
Prevalence of HIV-1 Natural Polymorphisms and Integrase-Resistance-Associated Mutations in African Children
 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Sample Collection
2.2. Genotyping and Analysis of Resistance-Associated Mutations
2.3. Data Analysis
3. Results
3.1. Characteristics of Study Subjects
3.2. INI-Resistance-Associated Mutations and Natural Polymorphism Patterns
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ji, H.; Patterson, A.; Taylor, T.; Rank, C.; Halverson, J.; Capina, R.; Brooks, J.; Sandstrom, P. Prevalence of Primary Drug Resistance Against HIV-1 Integrase Inhibitors in Canada. Am. J. Ther. 2018, 78, e1–e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolakowska, A.; Maresca, A.F.; Collins, I.J.; Cailhol, J. Update on Adverse Effects of HIV Integrase Inhibitors. Curr. Treat. Options Infect. Dis. 2019, 11, 372–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Gu, S.-X.; He, Q.; Fan, R. Advances in the development of HIV integrase strand transfer inhibitors. Eur. J. Med. Chem. 2021, 225, 113787. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-W.; Shen, M.-C.; Wang, W.-H.; Li, W.-Y.; Wang, J.-H.; Tseng, C.-Y.; Liu, P.-Y.; Wang, L.-S.; Lee, Y.-L.; Chen, Y.-M.A.; et al. High prevalence of HIV-1 transmitted drug resistance and factors associated with time to virological failure and viral suppression in Taiwan. J. Antimicrob. Chemother. 2021, 77, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Park, T.E.; Mohamed, A.; Kalabalik, J.; Sharma, R. Review of integrase strand transfer inhibitors for the treatment of human immunodeficiency virus infection. Expert Rev. Anti-Infect. Ther. 2015, 13, 1195–1212. [Google Scholar] [CrossRef] [PubMed]
- Messiaen, P.; Wensing, A.M.J.; Fun, A.; Nijhuis, M.; Brusselaers, N.; Vandekerckhove, L. Clinical Use of HIV Integrase Inhibitors: A Systematic Review and Meta-Analysis. PLoS ONE 2013, 8, e52562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abram, M.E.; Ram, R.R.; Margot, N.A.; Barnes, T.L.; White, K.L.; Callebaut, C.; Miller, M.D. Lack of impact of pre-existing T97A HIV-1 integrase mutation on integrase strand transfer inhibitor resistance and treatment outcome. PLoS ONE 2017, 12, e0172206. [Google Scholar] [CrossRef] [Green Version]
- Wainberg, M.A.; Zaharatos, G.J.; Brenner, B.G. Development of Antiretroviral Drug Resistance. N. Engl. J. Med. 2011, 365, 637–646. [Google Scholar] [CrossRef]
- Giovanetti, M.; Ciccozzi, M.; Parolin, C.; Borsetti, A. Molecular Epidemiology of HIV-1 in African Countries: A Comprehensive Overview. Pathogens 2020, 9, 1072. [Google Scholar] [CrossRef]
- Jordan, M.R.; Penazzato, M.; Cournil, A.; Vubil, A.; Jani, I.; Hunt, G.; Carmona, S.; Maphalala, G.; Mthethwa, N.; Watera, C.; et al. Human Immunodeficiency Virus (HIV) Drug Resistance in African Infants and Young Children Newly Diagnosed With HIV: A Multicountry Analysis. Clin. Infect. Dis. 2017, 65, 2018–2025. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Updated Recommendations on First-Line and Secondline Antiretroviral Regimens and Post-Exposure Prophylaxis and Recommendations on Early Infant Diagnosis of HIV: Interim Guidelines; WHO: Geneva, Switzerland, 2018; Available online: https://www.who.int/publications/i/item/WHO-CDS-HIV-18.51 (accessed on 2 January 2023).
- Turkova, A.; White, E.; Mujuru, H.A.; Kekitiinwa, A.R.; Kityo, C.M.; Violari, A.; Lugemwa, A.; Cressey, T.R.; Musoke, P.; Variava, E.; et al. Dolutegravir as First- or Second-Line Treatment for HIV-1 Infection in Children. N. Engl. J. Med. 2021, 385, 2531–2543. [Google Scholar] [CrossRef] [PubMed]
- Mazzuti, L.; Melengu, T.; Falasca, F.; Calabretto, M.; Cella, E.; Ciccozzi, M.; Mezzaroma, I.; Iaiani, G.; Spaziante, M.; D’Ettorre, G.; et al. Transmitted drug resistance mutations and trends of HIV-1 subtypes in treatment-naïve patients: A single-centre experience. J. Glob. Antimicrob. Resist. 2019, 20, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Fofana, D.B.; D’Almeida, M.; Lambert-Niclot, S.; Peytavin, G.; Girard, P.M.; Lafia, B.; Zohoun-Guidigbi, L.; Keke, R.K.; Soulie, C.; Marcelin, A.G.; et al. Resistance profile and treatment outcomes in HIV-infected children at virological failure in Benin, West Africa. J. Antimicrob. Chemother. 2018, 73, 3143–3147. [Google Scholar] [CrossRef] [PubMed]
- Margot, N.A.; Hluhanich, R.M.; Jones, G.S.; Andreatta, K.N.; Tsiang, M.; McColl, D.J.; White, K.L.; Miller, M.D. In vitro resistance selections using elvitegravir, raltegravir, and two metabolites of elvitegravir M1 and M4. Antivir. Res. 2011, 93, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Lataillade, M.; Chiarella, J.; Kozal, M.J. Natural Polymorphism of the HIV-1 Integrase Gene and Mutations associated with Integrase Inhibitor Resistance. Antivir. Ther. 2007, 12, 563–570. [Google Scholar] [CrossRef]
- Standford University. HIV drug resistance Database: INI Mutation Pattern and Susceptibility. Updated on 2022-10-15. [Online]. 2022. Available online: https://hivdb.stanford.edu/ (accessed on 2 January 2023).
- Ambrosioni, J.; Nicolás, D.; Manzardo, C.; Agüero, F.; Blanco, J.L.; Mosquera, M.D.M.; Peñafiel, J.; Gatell, J.M.; Marcos, M.A.; Miró, J.M. Integrase strand-transfer inhibitor polymorphic and accessory resistance substitutions in patients with acute/recent HIV infection. J. Antimicrob. Chemother. 2016, 72, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Stekler, J.D.; McKernan, J.; Milne, R.; Tapia, K.A.; Mykhalchenko, K.; Holte, S.; Maenza, J.; Stevens, C.E.; Buskin, S.E.; Mullins, J.I.; et al. Lack of Resistance to Integrase Inhibitors among Antiretroviral-Naive Subjects with Primary HIV-1 Infection, 2007–2013. Antivir. Ther. 2015, 20, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, C.; Hernández-Novoa, B.; Pérez-Elías, M.J.; Moreno, A.M.; Holguín, Á.; Dronda, F.; Casado, J.L.; Moreno, S. Prevalence of Primary Resistance Mutations to Integrase Inhibitors in Treatment-Naïve and -Experienced Patients Infected with B and Non-B HIV-1 Variants. HIV Clin. Trials 2013, 14, 10–16. [Google Scholar] [CrossRef]
- Madyadi, A.; Dhoro, M.; Shamu, T.; Washaya, T.; Kouamou, M.V.; Chimukangara, B.; Katzenstein, D.; Manasa, J. HIV-1 Genetic Diversity and Natural Polymorphisms of the Integrase Gene in Integrase Inhibitor-Naive Patients in Harare, Zimbabwe. AIDS Res. Hum. Retroviruses 2021, 37, 954–961. [Google Scholar] [CrossRef]
- McCluskey, S.M.; Kamelian, K.; Musinguzi, N.; Kigozi, S.; Yap, B.O.U.M.; Bwana, M.B.; Muzoora, C.; Brumme, Z.L.; Carrington, M.; Carlson, J.; et al. Pre-treatment integrase inhibitor resistance is uncommon in ART-naïve individuals with HIV-1 subtype A1 and D infections in Uganda Suzanne. AIDS 2022, 35, 1083–1089. [Google Scholar] [CrossRef]
- Alaoui, N.; El Alaoui, M.A.; Touil, N.; El Annaz, H.; Melloul, M.; Tagajdid, R.; Hjira, N.; Boui, M.; El Fahime, E.M.; Mrani, S. Prevalence of resistance to integrase strand-transfer inhibitors (INSTIs) among untreated HIV-1 infected patients in Morocco. BMC Res. Notes 2018, 11, 369. [Google Scholar] [CrossRef] [PubMed]
- Acharya, A.; Tagny, C.T.; Mbanya, D.; Fonsah, J.Y.; Nchindap, E.; Kenmogne, L.; Jihyun, M.; Njamnshi, A.K.; Kanmogne, G.D. Variability in HIV-1 Integrase Gene and 3′-Polypurine Tract Sequences in Cameroon Clinical Isolates, and Implications for Integrase Inhibitors Efficacy. Int. J. Mol. Sci. 2020, 21, 1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inzaule, S.C.; Hamers, R.L.; Noguera-Julian, M.; Casadellà, M.; Parera, M.; de Wit, T.F.R.; Paredes, R. Primary resistance to integrase strand transfer inhibitors in patients infected with diverse HIV-1 subtypes in sub-Saharan Africa. J. Antimicrob. Chemother. 2018, 73, 1167–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, A.J.; Rhee, S.-Y.; Shafer, R.W. Integrase Strand Transfer Inhibitor Resistance in Integrase Strand Transfer Inhibitor-Naive Persons. AIDS Res. Hum. Retroviruses 2021, 37, 736–743. [Google Scholar] [CrossRef]
- Arimide, D.A.; Szojka, Z.I.; Zealiyas, K.; Gebreegziabxier, A.; Adugna, F.; Sasinovich, S.; Björkman, P.; Medstrand, P. Pre-Treatment Integrase Inhibitor Resistance and Natural Polymorphisms among HIV-1 Subtype C Infected Patients in Ethiopia. Viruses 2022, 14, 729. [Google Scholar] [CrossRef]
- Rhee, S.-Y.; Tzou, P.; Shafer, R. Temporal Trends in HIV-1 Mutations Used for the Surveillance of Transmitted Drug Resistance. Viruses 2021, 13, 879. [Google Scholar] [CrossRef]
- Ceccherini-Silberstein, F.; Malet, I.; Fabeni, L.; Dimonte, S.; Svicher, V.; D’Arrigo, R.; Artese, A.; Costa, G.; Bono, S.; Alcaro, S.; et al. Specific HIV-1 integrase polymorphisms change their prevalence in untreated versus antiretroviral-treated HIV-1-infected patients, all naive to integrase inhibitors. J. Antimicrob. Chemother. 2010, 65, 2305–2318. [Google Scholar] [CrossRef]
- Low, A.; Prada, N.; Topper, M.; Vaida, F.; Castor, D.; Mohri, H.; Hazuda, D.; Muesing, M.; Markowitz, M. Natural Polymorphisms of Human Immunodeficiency Virus Type 1 Integrase and Inherent Susceptibilities to a Panel of Integrase Inhibitors. Antimicrob. Agents Chemother. 2009, 53, 4275–4282. [Google Scholar] [CrossRef] [Green Version]
- Hachiya, A.; Ode, H.; Matsuda, M.; Kito, Y.; Shigemi, U.; Matsuoka, K.; Imamura, J.; Yokomaku, Y.; Iwatani, Y.; Sugiura, W. Natural polymorphism S119R of HIV-1 integrase enhances primary INSTI resistance. Antivir. Res. 2015, 119, 84–88. [Google Scholar] [CrossRef]
- Tang, M.W.; Liu, T.F.; Shafer, R.W. The HIVdb System for HIV-1 Genotypic Resistance Interpretation. Intervirology 2012, 55, 98–101. [Google Scholar] [CrossRef]
- Doyle, T.; Dunn, D.T.; Ceccherini-Silberstein, F.; De Mendoza, C.; Garcia, F.; Smit, E.; Fearnhill, E.; Marcelin, A.-G.; Martinez-Picado, J.; Kaiser, R.; et al. Integrase inhibitor (INI) genotypic resistance in treatment-naive and raltegravir-experienced patients infected with diverse HIV-1 clades. J. Antimicrob. Chemother. 2015, 70, 3080–3086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anstett, K.; Cutillas, V.; Fusco, R.; Mesplède, T.; Wainberg, M.A. Polymorphic substitution E157Q in HIV-1 integrase increases R263K-mediated dolutegravir resistance and decreases DNA binding activity. J. Antimicrob. Chemother. 2016, 71, 2083–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, S.-Y.; Sankaran, K.; Varghese, V.; Winters, M.A.; Hurt, C.B.; Eron, J.J.; Parkin, N.; Holmes, S.P.; Holodniy, M.; Shafer, R.W. HIV-1 Protease, Reverse Transcriptase, and Integrase Variation. J. Virol. 2016, 90, 6058–6070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, C.; Geretti, A.M.; Zahonero, N.; Booth, C.; Strang, A.; Soriano, V.; De Mendoza, C. Integrase variability and susceptibility to HIV integrase inhibitors: Impact of subtypes, antiretroviral experience and duration of HIV infection. J. Antimicrob. Chemother. 2009, 65, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Sichtig, N.; Sierra, S.; Kaiser, R.; Däumer, M.; Reuter, S.; Schülter, E.; Altmann, A.; Fätkenheuer, G.; Dittmer, U.; Pfister, H.; et al. Evolution of raltegravir resistance during therapy. J. Antimicrob. Chemother. 2009, 64, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Armenia, D.; Fabeni, L.; Alteri, C.; Di Pinto, D.; Di Carlo, D.; Bertoli, A.; Gori, C.; Carta, S.; Fedele, V.; Forbici, F.; et al. HIV-1 integrase genotyping is reliable and reproducible for routine clinical detection of integrase resistance mutations even in patients with low-level viraemia. J. Antimicrob. Chemother. 2015, 70, 1865–1873. [Google Scholar] [CrossRef] [Green Version]
- Abram, M.E.; Hluhanich, R.M.; Goodman, D.D.; Andreatta, K.N.; Margot, N.A.; Ye, L.; Niedziela-Majka, A.; Barnes, T.L.; Novikov, N.; Chen, X.; et al. Impact of Primary Elvitegravir Resistance-Associated Mutations in HIV-1 Integrase on Drug Susceptibility and Viral Replication Fitness. Antimicrob. Agents Chemother. 2013, 57, 2654–2663. [Google Scholar] [CrossRef] [Green Version]
- Molina, J.-M.; LaMarca, A.; Andrade-Villanueva, J.; Clotet, B.; Clumeck, N.; Liu, Y.-P.; Zhong, L.; Margot, N.; Cheng, A.K.; Chuck, S.L. Efficacy and safety of once daily elvitegravir versus twice daily raltegravir in treatment-experienced patients with HIV-1 receiving a ritonavir-boosted protease inhibitor: Randomised, double-blind, phase 3, non-inferiority study. Lancet Infect. Dis. 2012, 12, 27–35. [Google Scholar] [CrossRef]
- Cooper, D.A.; Steigbigel, R.T.; Gatell, J.M.; Rockstroh, J.K.; Katlama, C.; Yeni, P.; Lazzarin, A.; Clotet, B.; Kumar, P.N.; Eron, J.E.; et al. Subgroup and Resistance Analyses of Raltegravir for Resistant HIV-1 Infection. N. Engl. J. Med. 2008, 359, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Eron, J.J.; Clotet, B.; Durant, J.; Katlama, C.; Kumar, P.; Lazzarin, A.; Poizot-Martin, I.; Richmond, G.; Soriano, V.; Ait-Khaled, M.; et al. Safety and Efficacy of Dolutegravir in Treatment-Experienced Subjects with Raltegravir-Resistant HIV Type 1 Infection: 24-Week Results of the VIKING Study. J. Infect. Dis. 2012, 207, 740–748. [Google Scholar] [CrossRef]
- Vavro, C.; Ruel, T.; Wiznia, A.; Montañez, N.; Nangle, K.; Horton, J.; Buchanan, A.M.; Stewart, E.L.; Palumbo, P. Emergence of Resistance in HIV-1 Integrase with Dolutegravir Treatment in a Pediatric Population from the IMPAACT P1093 Study. Antimicrob. Agents Chemother. 2022, 66, e01645-21. [Google Scholar] [CrossRef] [PubMed]
- Varghese, V.; Liu, T.F.; Rhee, S.-Y.; Libiran, P.; Trevino, C.; Fessel, W.J.; Shafer, R.W. HIV-1 Integrase Sequence Variability in Antiretroviral Naïve Patients and in Triple-Class Experienced Patients Subsequently Treated with Raltegravir. AIDS Res. Hum. Retroviruses 2010, 26, 1323–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diarra, H.; Makhulu, E.E.; Odhiambo, P.O.; Irekwa, R.M.; Kinyua, J.; Herren, J.K.; Mobegi, V.A. Molecular Investigation of Genetic Signatures of Selection in Plasmodium falciparum Actin-Binding Protein Coronin, Cysteine Desulfurase, and Plasmepsin 2 Gene in Mbita Field Isolates, Western Kenya. Open J. Genet. 2021, 11, 120–144. [Google Scholar] [CrossRef]
- Boerma, R.S.; Sigaloff, K.C.E.; Akanmu, A.S.; Inzaule, S.; van Hensbroek, M.B.; de Wit, T.F.R.; Calis, J.C. Alarming increase in pretreatment HIV drug resistance in children living in sub-Saharan Africa: A systematic review and meta-analysis. J. Antimicrob. Chemother. 2016, 72, 365–371. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Characteristics | ART-Naïve (n = 44) | ART-Treated (n = 31) | Total | p-Value |
|---|---|---|---|---|
| Gender, n (%) | ||||
| Male | 11 (25.0) | 24 (77.4) | 35/75 (46.6%) | <0.0001 ** |
| Female | 21 (47.7) | 7 (22.6) | 28/75 (37.4%) | |
| Not available | 12 (27.3) | −0 (0.0) | 12/75 (16%) | |
| Age, year, median (IQR) | 2.6 (1.5–5.0) | 10.0 (6.0–13.0) | 0.0005 | |
| CD4 cell/mm3, median (IQR) | 538 | 362 | 0.0659 | |
| (341–1003) | (170–607) | |||
| RNA HIV-1 copies/mL, median (IQR) * | ND | 54,000 (5543–170,000) | ||
| Duration of treatment, years, median (IQR) * | NA | 5 (3–7) | ||
| Treatment at enrollment *, n (%) | ||||
| 3TC + (ZDV or ABC) + NVP or EFV | NA | 28 (90.3) | ||
| TDF + (3TC or FTC or ABC) + LPV/r | NA | 3 (9.7) | ||
| Subtypes, n (%) | ||||
| CRF02_AG | 34 | 22 (71.0) | 56/75 (74.7%) | 0.5957 *** |
| CRF06_cpx | 9 | 6 (19.3) | 15/75 (20.0%) | |
| Other | 0 | 3 (9.7) | 3/75 (5.3%) | |
| INI accessory polymorphism mutations at described positions, n (%) **** | 15 (34.1) | 11 (35.5) | 1.0000 | |
| Other polymorphism mutations, n (%) **** | 38 (86.4) | 17 (54.8) | 0.0035 |
| Mutations | ART-Naïve | ART-Treated | Number Totals of Sequences with at Least One Mutation |
|---|---|---|---|
| INI accessory polymorphism mutations at described positions, n (%) | |||
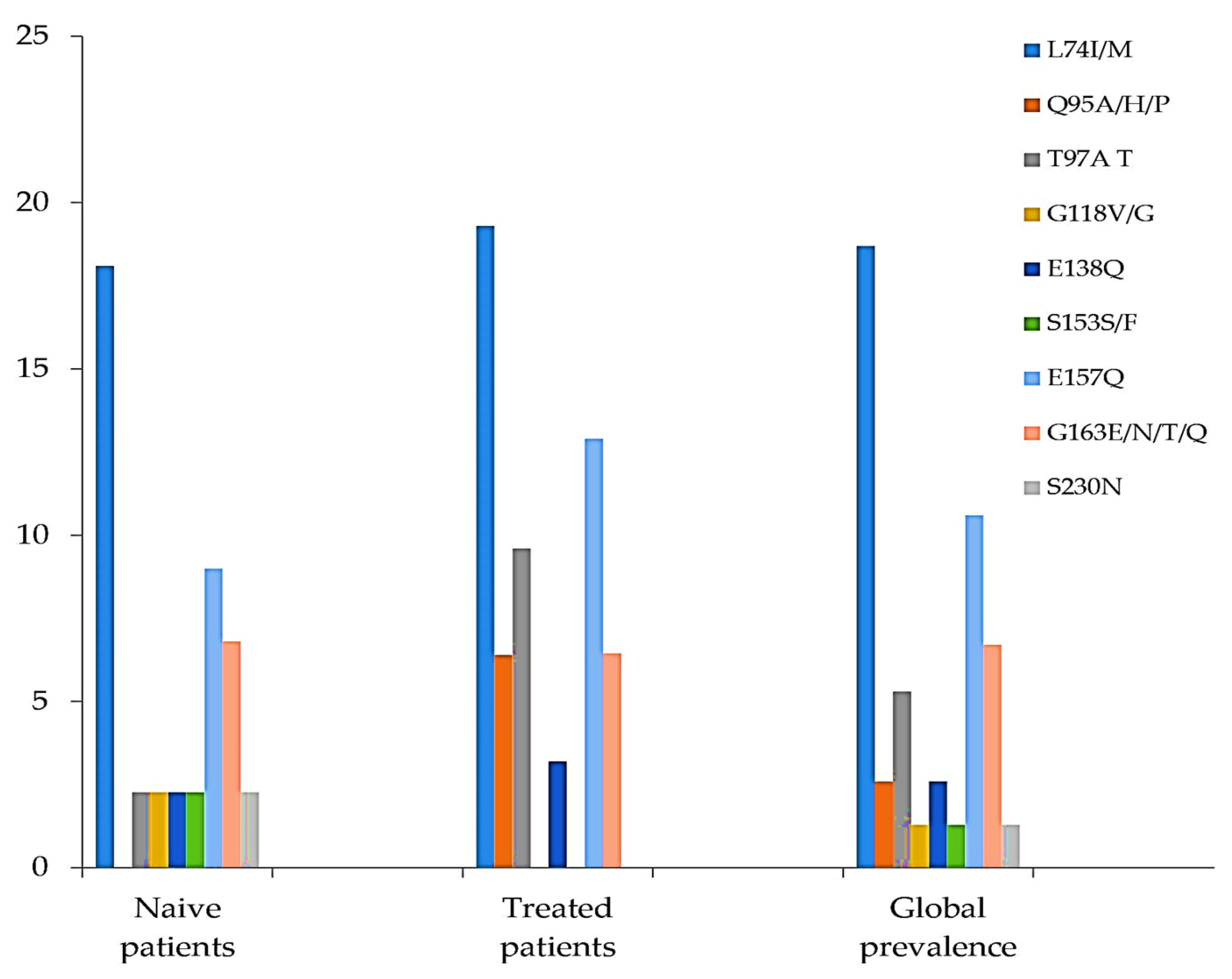
| L74I/M | 8/44 (18.1) | 6/31 (19.3) | 14/75 (18.7) |
| Q95A/H/P | 0 | 2/31 (6.4) | 2/75 (2.6) |
| T97A/T | 1/44 (2.3) | 3/31 (9.6) | 4/75 (5.3) |
| G118V/G | 1/44 (2.3) | 0 | 1/75 (1.3) |
| E138Q | 1/44 (2.3) | 1/31(3.2) | 2/75 (2.6) |
| S153F | 1/44 (2.3) | 0 | 1/75 (1.3) |
| E157Q | 4/44 (9) | 4/31 (12.9) | 8/75 (10.6) |
| G163E/N/T/Q | 3/44 (6.8) | 2/31 (6.4) | 5/75 (6.7) |
| S230N | 1/44 (2.3) | 0 | 1/75 (1.3) |
| Other polymorphism mutations, n (%) | |||
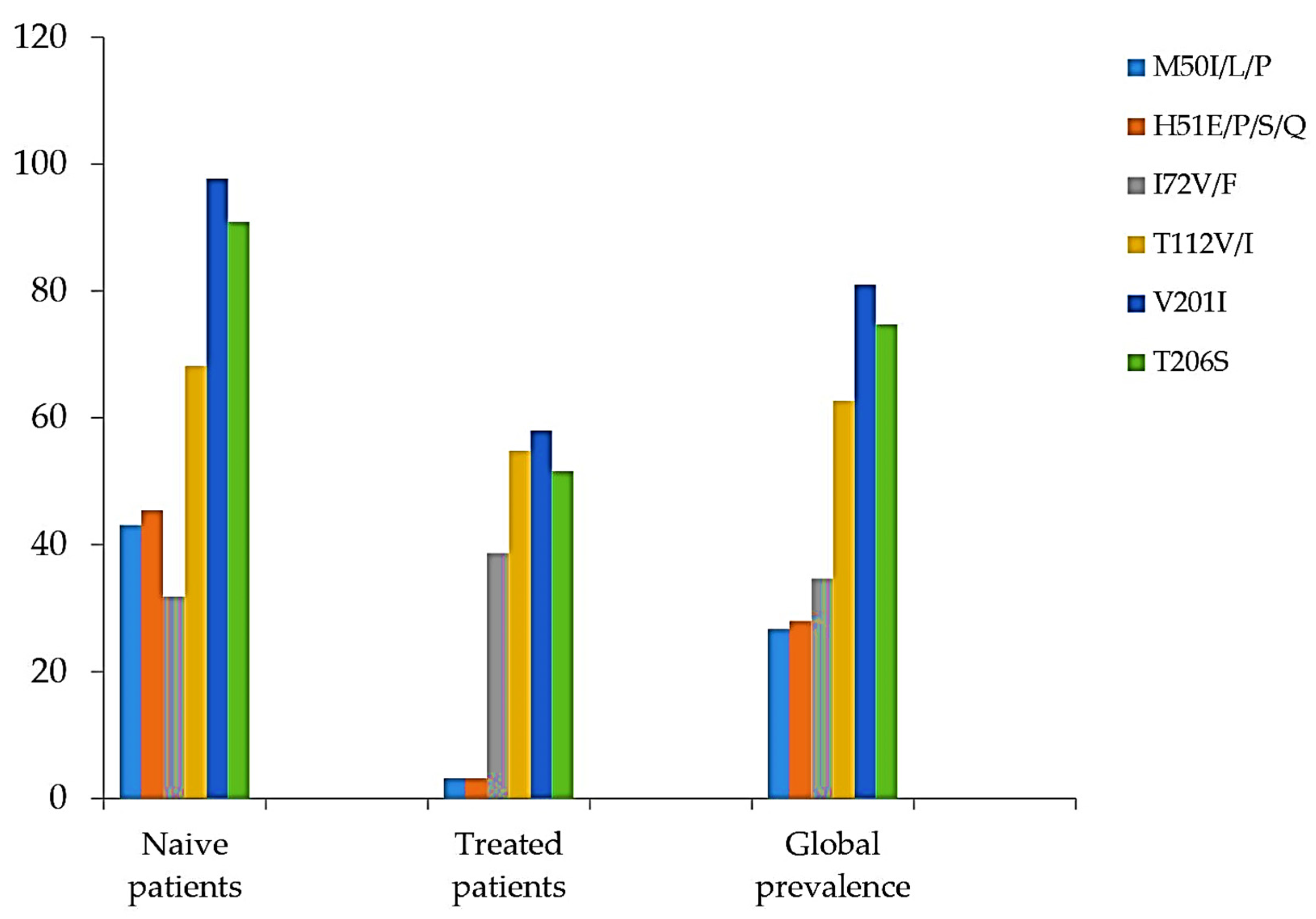
| M50I/L/P | 19/44 (43.1) | 1/31 (3.2) | 20/75 (26.7) |
| H51E/P/S/Q | 20/44 (45.4) | 1/31 (3.2) | 21/75 (28) |
| I72V/F | 14/44 (31.8) | 12/31 (38.7) | 26/75 (34.7) |
| T112V/I | 30/44 (68.1) | 17/31 (54.8) | 47/75 (62.7) |
| V201I | 43/44 (97.7) | 18/31 (58.0) | 61/75 (81) |
| T206S | 40/44 (90.9) | 16/31 (51.6) | 56/75 (74.7) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fofana, D.B.; Diarra, H.; Guindo, I.; Savadogo, M.K.; d’Almeida, M.; Diallo, F.I.; Baldé, A.; Soulié, C.; Kone, A.; Marcelin, A.-G.; et al. Prevalence of HIV-1 Natural Polymorphisms and Integrase-Resistance-Associated Mutations in African Children. Viruses 2023, 15, 546. https://doi.org/10.3390/v15020546
Fofana DB, Diarra H, Guindo I, Savadogo MK, d’Almeida M, Diallo FI, Baldé A, Soulié C, Kone A, Marcelin A-G, et al. Prevalence of HIV-1 Natural Polymorphisms and Integrase-Resistance-Associated Mutations in African Children. Viruses. 2023; 15(2):546. https://doi.org/10.3390/v15020546
Chicago/Turabian StyleFofana, Djeneba B., Houdou Diarra, Ibrahima Guindo, Mahamadou K. Savadogo, Marceline d’Almeida, Fatoumata I. Diallo, Aliou Baldé, Cathia Soulié, Amadou Kone, Anne-Geneviève Marcelin, and et al. 2023. "Prevalence of HIV-1 Natural Polymorphisms and Integrase-Resistance-Associated Mutations in African Children" Viruses 15, no. 2: 546. https://doi.org/10.3390/v15020546
APA StyleFofana, D. B., Diarra, H., Guindo, I., Savadogo, M. K., d’Almeida, M., Diallo, F. I., Baldé, A., Soulié, C., Kone, A., Marcelin, A.-G., Maiga, A. I., Lambert-Niclot, S., Maiga, M., McFall, S., Hawkins, C. A., Murphy, R. L., Sylla, M., Katlama, C., Holl, J. L., ... Morand-Joubert, L. (2023). Prevalence of HIV-1 Natural Polymorphisms and Integrase-Resistance-Associated Mutations in African Children. Viruses, 15(2), 546. https://doi.org/10.3390/v15020546