegus endogenous retrovirus HEMV uses murine sodium-dependent myo-inositol transporter 1 as a receptor. J. Virol. 2012, 86, 6341–6344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albritton, L.M.; Tseng, L.; Scadden, D.; Cunningham, J.M. A putative murine ecotropic retrovirus receptor gene encodes a multiple membrane-spanning protein and confers susceptibility to virus infection. Cell 1989, 57, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Sato, H.; Kubo, Y.; Wada, S.; Aida, Y. CAT1/SLC7A1 acts as a cellular receptor for bovine leukemia virus infection. FASEB J. 2019, 33, 14516–14527. [Google Scholar] [CrossRef]
- Chai, N.; Bates, P. Na+/H+ exchanger type 1 is a receptor for pathogenic subgroup J avian leukosis virus. Proc. Natl. Acad. Sci. USA 2006, 103, 5531–5536. [Google Scholar] [CrossRef] [Green Version]
- Tsang, J.; Ribet, D.; Heidmann, T.; Dewannieux, M. Identification of the Receptor Used by the Ecotropic Mouse GLN Endogenous Retrovirus. J. Virol. 2019, 93, e01125-18. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Stadler, C.K.; Gorman, K.; Jensen, N.; Kim, D.; Zheng, H.; Tang, S.; Switzer, W.M.; Pye, G.W.; Eiden, M.V. An exogenous retrovirus isolated from koalas with malignant neoplasias in a US zoo. Proc. Natl. Acad. Sci. USA 2013, 110, 11547–11552. [Google Scholar] [CrossRef] [Green Version]
- Shojima, T.; Yoshikawa, R.; Hoshino, S.; Shimode, S.; Nakagawa, S.; Ohata, T.; Nakaoka, R.; Miyazawa, T. Identification of a novel subgroup of Koala retrovirus from Koalas in Japanese zoos. J. Virol. 2013, 87, 9943–9948. [Google Scholar] [CrossRef] [Green Version]
- Mendoza, R.; Anderson, M.; Overbaugh, J. A putative thiamine transport protein is a receptor for feline leukemia virus subgroup A. J. Virol. 2006, 80, 3378–3385. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, N.M.; Farrell, K.; Eiden, M. In vitro characterization of a koala retrovirus. J. Virol. 2006, 80, 3104–3107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, Y.; Vile, R.G.; Simpson, G.; O’Hara, B.; Collins, M.K.; A Weiss, R. Feline leukemia virus subgroup B uses the same cell surface receptor as gibbon ape leukemia virus. J. Virol. 1992, 66, 1219–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olah, Z.; Lehel, C.; Anderson, W.B.; Eiden, M.V.; A Wilson, C. The cellular receptor for gibbon ape leukemia virus is a novel high affinity sodium-dependent phosphate transporter. J. Biol. Chem. 1994, 269, 25426–25431. [Google Scholar] [CrossRef] [PubMed]
- Kavanaugh, M.P.; Miller, D.G.; Zhang, W.; Law, W.; Kozak, S.L.; Kabat, D.; Miller, A.D. Cell-surface receptors for gibbon ape leukemia virus and amphotropic murine retrovirus are inducible sodium-dependent phosphate symporters. Proc. Natl. Acad. Sci. USA 1994, 91, 7071–7075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, C.A.; Eiden, M.V.; Anderson, W.B.; Lehel, C.; Olah, Z. The dual-function hamster receptor for amphotropic murine leukemia virus (MuLV), 10A1 MuLV, and gibbon ape leukemia virus is a phosphate symporter. J. Virol. 1995, 69, 534–537. [Google Scholar] [CrossRef]
- Soll, S.J.; Neil, S.; Bieniasz, P. Identification of a receptor for an extinct virus. Proc. Natl. Acad. Sci. USA 2010, 107, 19496–19501. [Google Scholar] [CrossRef] [Green Version]
- Tury, S.; Giovannini, D.; Ivanova, S.; Touhami, J.; Courgnaud, V.; Battini, J.-L. Identification of Copper Transporter 1 as a Receptor for Feline Endogenous Retrovirus ERV-DC14. J. Virol. 2022, 96, e0022922. [Google Scholar] [CrossRef]
- Sarangi, A.; Bupp, K.; Roth, M. Identification of a retroviral receptor used by an envelope protein derived by peptide library screening. Proc. Natl. Acad. Sci. USA 2007, 104, 11032–11037. [Google Scholar] [CrossRef] [Green Version]
- Quigley, J.G.; Burns, C.C.; Anderson, M.M.; Lynch, E.D.; Sabo, K.M.; Overbaugh, J.; Abkowitz, J.L. Cloning of the cellular receptor for feline leukemia virus subgroup C (FeLV-C), a retrovirus that induces red cell aplasia. Blood 2000, 95, 1093–1099. [Google Scholar] [CrossRef]
- Ericsson, T.A.; Takeuchi, Y.; Templin, C.; Quinn, G.; Farhadian, S.F.; Wood, J.C.; Oldmixon, B.A.; Suling, K.M.; Ishii, J.K.; Kitagawa, Y.; et al. Identification of receptors for pig endogenous retrovirus. Proc. Natl. Acad. Sci. USA 2003, 100, 6759–6764. [Google Scholar] [CrossRef] [Green Version]
- Mazari, P.M.; Argaw, T.; Valdivieso, L.; Zhang, X.; Marcucci, K.T.; Salomon, D.R.; Wilson, C.A.; Roth, M.J. Comparison of the convergent receptor utilization of a retargeted feline leukemia virus envelope with a naturally-occurring porcine endogenous retrovirus A. Virology 2012, 427, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Tailor, C.S.; Nouri, A.; Lee, C.G.; Kozak, C.; Kabat, D. Cloning and characterization of a cell surface receptor for xenotropic and polytropic murine leukemia viruses. Proc. Natl. Acad. Sci. USA 1999, 96, 927–932. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.L.; Guo, L.; Xu, S.; Holland, C.A.; Kitamura, T.; Hunter, K.W.; Cunningham, J.M. Receptors for polytropic and xenotropic mouse leukaemia viruses encoded by a single gene at Rmc1. Nat. Genet. 1999, 21, 216–219. [Google Scholar] [CrossRef]
- Esnault, C.; Priet, S.; Ribet, D.; Vernochet, C.; Bruls, T.; Lavialle, C.; Weissenbach, J.; Heidmann, T. A placenta-specific receptor for the fusogenic, endogenous retrovirus-derived, human syncytin-2. Proc. Natl. Acad. Sci. USA 2008, 105, 17532–17537. [Google Scholar] [CrossRef] [Green Version]
- Carette, J.E.; Raaben, M.; Wong, A.C.; Herbert, A.S.; Obernosterer, G.; Mulherkar, N.; Kuehne, A.I.; Kranzusch, P.J.; Griffin, A.M.; Ruthel, G.; et al. Ebola virus entry requires the cholesterol transporter Niemann-Pick C1. Nature 2011, 477, 340–343. [Google Scholar] [CrossRef]
- Bates, P.; Young, J.; Varmus, H. A receptor for subgroup A Rous sarcoma virus is related to the low density lipoprotein receptor. Cell 1993, 74, 1043–1051. [Google Scholar] [CrossRef]
- Young, J.A.; Bates, P.; Varmus, H. Isolation of a chicken gene that confers susceptibility to infection by subgroup A avian leukosis and sarcoma viruses. J. Virol. 1993, 67, 1811–1816. [Google Scholar] [CrossRef] [Green Version]
- Brojatsch, J.; Naughton, J.; Rolls, M.M.; Zingler, K.; Young, J.A. CAR1, a TNFR-related protein, is a cellular receptor for cytopathic avian leukosis-sarcoma viruses and mediates apoptosis. Cell 1996, 87, 845–855. [Google Scholar] [CrossRef] [Green Version]
- Elleder, D.; Stepanets, V.; Melder, D.C.; Šenigl, F.; Geryk, J.; Pajer, P.; Plachý, J.; Hejnar, J.; Svoboda, J.; Federspiel, M.J. The receptor for the subgroup C avian sarcoma and leukosis viruses, Tvc, is related to mammalian butyrophilins, members of the immunoglobulin superfamily. J. Virol. 2005, 79, 10408–10419. [Google Scholar] [CrossRef] [Green Version]
- Ferrada, E.; Superti-Furga, G. A structure and evolutionary-based classification of solute carriers. iScience 2022, 25, 105096. [Google Scholar] [CrossRef]
- Bacquin, A.; Bireau, C.; Tanguy, M.; Romanet, C.; Vernochet, C.; Dupressoir, A.; Heidmann, T. A Cell Fusion-Based Screening Method Identifies Glycosylphosphatidylinositol-Anchored Protein Ly6e as the Receptor for Mouse Endogenous Retroviral Envelope Syncytin-A. J. Virol. 2017, 91, e00832-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidmann, O.; Vernochet, C.; Dupressoir, A.; Heidmann, T. Identification of an endogenous retroviral envelope gene with fusogenic activity and placenta-specific expression in the rabbit: A new "syncytin" in a third order of mammals. Retrovirology 2009, 6, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazari, P.M.; Linder-Basso, D.; Sarangi, A.; Chang, Y.; Roth, M.J. Single-round selection yields a unique retroviral envelope utilizing GPR172A as its host receptor. Proc. Natl. Acad. Sci. USA 2009, 106, 5848–5853. [Google Scholar] [CrossRef] [Green Version]
- Mazari, P.M.; Roth, M.J. Library screening and receptor-directed targeting of gammaretroviral vectors. Future Microbiol. 2013, 8, 107–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydin, H.; Cook, J.; Lee, J. Crystal structures of beta- and gammaretrovirus fusion proteins reveal a role for electrostatic stapling in viral entry. J. Virol. 2014, 88, 143–153. [Google Scholar] [CrossRef]
- Lamb, D.; Schüttelkopf, A.W.; Van Aalten, D.M.F.; Brighty, D.W. Charge-surrounded pockets and electrostatic interactions with small ions modulate the activity of retroviral fusion proteins. PLoS Pathog. 2011, 7, e1001268. [Google Scholar] [CrossRef]
- Dean, T.T.; Serrao, V.; Lee, J. Structure of the Core Postfusion Porcine Endogenous Retrovirus Fusion Protein. mBio 2022, 13, e0292021. [Google Scholar] [CrossRef]
- Martinez-Molledo, M.; Nji, E.; Reyes, N. Structural insights into the lysophospholipid brain uptake mechanism and its inhibition by syncytin-2. Nat. Struct Mol. Biol. 2022, 29, 604–612. [Google Scholar] [CrossRef]
- Chen, C.P.; Chen, L.F.; Yang, S.R.; Chen, C.Y.; Ko, C.C.; Chang, G.D.; Chen, H. Functional characterization of the human placental fusogenic membrane protein syncytin 2. Biol. Reprod 2008, 79, 815–823. [Google Scholar] [CrossRef] [Green Version]
- Hotzel, I. Deep-Time Structural Evolution of Retroviral and Filoviral Surface Envelope Proteins. J. Virol. 2022, 96, e0006322. [Google Scholar] [CrossRef]
- Vogt, P.K.; Ishizaki, R. Patterns of viral interference in the avian leukosis and sarcoma complex. Virology 1966, 30, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Rein, A. Interference grouping of murine leukemia viruses: A distinct receptor for the MCF-recombinant viruses in mouse cells. Virology 1982, 120, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Rein, A.; Schultz, A. Different recombinant murine leukemia viruses use different cell surface receptors. Virology 1984, 136, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Sommerfelt, M.A.; Williams, B.P.; Clapham, P.R.; Solomon, E.; Goodfellow, P.N.; Weiss, R.A. Human T cell leukemia viruses use a receptor determined by human chromosome 17. Science 1988, 242, 1557–1559. [Google Scholar] [CrossRef] [PubMed]
- Barnard, R.J.; Elleder, D.; Young, J. Avian sarcoma and leukosis virus-receptor interactions: From classical genetics to novel insights into virus-cell membrane fusion. Virology 2006, 344, 25–29. [Google Scholar] [CrossRef]
- Wildum, S.; Schindler, M.; Münch, J.; Kirchhoff, F. Contribution of Vpu, Env, and Nef to CD4 down-modulation and resistance of human immunodeficiency virus type 1-infected T cells to superinfection. J. Virol. 2006, 80, 8047–8059. [Google Scholar] [CrossRef] [Green Version]
- Jobbagy, Z.; Garfield, S.; Baptiste, L.; Eiden, M.V.; Anderson, W.B. Subcellular redistribution of Pit-2 P(i) transporter/amphotropic leukemia virus (A-MuLV) receptor in A-MuLV-infected NIH 3T3 fibroblasts: Involvement in superinfection interference. J. Virol. 2000, 74, 2847–2854. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.W.; Cunningham, J.M. N-linked glycosylation of the receptor for murine ecotropic retroviruses is altered in virus-infected cells. J. Biol. Chem. 1993, 268, 16316–16320. [Google Scholar] [CrossRef]
- Bahrami, S.; Pagh, K.; Ejegod, D.; Duch, M.; Tolstrup, M.; Pedersen, F.S. Construction of a gammaretrovirus with a novel tropism and wild-type replication kinetics capable of using human APJ as entry receptor. J. Virol. 2012, 86, 10621–10627. [Google Scholar] [CrossRef] [Green Version]
- Delwart, E.L.; Panganiban, A.T. Role of reticuloendotheliosis virus envelope glycoprotein in superinfection interference. J. Virol. 1989, 63, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Bahrami, S.; Ejegod, D.; Sørensen, K.D.; Pedersen, F.S. Coupling of receptor interference and a host-dependent post-binding entry deficiency in a gammaretroviral envelope protein. Retrovirology 2010, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Eiden, M.V. A mutant retroviral receptor restricts virus superinfection interference and productive infection. Retrovirology 2012, 9, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Eiden, M.V. The receptors for gibbon ape leukemia virus and amphotropic murine leukemia virus are not downregulated in productively infected cells. Retrovirology 2011, 8, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, S.L.; Chung-Landers, M.; Honczarenko, M.; Gaulton, G.N. Linkage of reduced receptor affinity and superinfection to pathogenesis of TR1.3 murine leukemia virus. J. Virol. 2006, 80, 4601–4609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rainey, G.J.; Coffin, J.M. Evolution of broad host range in retroviruses leads to cell death mediated by highly cytopathic variants. J. Virol. 2006, 80, 562–570. [Google Scholar] [CrossRef]
- Reinhart, T.A.; Ghosh, A.K.; A Hoover, E.; I Mullins, J. Distinct superinfection interference properties yet similar receptor utilization by cytopathic and noncytopathic feline leukemia viruses. J. Virol. 1993, 67, 5153–5162. [Google Scholar] [CrossRef] [Green Version]
- Weller, S.K.; Joy, A.; Temin, H. Correlation between cell killing and massive second-round superinfection by members of some subgroups of avian leukosis virus. J. Virol. 1980, 33, 494–506. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, F.K.; Wang, T.; Nanua, S. Mink cell focus-forming murine leukemia virus killing of mink cells involves apoptosis and superinfection. J. Virol. 2001, 75, 6007–6015. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Liu, Q.; Wollenberg, K.; Martin, C.; Buckler-White, A.; Kozak, C.A. Evolution of functional and sequence variants of the mammalian XPR1 receptor for mouse xenotropic gammaretroviruses and the human-derived retrovirus XMRV. J. Virol. 2010, 84, 11970–11980. [Google Scholar] [CrossRef] [Green Version]
- Boso, G.; Lam, O.; Bamunusinghe, D.; Oler, A.J.; Wollenberg, K.; Liu, Q.; Shaffer, E.; Kozak, C.A. Patterns of Coevolutionary Adaptations across Time and Space in Mouse Gammaretroviruses and Three Restrictive Host Factors. Viruses 2021, 13, 1864. [Google Scholar] [CrossRef]
- Demogines, A.; Abraham, J.; Choe, H.; Farzan, M.; Sawyer, S.L. Dual host-virus arms races shape an essential housekeeping protein. PLoS Biol 2013, 11, e1001571. [Google Scholar] [CrossRef]
- Kizhatil, K.; Albritton, L.M. Requirements for different components of the host cell cytoskeleton distinguish ecotropic murine leukemia virus entry via endocytosis from entry via surface fusion. J. Virol. 1997, 71, 7145–7156. [Google Scholar] [CrossRef] [Green Version]
- Valle-Tenney, R.; Opazo, T.; Cancino, J.; Goff, S.P.; Arriagada, G. Dynein Regulators Are Important for Ecotropic Murine Leukemia Virus Infection. J. Virol. 2016, 90, 6896–6905. [Google Scholar] [CrossRef] [Green Version]
- Cheynet, V.; Ruggieri, A.; Oriol, G.; Blond, J.L.; Boson, B.; Vachot, L.; Verrier, B.; Cosset, F.-L.; Mallet, F. Synthesis, assembly, and processing of the Env ERVWE1/syncytin human endogenous retroviral envelope. J. Virol. 2005, 79, 5585–5593. [Google Scholar] [CrossRef] [Green Version]
- Dupressoir, A.; Vernochet, C.; Bawa, O.; Harper, F.; Pierron, G.; Opolon, P.; Heidmann, T. Syncytin-A knockout mice demonstrate the critical role in placentation of a fusogenic, endogenous retrovirus-derived, envelope gene. Proc. Natl. Acad. Sci. USA 2009, 106, 12127–12132. [Google Scholar] [CrossRef]
- Dupressoir, A.; Vernochet, C.; Harper, F.; Guegan, J.; Dessen, P.; Pierron, G.; Heidmann, T. A pair of co-opted retroviral envelope syncytin genes is required for formation of the two-layered murine placental syncytiotrophoblast. Proc. Natl. Acad. Sci. USA 2011, 108, E1164–E1173. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Receptor | Transporter of | Fold | # of Membrane-Spanning Domains | Virus | Classification | Source |
|---|---|---|---|---|---|---|
| Slc1a4 (ASCT1) | Neutral AA | Glt | 8 | REV | Gammaretrovirus | [185] |
| Slc1a4 (ASCT1) | Neutral AA | Glt | 8 | RD114 | Gammaretrovirus | [186] |
| Slc1a4 (ASCT1) | Neutral AA | Glt | 8 | SNV | Gammaretrovirus | [187] |
| Slc1a4/Slc1a5 (ASCT1/ASCT2) | Neutral AA | Glt | 8 | HERV-W/SYNCYTIN-1 | Gamma-like ERV | [50] |
| Slc1a5 (ASCT2) | Neutral AA | Glt | 8 | SUPYN | Gamma-like ERV | [188] |
| Slc1a5 (ASCT2) | Neutral AA | Glt | 8 | SRV1, SRV2, SRV3 (MPMV), SRV4, SRV5 | Betaretrovirus * | [189] |
| Slc1a5 (ASCT2) | Neutral AA | Glt | 8 | BaERV | Gammaretrovirus | [190] |
| Slc2A1 (Glut-1) | Glucose | MFS | 12 | HTLV | Deltaretrovirus | [191] |
| Slc5a3 (Smit1) | Sodium/myo-inositol | LeuT | 12 | M813 | Gammaretrovirus | [192] |
| Slc5a3 (Smit1) | Sodium/myo-inositol | LeuT | 12 | HEMV | Gammaretrovirus | [193] |
| Slc7a1 (mCAT1) | Cationic AA | LeuT | 12 | MoMLV | Gammaretrovirus | [194] |
| Slc7a1 (CAT1) | Cationic AA | LeuT | 12 | BLV | Deltaretrovirus | [195] |
| Slc9a1 (NHE-1) | Sodium/Hydrogen | NHE1 | 13 | ALV-J | Alpharetrovirus | [196] |
| Slc16a1 (MCT1) | monocarboxylate | MFS | 12 | HERV-T | Gammaretrovirus | [72] |
| Slc19a1 | Folate | MFS | 12 | GLN-MLV | Gammaretrovirus | [197] |
| Slc19a2 (THTR1) | Thiamine | MFS | 12 | KoRV-B | Gammaretrovirus | [198] |
| Slc19a2 (THTR1) | Thiamine | MFS | 12 | KoRV-J | Gammaretrovirus | [199] |
| Slc19a2 (THTR1) | Thiamine | MFS | 12 | FeLV-A | Gammaretrovirus | [200] |
| Slc20a1 (PiT1) | Phosphate | PiT | 10, 12 | KoRV-A | Gammaretrovirus | [201] |
| Slc20a1 (PiT1) | Phosphate | PiT | 10, 12 | FeLV-B | Gammaretrovirus | [202] |
| Slc20a1/Slc20a2 (PiT1/PiT2) | Phosphate | PiT | 10, 12 | GALV/WMV | Gammaretrovirus | [203] |
| Slc20a1 (PiT1) | Phosphate | PiT | 10, 12 | AMLV (4070A, 229A) | Gammaretrovirus | [204] |
| Slc20a1 (PiT1) | Phosphate | PiT | 10, 12 | FeLV-T | Gammaretrovirus | [179] |
| Slc20a1 (PiT1) | Phosphate | PiT | 10, 12 | 10A1-MLV | Gammaretrovirus | [205] |
| Slc31a1 (CTR1) | Copper ion | Ctr | 3 | cERV-1/2 | Gamma-like ERV | [206] |
| Slc31a1 (CTR1) | Copper ion | Ctr | 3 | FeLV-D | Gammaretrovirus | [207] |
| Slc35f2 | Nucleotide sugar | - | 10 | FeLV-A5 * | Gammaretrovirus | [208] |
| Slc49a1/Slc49a2 | Heme | MFS | 12 | FeLV-C | Gammaretrovirus | [209] |
| Slc52a2 (GHB/RFVT2/ PAR-2) | Riboflavin | MFS | 12 | PERV-A, FeLV-CP * | Gammaretrovirus | [210,211] |
| Slc53a1 (Xpr1) | Phosphate | Slc53 | 8 | XMLV | Gammaretrovirus | [212,213] |
| Slc53a1 (Xpr1) | Phosphate | Slc53 | 8 | PMLV | Gammaretrovirus | [212,213] |
| Slc59a1 (MFSD2A) | Lysophosphatidylcholine | MFS | 12 | HERV FRD/SYNCYTIN-2 | Gamma-like ERV | [214] |
| Slc65a1 (NPC1) | Cholesterol | NPC1 | 13 | Ebola virus | Non-retrovirus (filovirus) | [215] |
| TVA | Cobalamin (?) | LDLR | 1 | ALV-A/RSV-A | Alpharetrovirus | [216,217] |
| CAR1 (TVB) | Tumor necrosis factor | TNFR | 1 | ALV-B/D/E | Alpharetrovirus | [218] |
| TVC | Immunoglobulin | Ig | 1 | ALV-C | Alpharetrovirus | [219] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hogan, V.; Johnson, W.E. Unique Structure and Distinctive Properties of the Ancient and Ubiquitous Gamma-Type Envelope Glycoprotein. Viruses 2023, 15, 274. https://doi.org/10.3390/v15020274
Hogan V, Johnson WE. Unique Structure and Distinctive Properties of the Ancient and Ubiquitous Gamma-Type Envelope Glycoprotein. Viruses. 2023; 15(2):274. https://doi.org/10.3390/v15020274
Chicago/Turabian StyleHogan, Victoria, and Welkin E. Johnson. 2023. "Unique Structure and Distinctive Properties of the Ancient and Ubiquitous Gamma-Type Envelope Glycoprotein" Viruses 15, no. 2: 274. https://doi.org/10.3390/v15020274
APA StyleHogan, V., & Johnson, W. E. (2023). Unique Structure and Distinctive Properties of the Ancient and Ubiquitous Gamma-Type Envelope Glycoprotein. Viruses, 15(2), 274. https://doi.org/10.3390/v15020274