

Low Prevalence of SARS-CoV-2 Antibodies in Canine and Feline Serum Samples Collected during the COVID-19 Pandemic in Hong Kong and Korea

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Description
2.1.1. Seoul, Korea
2.1.2. Hong Kong, China
2.2. Assay Description and Optimisation
2.3. Confirmatory Serological Tests: Surrogate Virus Neutralization and Plaque Reduction Neutralization Tests
2.4. Detection of SARS-CoV-2 RNA Shedding in Oronasal Secretions of Healthy-Free Roaming Cats in Hong Kong by RT-PCR
2.5. Statistical Analyses
3. Results
3.1. Intra- and Inter-Assay Coefficients of Variation (CV)
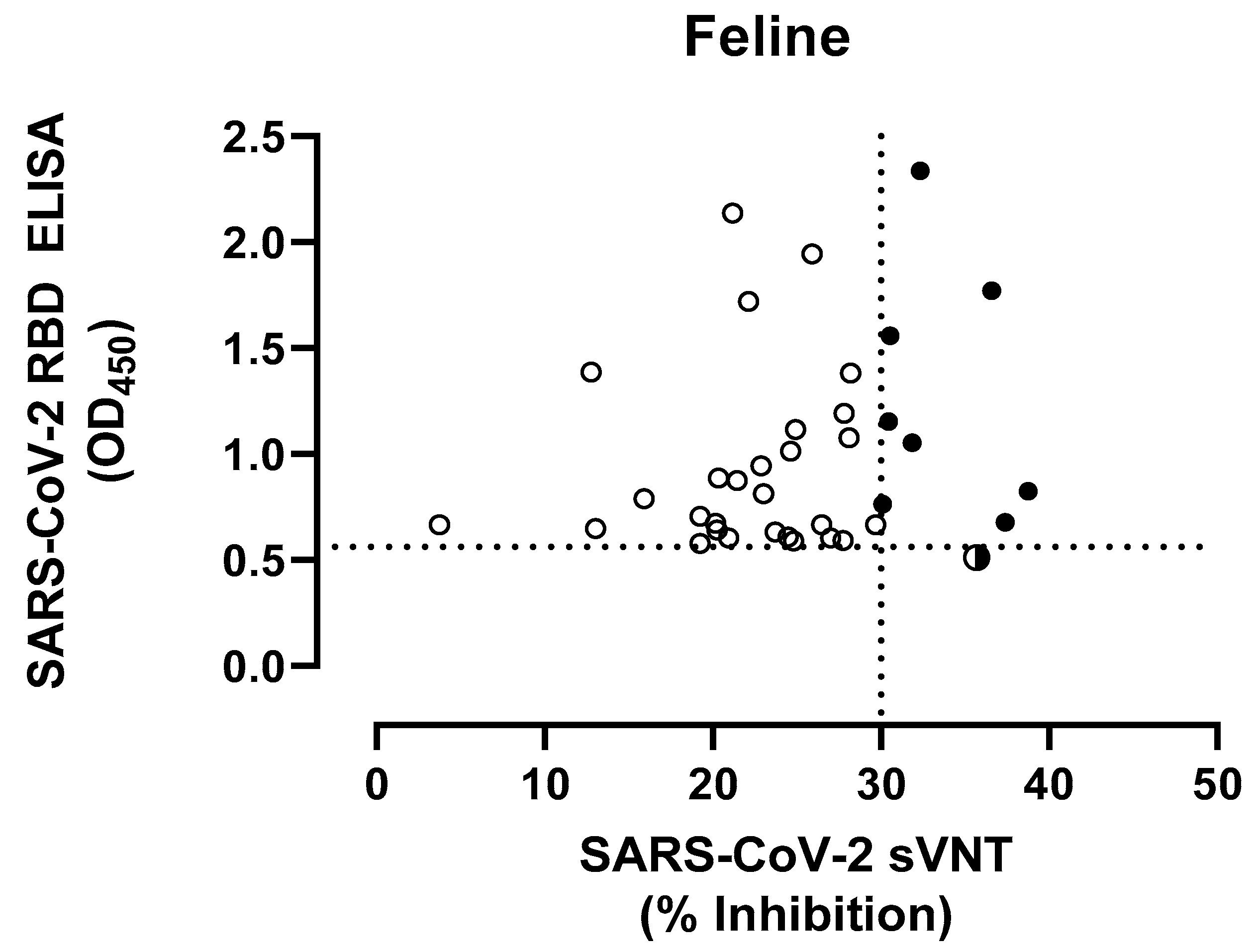
3.2. SARS-CoV-2 Seroprevalence in Domestic Cats
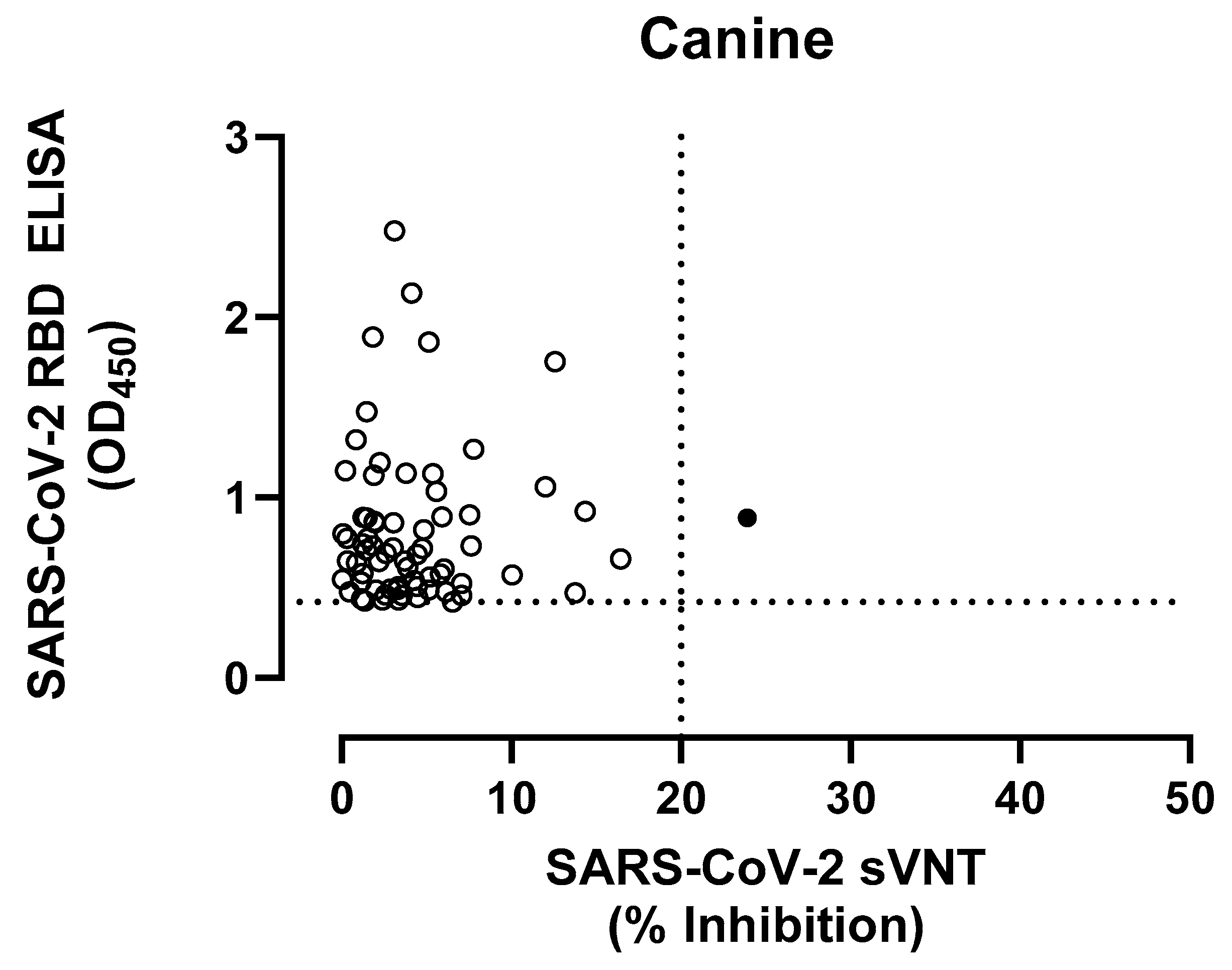
3.3. SARS-CoV-2 Seroprevalence in Domestic Dogs
3.4. Prevalence of SARS-CoV-2 RNA Shedding in Oronasal Secretions of Healthy-Free Roaming Cats in Hong Kong
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ludwig, S.; Zarbock, A. Coronaviruses and SARS-CoV-2: A Brief Overview. Anesth. Analg. 2020, 131, 93–96. [Google Scholar] [CrossRef]
- Voskarides, K. Animal-to-Human Viral Transitions: Is SARS-CoV-2 an Evolutionarily Successful One? J. Mol. Evol. 2020, 88, 421–423. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Li, T.; Zhang, S.; Wang, L.; Wu, X.; Liu, J. The genetic sequence, origin, and diagnosis of SARS-CoV-2. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1629–1635. [Google Scholar] [CrossRef]
- Guan, Y.; Zheng, B.J.; He, Y.Q.; Liu, X.L.; Zhuang, Z.X.; Cheung, C.L.; Luo, S.W.; Li, P.H.; Zhang, L.J. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 2003, 302, 276–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavanagh, D. Nidovirales: A new order comprising Coronaviridae and Arteriviridae. Arch. Virol. 1997, 142, 629–633. [Google Scholar] [PubMed]
- Graham, R.L.; Baric, R.S. Recombination, reservoirs, and the modular spike: Mechanisms of coronavirus cross-species transmission. J. Virol. 2010, 84, 3134–3146. [Google Scholar] [CrossRef] [Green Version]
- Fung, T.S.; Liu, D.X. Human Coronavirus: Host-Pathogen Interaction. Annu. Rev. Microbiol. 2019, 73, 529–557. [Google Scholar] [CrossRef] [Green Version]
- de Wit, E.; van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef]
- Wong, G.; Bi, Y.H.; Wang, Q.H.; Chen, X.W.; Zhang, Z.G.; Yao, Y.G. Zoonotic origins of human coronavirus 2019 (HCoV-19/SARS-CoV-2): Why is this work important? Zool. Res. 2020, 41, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Azhar, E.I.; El-Kafrawy, S.A.; Farraj, S.A.; Hassan, A.M.; Al-Saeed, M.S.; Hashem, A.M.; Madani, T.A. Evidence for camel-to-human transmission of MERS coronavirus. N. Engl. J. Med. 2014, 370, 2499–2505. [Google Scholar] [CrossRef] [PubMed]
- Reusken, C.B.; Haagmans, B.L.; Muller, M.A.; Gutierrez, C.; Godeke, G.J.; Meyer, B.; Muth, D.; Raj, V.S.; Smits-De Vries, L.; Corman, V.M.; et al. Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: A comparative serological study. Lancet Infect. Dis. 2013, 13, 859–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latif, A.A.; Mukaratirwa, S. Zoonotic origins and animal hosts of coronaviruses causing human disease pandemics: A review. Onderstepoort. J. Vet. Res. 2020, 87, e1–e9. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sit, T.H.C.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; YU, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; et al. Infection of dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef] [PubMed]
- Barrs, V.R.; Peiris, M.; Tam, K.W.S.; Law, P.Y.T.; Brackman, C.J.; To, E.M.W.; Yu, V.Y.T.; Chu, D.K.W.; Perera, R.; Sit, T.H.C. SARS-CoV-2 in Quarantined Domestic Cats from COVID-19 Households or Close Contacts, Hong Kong, China. Emerg. Infect. Dis. 2020, 26, 3071–3074. [Google Scholar] [CrossRef]
- Newman, A.; Smith, D.; Ghai, R.R.; Wallace, R.M.; Torchetti, M.K.; Loiacono, C.; Murrell, L.S.; Carpenter, A.; Moroff, S.; Rooney, J.A.; et al. First Reported Cases of SARS-CoV-2 Infection in Companion Animals—New York, March-April 2020. MMWR Morb. Mortal Wkly. Rep. 2020, 69, 710–713. [Google Scholar] [CrossRef] [PubMed]
- Prince, T.; Smith, S.L.; Radford, A.D.; Solomon, T.; Hughes, G.L.; Patterson, E.I. SARS-CoV-2 Infections in Animals: Reservoirs for Reverse Zoonosis and Models for Study. Viruses 2021, 13, 494. [Google Scholar] [CrossRef]
- Kok, K.H.; Wong, S.C.; Chan, W.M.; Wen, L.; Chu, A.W.; Ip, J.D.; Lee, L.K.; Wong, I.T.; Lo, H.W.; Cheng, V.C.; et al. Co-circulation of two SARS-CoV-2 variant strains within imported pet hamsters in Hong Kong. Emerg. Microbes. Infect. 2022, 11, 689–698. [Google Scholar] [CrossRef]
- Pickering, B.; Lung, O.; Maguire, F.; Kruczkiewicz, P.; Kotwa, J.D.; Buchanan, T.; Gagnier, M.; Guthrie, J.L.; Jardine, C.M.; Marchand-Austin, A.; et al. Divergent SARS-CoV-2 variant emerges in white-tailed deer with deer-to-human transmission. Nat. Microbiol. 2022, 7, 2011–2024. [Google Scholar] [CrossRef]
- Pagani, G.; Lai, A.; Bergna, A.; Rizzo, A.; Stranieri, A.; Giordano, A.; Paltrinieri, S.; Lelli, D.; Decaro, N.; Rusconi, S.; et al. Human-to-Cat SARS-CoV-2 Transmission: Case Report and Full-Genome Sequencing from an Infected Pet and Its Owner in Northern Italy. Pathogens 2021, 10, 252. [Google Scholar] [CrossRef]
- Hosie, M.J.; Epifano, I.; Herder, V.; Orton, R.J.; Stevenson, A.; Johnson, N.; MacDonald, E.; Dunbar, D.; McDonald, M.; Howie, F.; et al. Detection of SARS-CoV-2 in respiratory samples from cats in the UK associated with human-to-cat transmission. Vet. Rec. 2021, 188, e247. [Google Scholar] [CrossRef] [PubMed]
- Garigliany, M.; Van Laere, A.S.; Clercx, C.; Giet, D.; Escriou, N.; Huon, C.; van der Werf, S.; Eloit, M.; Desmecht, D. SARS-CoV-2 Natural Transmission from Human to Cat, Belgium, March 2020. Emerg. Infect. Dis. 2020, 26, 3069–3071. [Google Scholar] [CrossRef]
- Piewbang, C.; Poonsin, P.; Lohavicharn, P.; Wardhani, S.W.; Dankaona, W.; Puenpa, J.; Poovorawan, Y.; Techangamsuwan, S. SARS-CoV-2 Transmission from Human to Pet and Suspected Transmission from Pet to Human, Thailand. J. Clin. Microbiol. 2022, 60, e0105822. [Google Scholar] [CrossRef] [PubMed]
- Hamer, S.A.; Pauvolid-Correa, A.; Zecca, I.B.; Davila, E.; Auckland, L.D.; Roundy, C.M.; Tang, W.; Torchetti, M.K.; Killian, M.L.; Jenkins-Moore, M.; et al. SARS-CoV-2 Infections and Viral Isolations among Serially Tested Cats and Dogs in Households with Infected Owners in Texas, USA. Viruses 2021, 13, 938. [Google Scholar] [CrossRef] [PubMed]
- Goryoka, G.W.; Cossaboom, C.M.; Gharpure, R.; Dawson, P.; Tansey, C.; Rossow, J.; Mrotz, V.; Rooney, J.; Torchetti, M.; Loiacono, C.M.; et al. One Health Investigation of SARS-CoV-2 Infection and Seropositivity among Pets in Households with Confirmed Human COVID-19 Cases-Utah and Wisconsin 2020. Viruses 2021, 13, 1813. [Google Scholar] [CrossRef]
- Fritz, M.; Rosolen, B.; Krafft, E.; Becquart, P.; Elguero, E.; Vratskikh, O.; Denolly, S.; Boson, B.; Vanhomwegen, J.; Gouilh, M.A.; et al. High prevalence of SARS-CoV-2 antibodies in pets from COVID-19+ households. One Health 2021, 11, 100192. [Google Scholar] [CrossRef]
- Wang, A.; Zhu, X.; Chen, Y.; Sun, Y.; Liu, H.; Ding, P.; Zhou, J.; Liu, Y.; Liang, C.; Yin, J.; et al. Serological survey of SARS-CoV-2 in companion animals in China. Front. Vet. Sci. 2022, 9, 986619. [Google Scholar] [CrossRef]
- Michelitsch, A.; Hoffmann, D.; Wernike, K.; Beer, M. Occurrence of Antibodies against SARS-CoV-2 in the Domestic Cat Population of Germany. Vaccines 2020, 8, 772. [Google Scholar] [CrossRef]
- Stevanovic, V.; Vilibic-Cavlek, T.; Tabain, I.; Benvin, I.; Kovac, S.; Hruskar, Z.; Mauric, M.; Milasincic, L.; Antolasic, L.; Skrinjaric, A.; et al. Seroprevalence of SARS-CoV-2 infection among pet animals in Croatia and potential public health impact. Transbound. Emerg. Dis. 2021, 68, 1767–1773. [Google Scholar] [CrossRef]
- Zhao, S.; Schuurman, N.; Li, W.; Wang, C.; Smit, L.A.M.; Broens, E.M.; Wagenaar, J.A.; van Kuppeveld, F.J.M.; Bosch, B.J.; Egberink, H. Serologic Screening of Severe Acute Respiratory Syndrome Coronavirus 2 Infection in Cats and Dogs during First Coronavirus Disease Wave, the Netherlands. Emerg. Infect. Dis. 2021, 27, 1362–1370. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.I.; Elia, G.; Grassi, A.; Giordano, A.; Desario, C.; Medardo, M.; Smith, S.L.; Anderson, E.R.; Prince, T.; Patterson, G.T.; et al. Evidence of exposure to SARS-CoV-2 in cats and dogs from households in Italy. Nat. Commun. 2020, 11, 6231. [Google Scholar] [CrossRef] [PubMed]
- Kaczorek-Łukowska, E.; Wernike, K.; Beer, M.; Wróbel, M.; Małaczewska, J.; Mikulska-Skupień, E.; Malewska, K.; Mielczarska, I.; Siwicki, A.K. High Seroprevalence against SARS-CoV-2 among Dogs and Cats, Poland, 2021/2022. Animals 2022, 12, 2016. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira-Filho, E.F.; de Carvalho, O.V.; Carneiro, I.O.; Fernandes, F.D.; Vaz, S.N.; Pedroso, C.; Gonzalez-Auza, L.; Urbieta, V.C.; Kühne, A.; Mayoral, R.; et al. Frequent Infection of Cats With SARS-CoV-2 Irrespective of Pre-Existing Enzootic Coronavirus Immunity, Brazil 2020. Front. Immunol. 2022, 13, 857322. [Google Scholar] [CrossRef]
- Jara, L.M.; Ferradas, C.; Schiaffino, F.; Sánchez-Carrión, C.; Martínez-Vela, A.; Ulloa, A.; Isasi-Rivas, G.; Montalván, A.; Sarmiento, L.G.; Fernández, M.; et al. Evidence of neutralizing antibodies against SARS-CoV-2 in domestic cats living with owners with a history of COVID-19 in Lima —Peru. One Health 2021, 13, 100318. [Google Scholar] [CrossRef]
- Bessiere, P.; Vergne, T.; Battini, M.; Brun, J.; Averso, J.; Joly, E.; Guerin, J.L.; Cadiergues, M.C. SARS-CoV-2 Infection in Companion Animals: Prospective Serological Survey and Risk Factor Analysis in France. Viruses 2022, 14, 1178. [Google Scholar] [CrossRef]
- Kannekens-Jager, M.M.; de Rooij, M.M.T.; de Groot, Y.; Biesbroeck, E.; de Jong, M.K.; Pijnacker, T.; Smit, L.A.M.; Schuurman, N.; Broekhuizen-Stins, M.J.; Zhao, S.; et al. SARS-CoV-2 infection in dogs and cats is associated with contact to COVID-19-positive household members. Transbound Emerg. Dis. 2022, 69, 4034–4040. [Google Scholar] [CrossRef]
- Barroso, R.; Vieira-Pires, A.; Antunes, A.; Fidalgo-Carvalho, I. Susceptibility of Pets to SARS-CoV-2 Infection: Lessons from a Seroepidemiologic Survey of Cats and Dogs in Portugal. Microorganisms 2022, 10, 345. [Google Scholar] [CrossRef]
- Michelitsch, A.; Schön, J.; Hoffmann, D.; Beer, M.; Wernike, K. The Second Wave of SARS-CoV-2 Circulation-Antibody Detection in the Domestic Cat Population in Germany. Viruses 2021, 13, 1009. [Google Scholar] [CrossRef]
- Smith, S.L.; Anderson, E.R.; Cansado-Utrilla, C.; Prince, T.; Farrell, S.; Brant, B.; Smyth, S.; Noble, P.M.; Pinchbeck, G.L.; Marshall, N.; et al. SARS-CoV-2 neutralising antibodies in dogs and cats in the United Kingdom. Curr. Res. Virol. Sci. 2021, 2, 100011. [Google Scholar] [CrossRef]
- Census and Statistics Department. Thematic Household Survey Report No. 66—Keeping of Dogs and Cats; Census and Statistics Department: Hong Kong SAR, China, 2019.
- Ministry of Agriculture, Food and Rural Affairs. 8.60 Million Companion Animals are Raised in 6.38 Million Households Nationwide 2021. In Korean. Available online: https://www.mafra.go.kr/mafra/293/subview.do?enc=Zm5jdDF8QEB8JTJGYmJzJTJGbWFmcmElMkY2OCUyRjMyNjk3MCUyRmFydGNsVmlldy5kbyUzRg%3D%3D (accessed on 1 November 2022).
- Perera, R.A.; Mok, C.K.; Tsang, O.T.; Lv, H.; Ko, R.L.; Wu, N.C.; Perera, R.A.; Yuan, M.; Leung, W.S.; Chan, J.M.; et al. Serological assays for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), March 2020. Euro. Surveill. 2020, 25, 2000421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshpande, S.S. Enzyme Immunoassays: From Concept to Product Development; Chapman & Hall: New York, NY, USA, 1996. [Google Scholar]
- Donnik, I.M.; Popov, I.V.; Sereda, S.V.; Popov, I.V.; Chikindas, M.L.; Ermakov, A.M. Coronavirus Infections of Animals: Future Risks to Humans. Biol. Bull. Russ. Acad. Sci. 2021, 48, 26–37. [Google Scholar] [CrossRef]
- The Government of the Hong Kong Special Administrative Region. Local Situation Dashboard. Available online: https://www.coronavirus.gov.hk/eng/index.html (accessed on 1 November 2022).
- Schulz, C.; Martina, B.; Mirolo, M.; Muller, E.; Klein, R.; Volk, H.; Egberink, H.; Gonzalez-Hernandez, M.; Kaiser, F.; von Kockritz-Blickwede, M.; et al. SARS-CoV-2-Specific Antibodies in Domestic Cats during First COVID-19 Wave, Europe. Emerg. Infect. Dis. 2021, 27, 3115–3118. [Google Scholar] [CrossRef]
- Klaus, J.; Meli, M.L.; Willi, B.; Nadeau, S.; Beisel, C.; Stadler, T.; Eth Sars-Co, V.S.T.; Egberink, H.; Zhao, S.; Lutz, H.; et al. Detection and Genome Sequencing of SARS-CoV-2 in a Domestic Cat with Respiratory Signs in Switzerland. Viruses 2021, 13, 496. [Google Scholar] [CrossRef]
- Perera, R.; Ko, R.; Tsang, O.T.Y.; Hui, D.S.C.; Kwan, M.Y.M.; Brackman, C.J.; To, E.M.W.; Yen, H.L.; Leung, K.; Cheng, S.M.S.; et al. Evaluation of a SARS-CoV-2 Surrogate Virus Neutralization Test for Detection of Antibody in Human, Canine, Cat, and Hamster Sera. J. Clin. Microbiol. 2021, 59, e02504-20. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.; Tiu, C.; Hu, Z.; Chen, V.C.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2-spike protein-protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Hillyer, C.; Du, L. Neutralizing Antibodies against SARS-CoV-2 and Other Human Coronaviruses. Trends Immunol. 2020, 41, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Dileepan, M.; Di, D.; Huang, Q.; Ahmed, S.; Heinrich, D.; Ly, H.; Liang, Y. Seroprevalence of SARS-CoV-2 (COVID-19) exposure in pet cats and dogs in Minnesota, USA. Virulence 2021, 12, 1597–1609. [Google Scholar] [CrossRef] [PubMed]
- Felten, S.; Klein-Richers, U.; Hofmann-Lehmann, R.; Bergmann, M.; Unterer, S.; Leutenegger, C.M.; Hartmann, K. Correlation of Feline Coronavirus Shedding in Feces with Coronavirus Antibody Titer. Pathogens 2020, 9, 598. [Google Scholar] [CrossRef]
- Klaus, J.; Zini, E.; Hartmann, K.; Egberink, H.; Kipar, A.; Bergmann, M.; Palizzotto, C.; Zhao, S.; Rossi, F.; Franco, V.; et al. SARS-CoV-2 Infection in Dogs and Cats from Southern Germany and Northern Italy during the First Wave of the COVID-19 Pandemic. Viruses 2021, 13, 1453. [Google Scholar] [CrossRef]
- Ito, G.; Goto-Koshino, Y.; Kuroda, Y.; Eunsil, P.; Maeda, K.; Soma, T.; Momoi, Y. Seroprevalence of antibodies against severe acute respiratory coronavirus 2 (SARS-CoV-2) in household dogs in Japan. J. Vet. Med. Sci. 2021, 83, 1722–1725. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Total Sera | Origin | Collection Date | No. of Sera | No. of ELISA Positive (%) | No. of sVNT Positive (%) | No. of PRNT Positive (%) |
|---|---|---|---|---|---|---|---|
| Cats | 1040 | Korea | 04/2020 to 07/2020 | 488 | 22 (4.51%) | 8 (1.64%) | 2 (0.41%) |
| Hong Kong | 552 | 14 (2.54%) | 1 (0.18%) | 0 (0%) | |||
| -Veterinary clinic | 03/2020 to 04/2021 | 251 | 13 | 1 | 0 | ||
| -Animal shelter | 01/2021 to 05/2021 | 301 | 1 | 0 | 0 |
| Species | Total Sera | Origin | Collection Date | No. of Sera | No. of ELISA Positive (%) | No. of sVNT Positive (%) | No. of PRNT Positive (%) |
|---|---|---|---|---|---|---|---|
| Dogs | 855 | Korea | 04/20–05/20 | 344 | 17 (4.94%) | 1 (0.29%) | 0 |
| Hong Kong | 01/21–08/21 | 511 | 33 (6.46%) | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Go, Y.Y.; Carrai, M.; Choi, Y.R.; Brackman, C.J.; Tam, K.W.S.; Law, P.Y.T.; Woodhouse, F.; Gray, J.; Kim, J.H.; Park, J.; et al. Low Prevalence of SARS-CoV-2 Antibodies in Canine and Feline Serum Samples Collected during the COVID-19 Pandemic in Hong Kong and Korea. Viruses 2023, 15, 582. https://doi.org/10.3390/v15020582
Go YY, Carrai M, Choi YR, Brackman CJ, Tam KWS, Law PYT, Woodhouse F, Gray J, Kim JH, Park J, et al. Low Prevalence of SARS-CoV-2 Antibodies in Canine and Feline Serum Samples Collected during the COVID-19 Pandemic in Hong Kong and Korea. Viruses. 2023; 15(2):582. https://doi.org/10.3390/v15020582
Chicago/Turabian StyleGo, Yun Young, Maura Carrai, Yan Ru Choi, Christopher J. Brackman, Karina W. S. Tam, Pierra Y. T. Law, Fiona Woodhouse, Jane Gray, Ji Hun Kim, Joohyung Park, and et al. 2023. "Low Prevalence of SARS-CoV-2 Antibodies in Canine and Feline Serum Samples Collected during the COVID-19 Pandemic in Hong Kong and Korea" Viruses 15, no. 2: 582. https://doi.org/10.3390/v15020582
APA StyleGo, Y. Y., Carrai, M., Choi, Y. R., Brackman, C. J., Tam, K. W. S., Law, P. Y. T., Woodhouse, F., Gray, J., Kim, J. H., Park, J., Jeon, C. W., Jang, H., Magouras, I., Decaro, N., Cheng, S. M. S., Peiris, M., Beatty, J. A., & Barrs, V. R. (2023). Low Prevalence of SARS-CoV-2 Antibodies in Canine and Feline Serum Samples Collected during the COVID-19 Pandemic in Hong Kong and Korea. Viruses, 15(2), 582. https://doi.org/10.3390/v15020582