
Alterations in Abundance and Compartmentalization of miRNAs in Blood Plasma Extracellular Vesicles and Extracellular Condensates during HIV/SIV Infection and Its Modulation by Antiretroviral Therapy (ART) and Delta-9-Tetrahydrocannabinol (Δ9-THC)

 , and
, and 
Abstract
:1. Introduction
2. Results
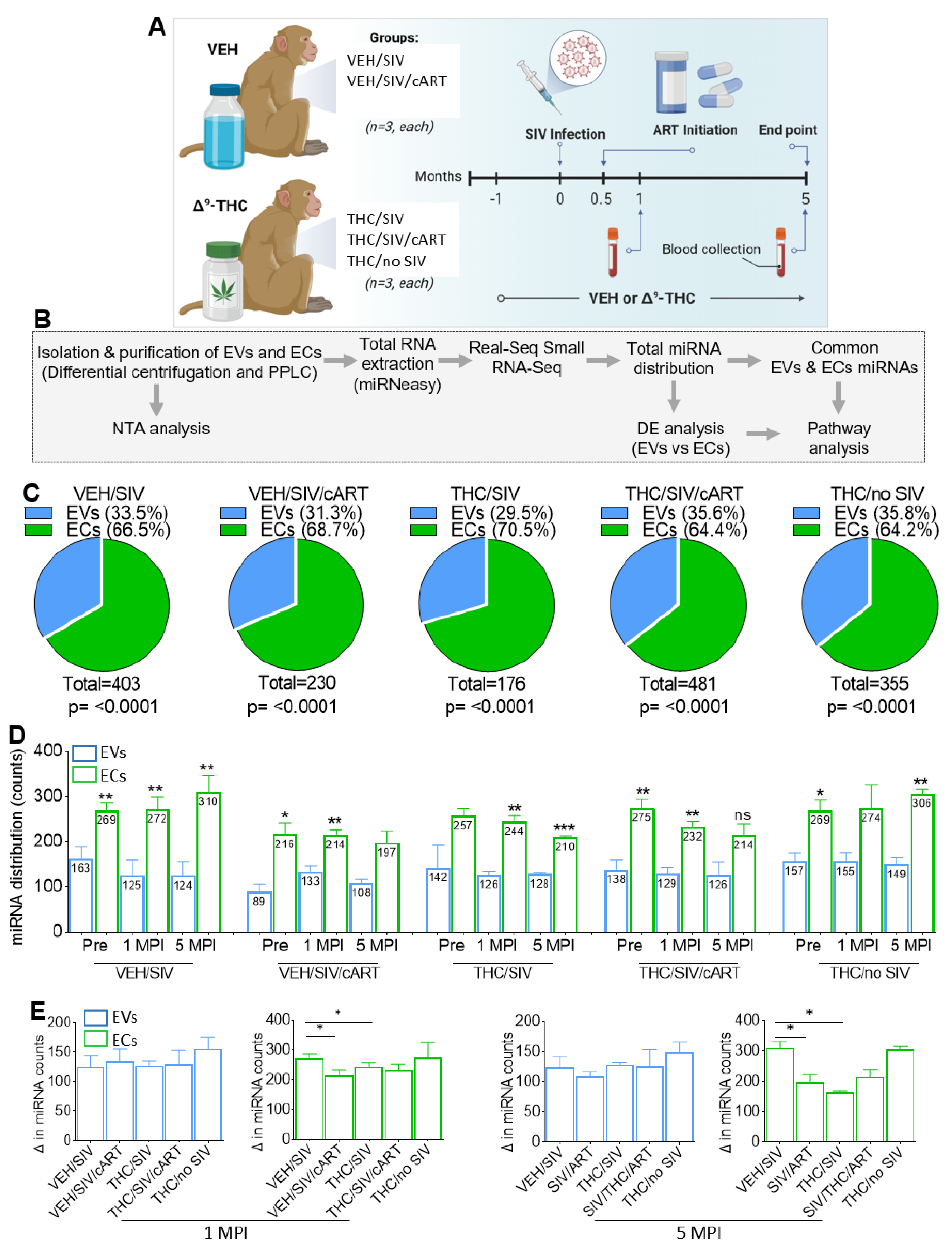
2.1. The Isolation Profiles of EDTA Blood Plasma exmiRNA Carriers (EVs and ECs) Are Different but Not Changed by SIV, THC, cART, or Both THC and cART Administered Together
2.2. Circulating miRNA Repertoire and Their Association with EVs and ECs following SIV Infection and Treatment
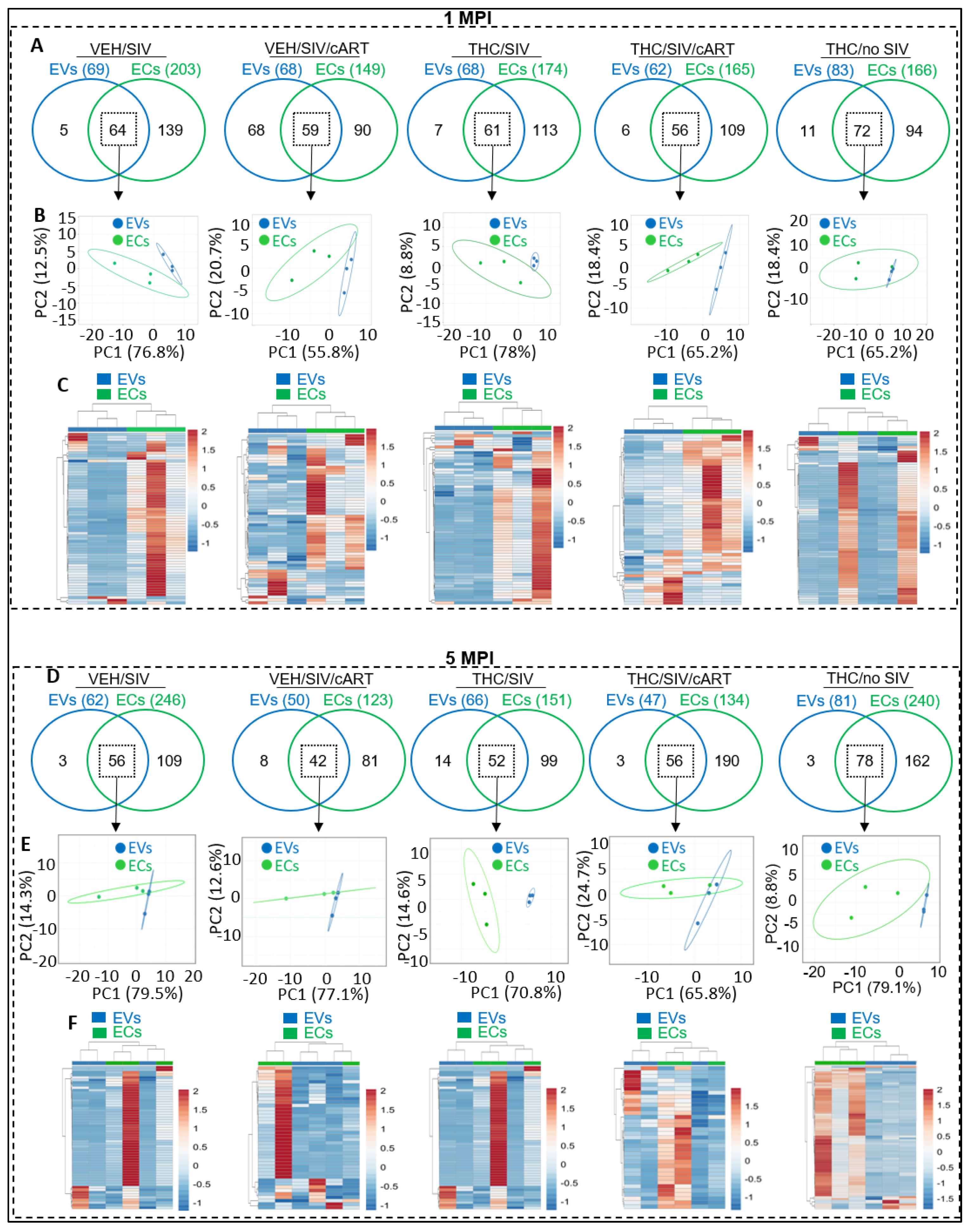
2.3. Differences and Similarities in EV and EC Associated miRNA Repertoires following SIV Infection and Treatment with cART and THC
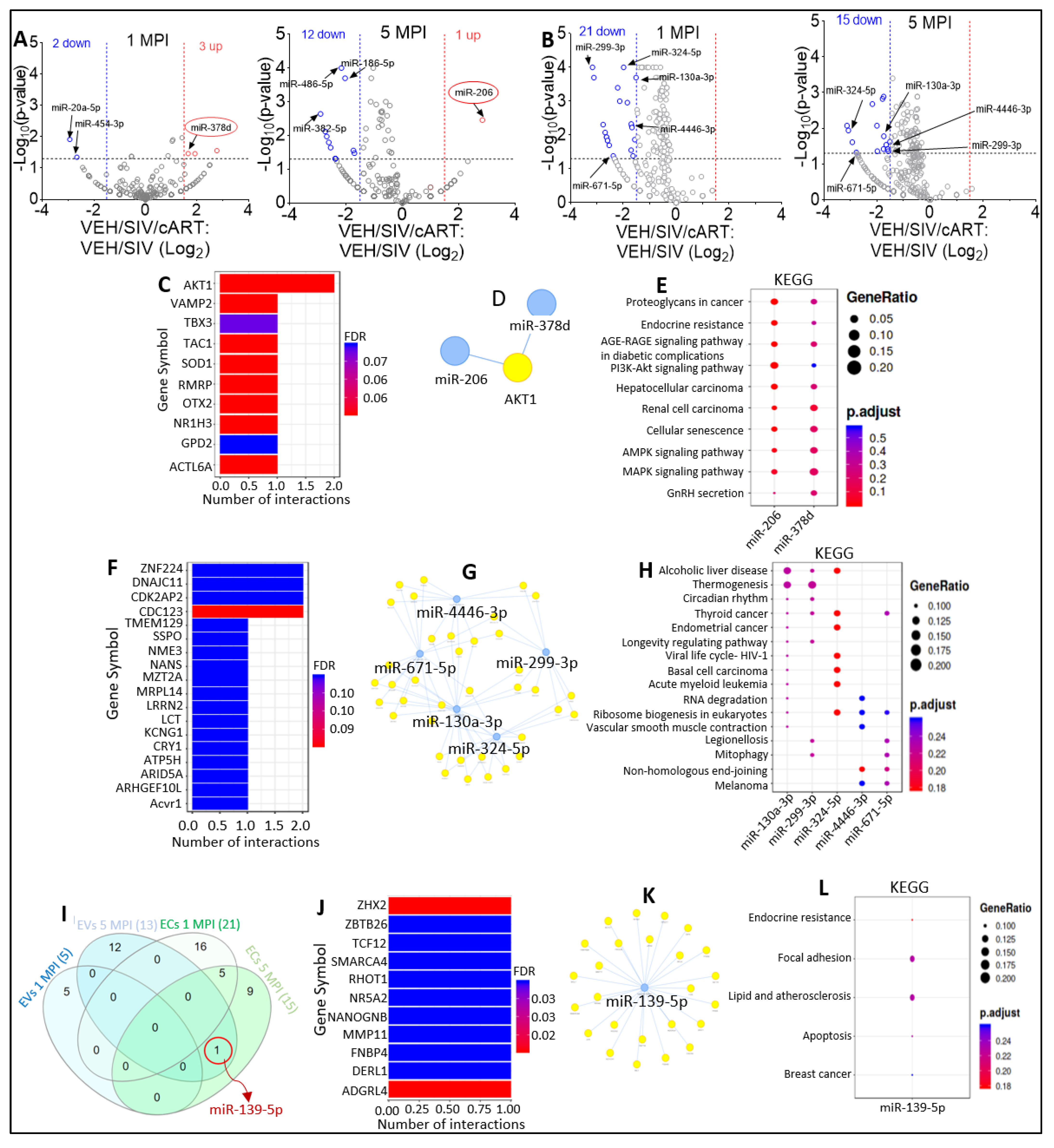
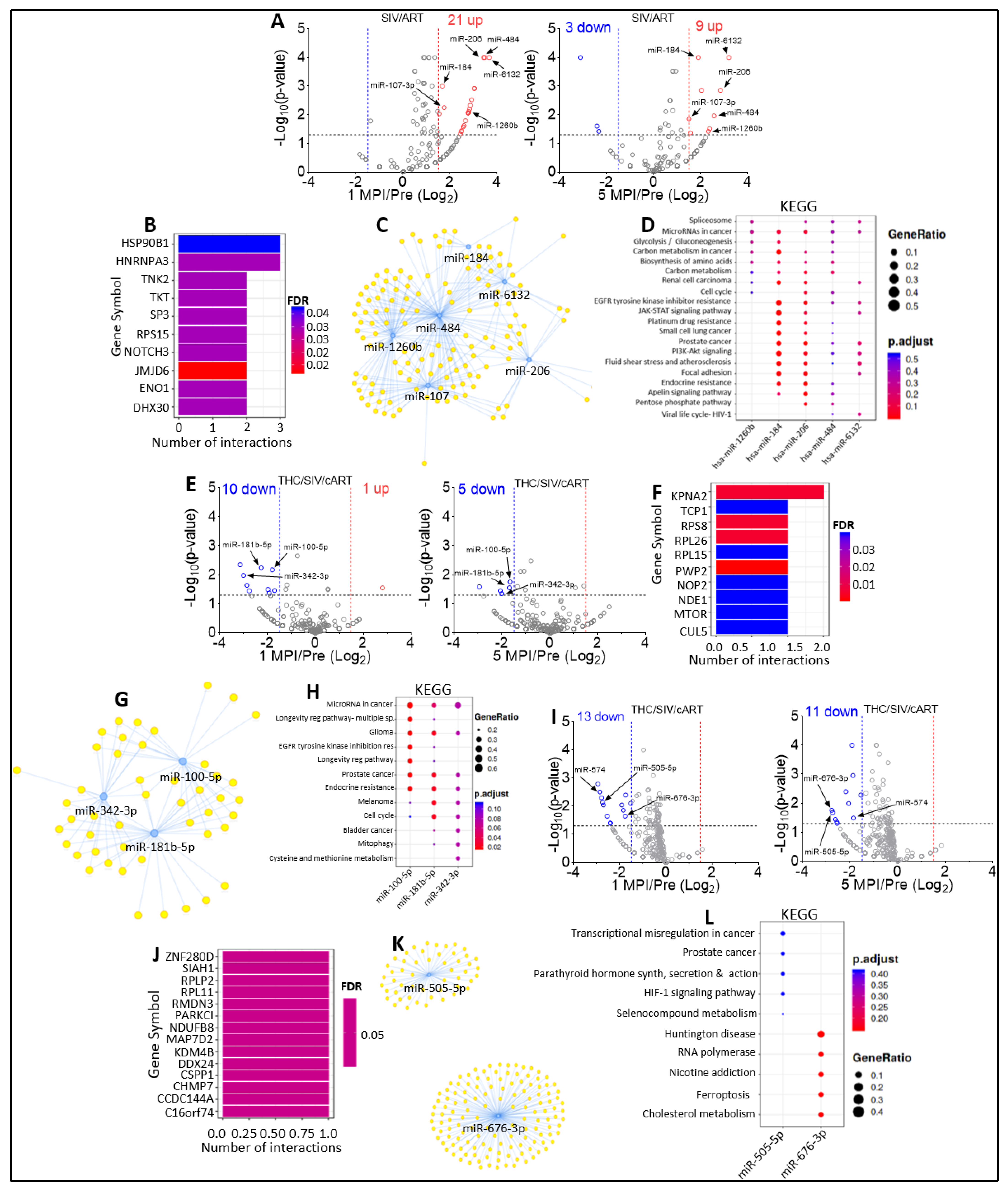
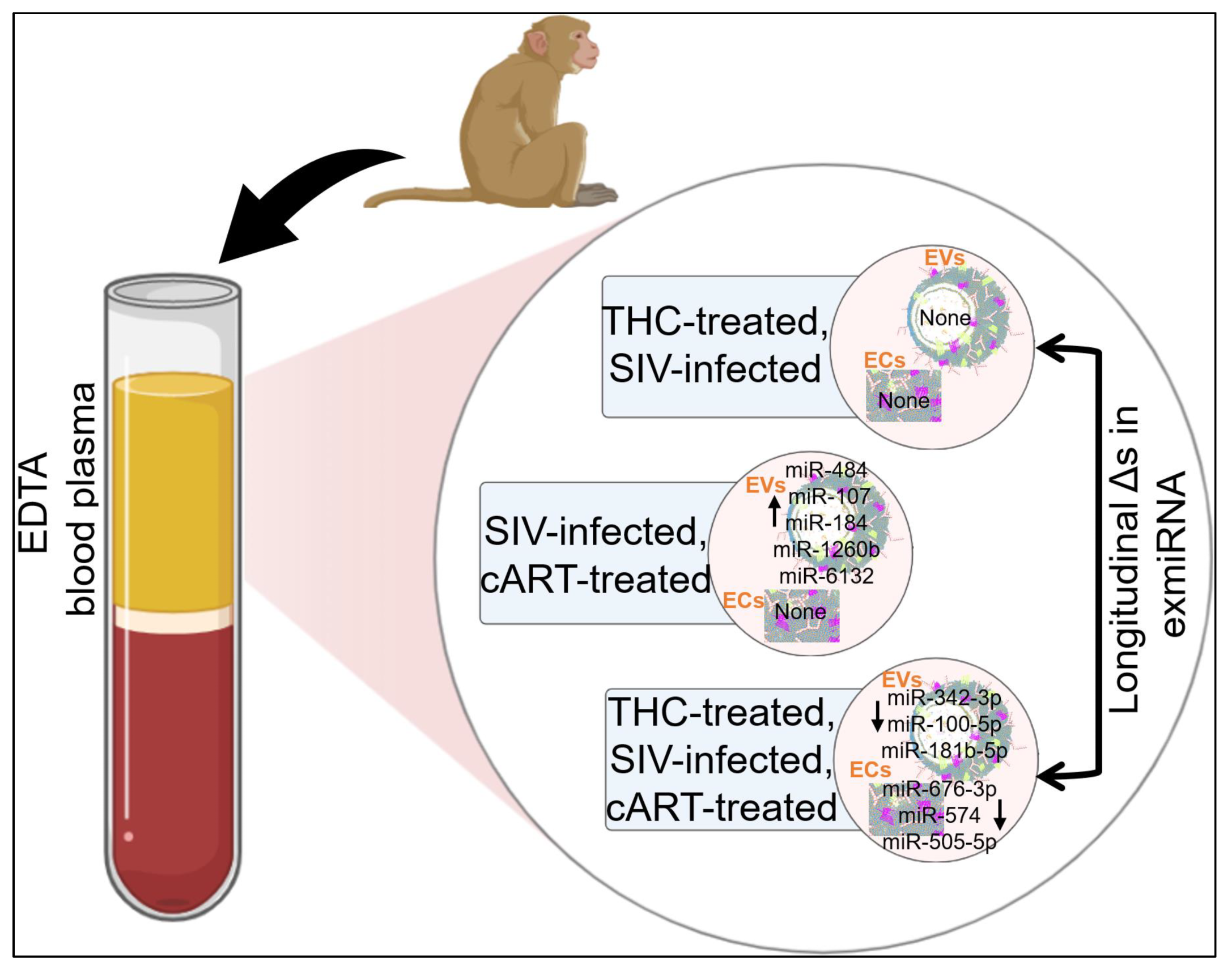
2.4. Varying miRNA Abundance in EVs and ECs in Response to SIV Infection and Viral Suppression by ART
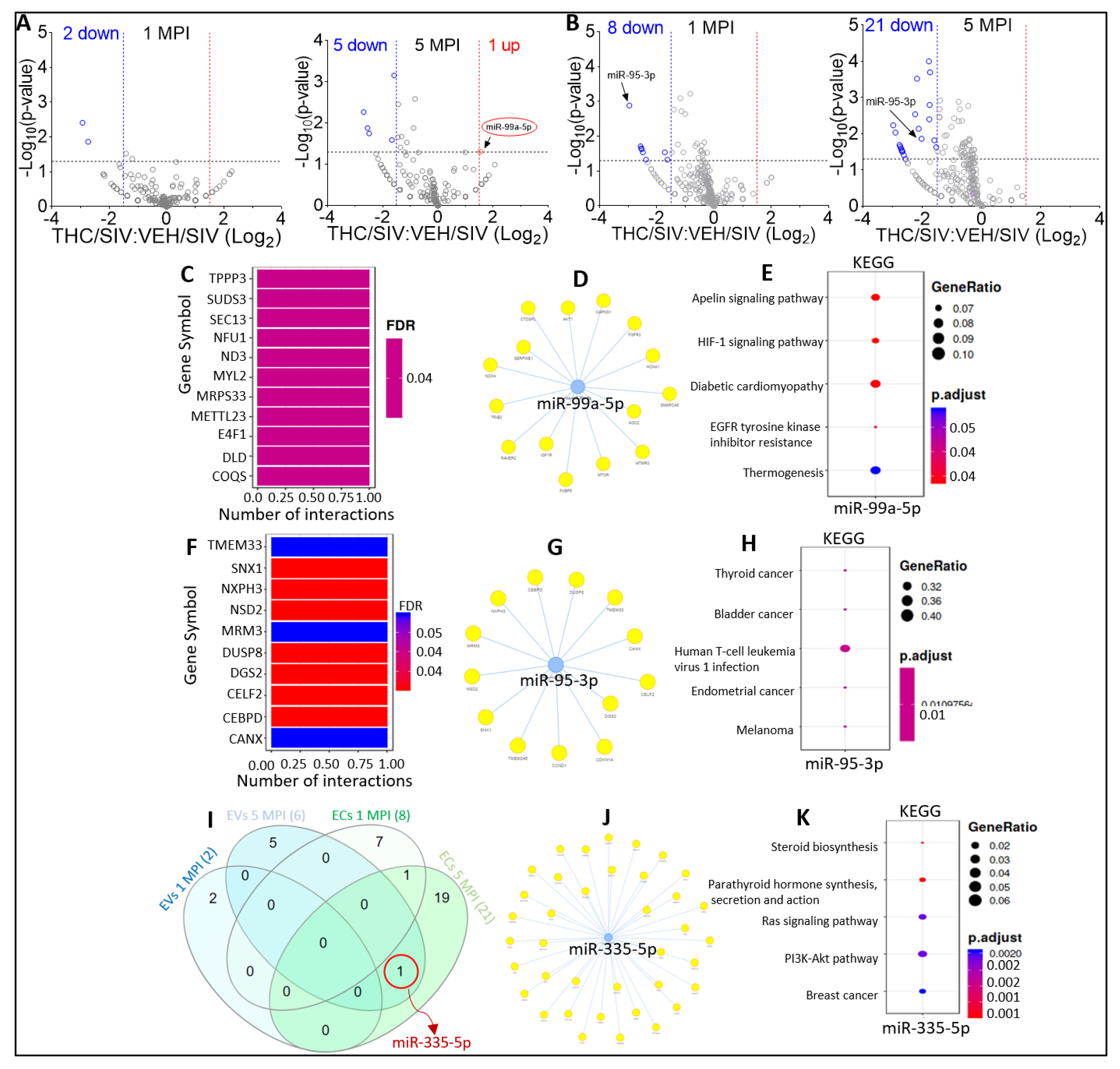
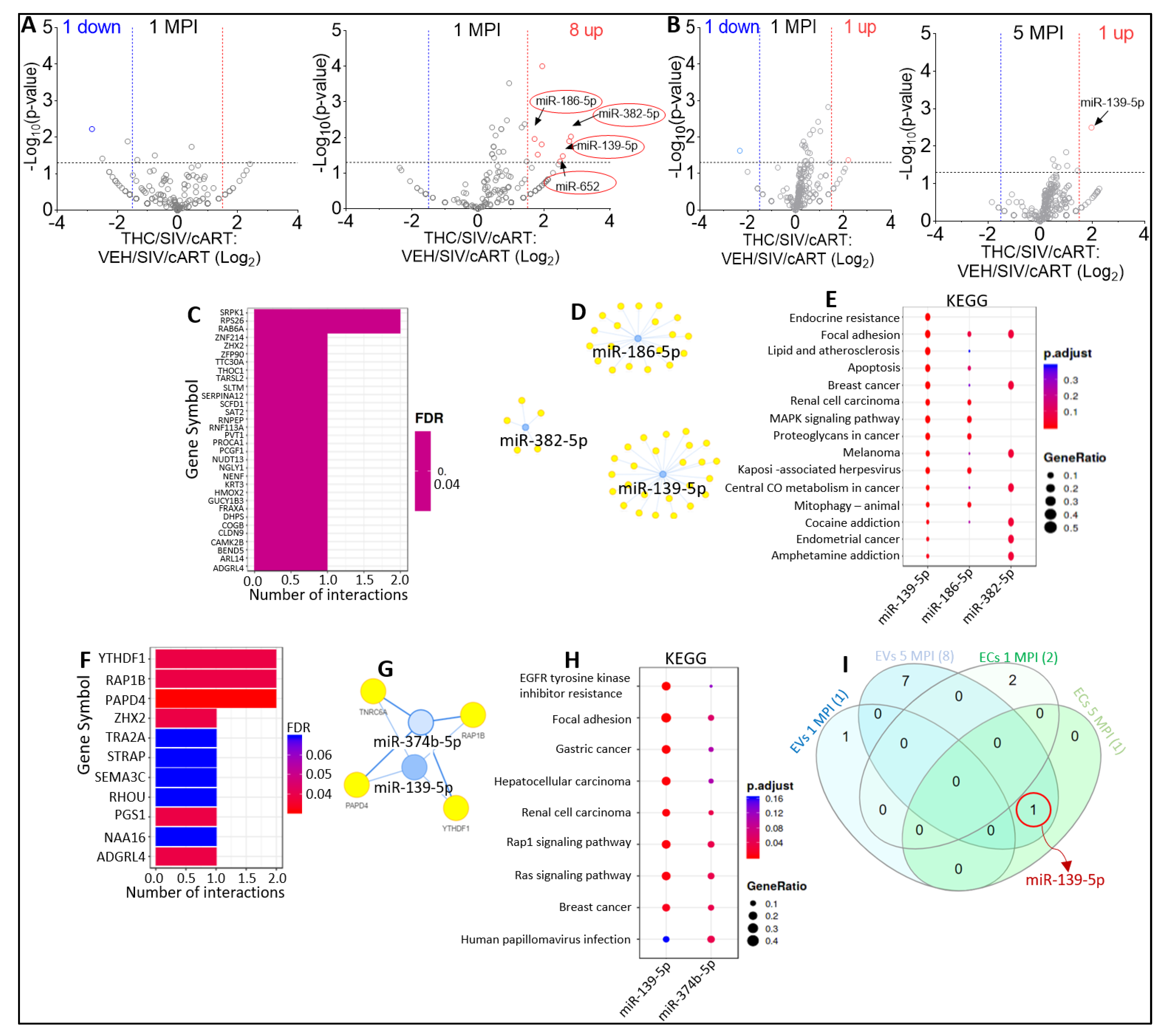
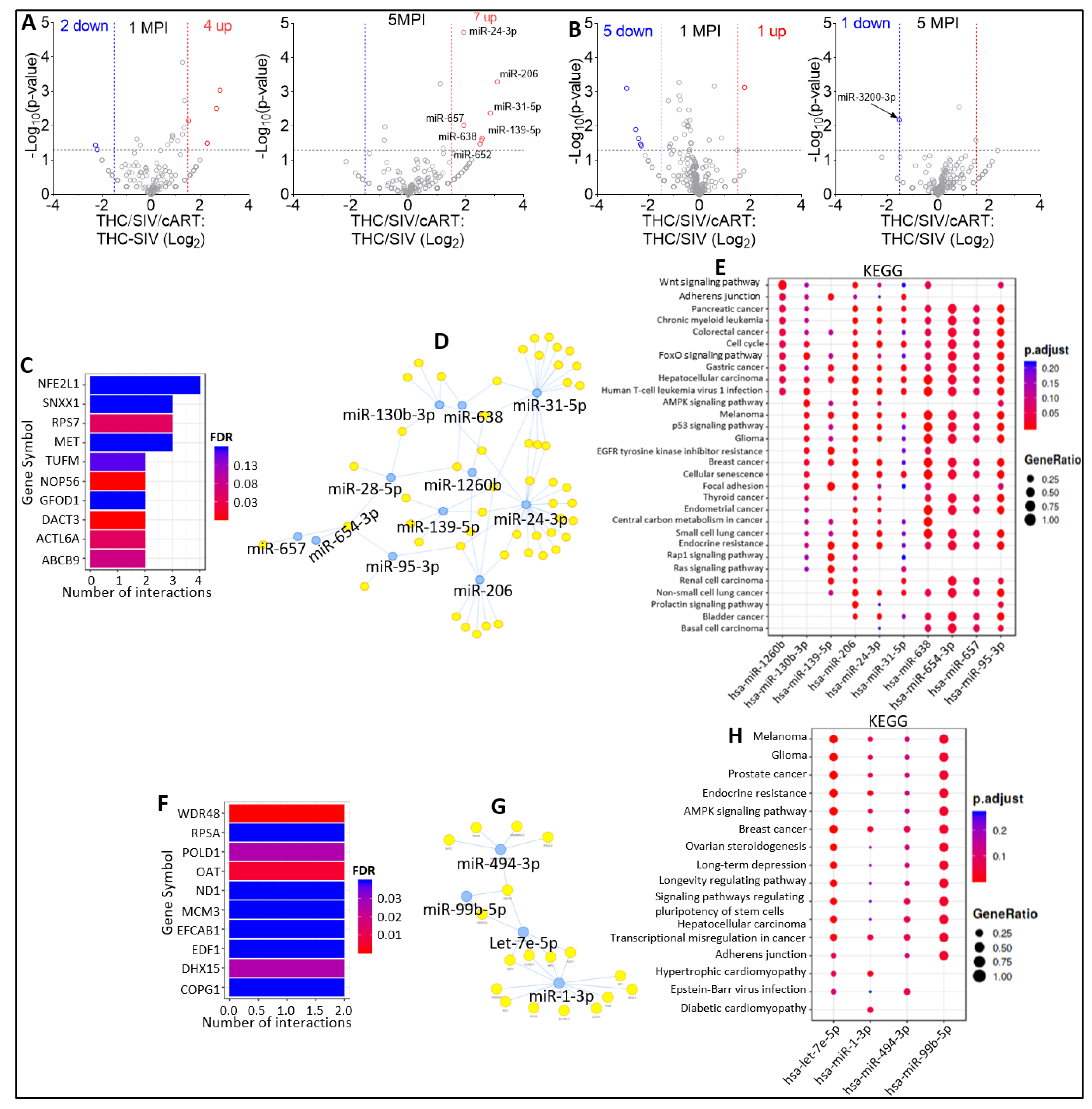
2.5. Effect of THC on miRNA Profiles following SIV Infection in the Absence of Viral Suppression by cART
2.6. Effect of THC on miRNA Profile of SIV-infected Rhesus Macaques under Suppressive cART
2.7. Effect of cART on miRNA Profile of SIV-infected Rhesus Macaque in the Presence of Anti-Inflammatory THC
2.8. Defining Clinically Relevant miRNAs through Bioinformatics Integrative Analysis of Longitudinal Changes in the miRNAome of RM EVs and ECs Induced by SIV, cART and/or THC
3. Discussion
4. Materials and Methods
4.1. Macaques and Viruses (used for this Study and the Primary Study Presented in Manuscript 1)
4.2. Isolation of EVs and ECs (used for this Study and the Primary Study in Manuscript 1)
4.3. Nanoparticle Tracking Analysis (NTA) (used for this Study and the Primary Study in Manuscript 1)
4.4. Total RNA Isolation(used for this Study and the Primary Study in Manuscript 1)
4.5. Library Preparation and sRNA Sequencing (used for this Study and the Primary Study in Manuscript 1)
4.6. Identification of Common miRNAs (used for this Study and the Primary Study in Manuscript 1)
4.7. PCA Plot and Heatmap Generation (used for this Study and the Primary Study in Manuscript 1)
4.8. Identification of Differentially Enriched miRNAs (used for this Study and the Primary Study in Manuscript 1)
4.9. miRNA-Target Enrichment Analysis (used for this Study and the Primary Study in Manuscript 1)
4.10. Statistical Analyses (used for this Study and the Primary Study in Manuscript 1)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ECs | Extracellular condensates |
| EVs | Extracellular vesicles |
| ART | Antiretroviral therapy |
| THC | Delta-9-tetrahydrocannabinol |
| VEH | Vehicle |
References
- Moore, R.D. Epidemiology of HIV Infection in the United States: Implications for Linkage to Care. Clin. Infect. Dis. 2011, 52, S208–S213. [Google Scholar] [CrossRef] [Green Version]
- Hemelaar, J.; Elangovan, R.; Yun, J.; Dickson-Tetteh, L.; Fleminger, I.; Kirtley, S.; Williams, B.; Gouws-Williams, E.; Ghys, P.D.; Characterisation, W.-U.N.f.H.I. Global and regional molecular epidemiology of HIV-1, 1990-2015: A systematic review, global survey, and trend analysis. Lancet Infect Dis. 2019, 19, 143–155. [Google Scholar] [CrossRef]
- Vijayan, K.K.V.; Karthigeyan, K.P.; Tripathi, S.P.; Hanna, L.E. Pathophysiology of CD4+T-Cell Depletion in HIV-1 and HIV-2 infections. Front. Immunol. 2017, 8, 580. [Google Scholar] [CrossRef] [Green Version]
- Deeks, S.G.; Lewin, S.R.; Havlir, D.V. The end of AIDS: HIV infection as a chronic disease. Lancet 2013, 382, 1525–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatano, H. Immune activation and HIV persistence: Considerations for novel therapeutic interventions. Curr. Opin. Hiv Aids 2013, 8, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Zicari, S.; Sessa, L.; Cotugno, N.; Ruggiero, A.; Morrocchi, E.; Concato, C.; Rocca, S.; Zangari, P.; Manno, E.C.; Palma, P. Immune Activation, Inflammation, and Non-AIDS Co-Morbidities in HIV-Infected Patients under Long-Term ART. Viruses-Basel 2019, 11, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrams, D.I.; Jay, C.A.; Shade, S.B.; Vizoso, H.; Reda, H.; Press, S.; Kelly, M.E.; Rowbotham, M.C.; Petersen, K.L. Cannabis in painful HIV-associated sensory neuropathy - A randomized placebo-controlled trial. Neurology 2007, 68, 515–521. [Google Scholar] [CrossRef]
- Abrams, D.I.; Hilton, J.F.; Leiser, R.J.; Shade, S.B.; Elbeik, T.A.; Aweeka, F.T.; Benowitz, N.L.; Bredt, B.M.; Kosel, B.; Aberg, J.A.; et al. Short-term effects of cannabinoids in patients with HIV-1 infection - A randomized, placebo-controlled clinical trial. Ann. Intern. Med. 2003, 139, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Wilson, N.L.; Peterson, S.N.; Ellis, R.J. Cannabis and the Gut-Brain Axis Communication in HIV Infection. Cannabis Cannabinoid Res. 2021, 6, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Sereti, I.; Altfeld, M. Immune activation and HIV: An enduring relationship. Curr. Opin. Hiv Aids 2016, 11, 129. [Google Scholar] [CrossRef] [Green Version]
- Haney, M.; Rabkin, J.; Gunderson, E.; Foltin, R.W. Dronabinol and marijuana in HIV(+) marijuana smokers: Acute effects on caloric intake and mood. Psychopharmacology 2005, 181, 170–178. [Google Scholar] [CrossRef]
- Riggs, P.K.; Vaida, F.; Rossi, S.S.; Sorkin, L.S.; Gouaux, B.; Grant, I.; Ellis, R.J. A pilot study of the effects of cannabis on appetite hormones in HIV-infected adult men. Brain Res. 2012, 1431, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Wilson, N.L.; Azuero, A.; Vance, D.E.; Richman, J.S.; Moneyham, L.D.; Raper, J.L.; Heath, S.L.; Kempf, M.C. Identifying Symptom Patterns in People Living With HIV Disease. Janac-J. Assoc. Nurses Aids Care 2016, 27, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Brooks, E.; Gundersen, D.C.; Flynn, E.; Brooks-Russell, A.; Bull, S. The clinical implications of legalizing marijuana: Are physician and non-physician providers prepared? Addict. Behav. 2017, 72, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Milloy, M.J.; Marshall, B.; Kerr, T.; Richardson, L.; Hogg, R.; Guillemi, S.; Montaner, J.S.; Wood, E. High-intensity cannabis use associated with lower plasma human immunodeficiency virus-1 RNA viral load among recently infected people who use injection drugs. Drug Alcohol Rev. 2015, 34, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, M.D.; Crawford, R.B.; Henriquez, J.E.; Aldhamen, Y.A.; Gulick, P.; Amalfitano, A.; Kaminski, N.E. HIV-infected cannabis users have lower circulating CD16+ monocytes and IFN-gamma-inducible protein 10 levels compared with nonusing HIV patients. Aids 2018, 32, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Zurier, R.B.; Burstein, S.H. Cannabinoids, inflammation, and fibrosis. Faseb J. 2016, 30, 3682–3689. [Google Scholar] [CrossRef] [Green Version]
- Tambaro, S.; Casu, M.A.; Mastinu, A.; Lazzari, P. Evaluation of selective cannabinoid CB(1) and CB(2) receptor agonists in a mouse model of lipopolysaccharide-induced interstitial cystitis. Eur. J. Pharmacol. 2014, 729, 67–74. [Google Scholar] [CrossRef]
- Castro, F.O.F.; Silva, J.M.; Dorneles, G.P.; Barros, J.B.S.; Ribeiro, C.B.; Noronha, I.; Barbosa, G.R.; Souza, L.C.S.; Guilarde, A.O.; Pereira, A.; et al. Distinct inflammatory profiles in HIV-infected individuals under antiretroviral therapy using cannabis, cocaine or cannabis plus cocaine. AIDS 2019, 33, 1831–1842. [Google Scholar] [CrossRef]
- Galaj, E.; Bi, G.H.; Yang, H.J.; Xi, Z.X. Cannabidiol attenuates the rewarding effects of cocaine in rats by CB2, 5-HT(1A) and TRPV1 receptor mechanisms. Neuropharmacology 2020, 167, 107740. [Google Scholar] [CrossRef]
- Molina, P.E.; Winsauer, P.; Zhang, P.; Walker, E.; Birke, L.; Amedee, A.; Stouwe, C.V.; Troxclair, D.; McGoey, R.; Varner, K.; et al. Cannabinoid Administration Attenuates the Progression of Simian Immunodeficiency Virus. Aids Res. Hum. Retrovir. 2011, 27, 585-U122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winsauer, P.J.; Molina, P.E.; Amedee, A.M.; Filipeanu, C.M.; McGoey, R.R.; Troxclair, D.A.; Walker, E.M.; Birke, L.L.; Stouwe, C.V.; Howard, J.M.; et al. Tolerance to Chronic Delta-9-Tetrahydrocannabinol (Delta(9)-THC) in Rhesus Macaques Infected With Simian Immunodeficiency Virus. Exp. Clin. Psychopharmacol. 2011, 19, 154–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noorbakhsh, F.; Ramachandran, R.; Barsby, N.; Ellestad, K.K.; LeBlanc, A.; Dickie, P.; Baker, G.; Hollenberg, M.D.; Cohen, E.A.; Power, C. MicroRNA profiling reveals new aspects of HIV neurodegeneration: Caspase-6 regulates astrocyte survival. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2010, 24, 1799–1812. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W.; Watson, A.K.; Blankson, J.N.; Clements, J.E. Relationships of PBMC microRNA expression, plasma viral load, and CD4+ T-cell count in HIV-1-infected elite suppressors and viremic patients. Retrovirology 2012, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct Target Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinniah, R.; Adimulam, T.; Nandlal, L.; Arumugam, T.; Ramsuran, V. The Effect of miRNA Gene Regulation on HIV Disease. Front. Genet. 2022, 13, 862642. [Google Scholar] [CrossRef]
- Bali, P.; Kenny, P.J. MicroRNAs and Drug Addiction. Front. Genet. 2013, 4, 43. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.W.; Mendell, J.T. MicroRNAs in cell proliferation, cell death, and tumorigenesis. Br. J. Cancer 2007, 96, R40–R44. [Google Scholar] [CrossRef]
- Kaddour, H.; Lyu, Y.; Shouman, N.; Mohan, M.; Okeoma, C.M. Development of Novel High-Resolution Size-Guided Turbidimetry-Enabled Particle Purification Liquid Chromatography (PPLC): Extracellular Vesicles and Membraneless Condensates in Focus. Int. J. Mol. Sci. 2020, 21, 5361. [Google Scholar] [CrossRef]
- Kaddour, H.; Kopcho, S.; Lyu, Y.; Shouman, N.; Paromov, V.; Pratap, S.; Dash, C.; Kim, E.-Y.; Martinson, J.; McKay, H.; et al. HIV-infection and cocaine use regulate semen extracellular vesicles proteome and miRNAome in a manner that mediates strategic monocyte haptotaxis governed by miR-128 network. Cell. Mol. Life Sci. 2021, 79, 5. [Google Scholar] [CrossRef]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Skog, J.; Würdinger, T.; van Rijn, S.; Meijer, D.H.; Gainche, L.; Sena-Esteves, M.; Curry, W.T., Jr.; Carter, B.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [Green Version]
- Simon, L.; Ford, S.M., Jr.; Song, K.; Berner, P.; Vande Stouwe, C.; Nelson, S.; Bagby, G.J.; Molina, P.E. Decreased myoblast differentiation in chronic binge alcohol-administered simian immunodeficiency virus-infected male macaques: Role of decreased miR-206. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R240–R250. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Abbas, W.; Colin, L.; Khan, K.A.; Bouchat, S.; Varin, A.; Larbi, A.; Gatot, J.S.; Kabeya, K.; Vanhulle, C.; et al. Tuning of AKT-pathway by Nef and its blockade by protease inhibitors results in limited recovery in latently HIV infected T-cell line. Sci. Rep. 2016, 6, 24090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquereau, S.; Kumar, A.; Abbas, W.; Herbein, G. Counteracting Akt Activation by HIV Protease Inhibitors in Monocytes/Macrophages. Viruses 2018, 10, 190. [Google Scholar] [CrossRef] [Green Version]
- Shenoy, A.; Danial, M.; Blelloch, R.H. Let-7 and miR-125 cooperate to prime progenitors for astrogliogenesis. EMBO J. 2015, 34, 1180–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Han, L.; Chen, F. Let-7a-5p regulates the inflammatory response in chronic rhinosinusitis with nasal polyps. Diagn. Pathol. 2021, 16, 27. [Google Scholar] [CrossRef]
- Yuan, H.; Zhang, H.; Hong, L.; Zhao, H.; Wang, J.; Li, H.; Che, H.; Zhang, Z. MicroRNA let-7c-5p Suppressed Lipopolysaccharide-Induced Dental Pulp Inflammation by Inhibiting Dentin Matrix Protein-1-Mediated Nuclear Factor kappa B (NF-κB) Pathway In Vitro and In Vivo. Med. Sci. Monit. 2018, 24, 6656–6665. [Google Scholar] [CrossRef]
- Lv, J.; Zeng, Y.; Qian, Y.; Dong, J.; Zhang, Z.; Zhang, J. MicroRNA let-7c-5p improves neurological outcomes in a murine model of traumatic brain injury by suppressing neuroinflammation and regulating microglial activation. Brain Res. 2018, 1685, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, D.L.; Zuluaga-Ramirez, V.; Gajghate, S.; Reichenbach, N.L.; Polyak, B.; Persidsky, Y.; Rom, S. miR-98 reduces endothelial dysfunction by protecting blood-brain barrier (BBB) and improves neurological outcomes in mouse ischemia/reperfusion stroke model. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2020, 40, 1953–1965. [Google Scholar] [CrossRef]
- Rom, S.; Dykstra, H.; Zuluaga-Ramirez, V.; Reichenbach, N.L.; Persidsky, Y. miR-98 and let-7g* protect the blood-brain barrier under neuroinflammatory conditions. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2015, 35, 1957–1965. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, D.L.; Gajghate, S.; Reichenbach, N.L.; Winfield, M.; Persidsky, Y.; Heldt, N.A.; Rom, S. let-7g counteracts endothelial dysfunction and ameliorating neurological functions in mouse ischemia/reperfusion stroke model. Brain Behav. Immun. 2020, 87, 543–555. [Google Scholar] [CrossRef]
- Swaminathan, S.; Suzuki, K.; Seddiki, N.; Kaplan, W.; Cowley, M.J.; Hood, C.L.; Clancy, J.L.; Murray, D.D.; Méndez, C.; Gelgor, L.; et al. Differential regulation of the Let-7 family of microRNAs in CD4+ T cells alters IL-10 expression. J. Immunol. 2012, 188, 6238–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yin, Y.; Zhang, S.; Luo, H.; Zhang, H. HIV-1 Infection-Induced Suppression of the Let-7i/IL-2 Axis Contributes to CD4(+) T Cell Death. Sci. Rep. 2016, 6, 25341. [Google Scholar] [CrossRef] [Green Version]
- Ayuso, C.; Rimola, J.; Vilana, R.; Burrel, M.; Darnell, A.; García-Criado, Á.; Bianchi, L.; Belmonte, E.; Caparroz, C.; Barrufet, M.; et al. Diagnosis and staging of hepatocellular carcinoma (HCC): Current guidelines. Eur. J. Radiol. 2018, 101, 72–81. [Google Scholar] [CrossRef]
- Marquez-Pedroza, J.; Cárdenas-Bedoya, J.; Morán-Moguel, M.C.; Escoto-Delgadillo, M.; Torres-Mendoza, B.M.; Pérez-Ríos, A.M.; González-Enriquez, G.V.; Vázquez-Valls, E. Plasma microRNA expression levels in HIV-1-positive patients receiving antiretroviral therapy. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egaña-Gorroño, L.; Escribà, T.; Boulanger, N.; Guardo, A.C.; León, A.; Bargalló, M.E.; Garcia, F.; Gatell, J.M.; Plana, M.; Arnedo, M. Differential microRNA expression profile between stimulated PBMCs from HIV-1 infected elite controllers and viremic progressors. PloS ONE 2014, 9, e106360. [Google Scholar] [CrossRef]
- Qi, Y.; Hu, H.; Guo, H.; Xu, P.; Shi, Z.; Huan, X.; Zhu, Z.; Zhou, M.; Cui, L. MicroRNA profiling in plasma of HIV-1 infected patients: Potential markers of infection and immune status. J. Public Health Emerg. 2017, 1. [Google Scholar] [CrossRef]
- Garnica, T.K.; Lesbon, J.C.C.; Ávila, A.; Rochetti, A.L.; Matiz, O.R.S.; Ribeiro, R.C.S.; Zoppa, A.; Nishiya, A.T.; Costa, M.T.; de Nardi, A.B.; et al. Liquid biopsy based on small extracellular vesicles predicts chemotherapy response of canine multicentric lymphomas. Sci. Rep. 2020, 10, 20371. [Google Scholar] [CrossRef]
- Khani, A.T.; Sharifzad, F.; Mardpour, S.; Hassan, Z.M.; Ebrahimi, M. Tumor extracellular vesicles loaded with exogenous Let-7i and miR-142 can modulate both immune response and tumor microenvironment to initiate a powerful anti-tumor response. Cancer Lett. 2021, 51, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Eberhardt, M.; Blume, K.; Vera, J.; Baur, A.S. Evidence for liver and peripheral immune cells secreting tumor-suppressive extracellular vesicles in melanoma patients. EBioMedicine 2020, 62, 103119. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Luo, X.; Xie, Y.; Li, P.; Hu, F.; Chu, J.; Chen, X.; Song, W.; Wang, A.; Tian, G.; et al. GC-Derived EVs Enriched with MicroRNA-675-3p Contribute to the MAPK/PD-L1-Mediated Tumor Immune Escape by Targeting CXXC4. Mol. Therapy. Nucleic acids 2020, 22, 615–626. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, F.; Wang, Z.; Tang, L.; Zou, B.; Xu, P.; Yu, T. Hypoxic bone marrow mesenchymal cell-extracellular vesicles containing miR-328-3p promote lung cancer progression via the NF2-mediated Hippo axis. J. Cell. Mol. Med. 2021, 25, 96–106. [Google Scholar] [CrossRef]
- Matthews, O.; Morrison, E.E.; Tranter, J.D.; Starkey Lewis, P.; Toor, I.S.; Srivastava, A.; Sargeant, R.; Rollison, H.; Matchett, K.P.; Kendall, T.J.; et al. Transfer of hepatocellular microRNA regulates cytochrome P450 2E1 in renal tubular cells. EBioMedicine 2020, 62, 103092. [Google Scholar] [CrossRef]
- Qin, Y.; Zhang, J.; Avellán-Llaguno, R.D.; Zhang, X.; Huang, Q. DEHP-elicited small extracellular vesicles miR-26a-5p promoted metastasis in nearby normal A549 cells. Environ. Pollut. 2021, 272, 116005. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Wang, X.; Zou, B.; Mei, J.; Peng, X.; Wu, Z. Extracellular vesicles-encapsulated microRNA-10a-5p shed from cancer-associated fibroblast facilitates cervical squamous cell carcinoma cell angiogenesis and tumorigenicity via Hedgehog signaling pathway. Cancer Gene Ther. 2021, 28, 529–542. [Google Scholar] [CrossRef]
- Lässer, C.; Alikhani, V.S.; Ekström, K.; Eldh, M.; Paredes, P.T.; Bossios, A.; Sjöstrand, M.; Gabrielsson, S.; Lötvall, J.; Valadi, H. Human saliva, plasma and breast milk exosomes contain RNA: Uptake by macrophages. J. Transl. Med. 2011, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Torben, W.; Mansfield, J.; Alvarez, X.; Vande Stouwe, C.; Li, J.; Byrareddy, S.N.; Didier, P.J.; Pahar, B.; Molina, P.E.; et al. Cannabinoid Attenuation of Intestinal Inflammation in Chronic SIV-Infected Rhesus Macaques Involves T Cell Modulation and Differential Expression of Micro-RNAs and Pro-inflammatory Genes. Front. Immunol. 2019, 10, 914. [Google Scholar] [CrossRef]
- Chandra, L.C.; Kumar, V.; Torben, W.; Vande Stouwe, C.; Winsauer, P.; Amedee, A.; Molina, P.E.; Mohan, M. Chronic administration of Delta9-tetrahydrocannabinol induces intestinal anti-inflammatory microRNA expression during acute simian immunodeficiency virus infection of rhesus macaques. J. Virol. 2015, 89, 1168–1181. [Google Scholar] [CrossRef] [Green Version]
- Lyu, Y.; Kaddour, H.; Kopcho, S.; Panzner, T.D.; Shouman, N.; Kim, E.-Y.; Martinson, J.; McKay, H.; Martinez-Maza, O.; Margolick, J.B.; et al. Human Immunodeficiency Virus (HIV) Infection and Use of Illicit Substances Promote Secretion of Semen Exosomes that Enhance Monocyte Adhesion and Induce Actin Reorganization and Chemotactic Migration. Cells 2019, 8, 1027. [Google Scholar] [CrossRef] [Green Version]
- Kechin, A.; Boyarskikh, U.; Kel, A.; Filipenko, M. cutPrimers: A New Tool for Accurate Cutting of Primers from Reads of Targeted Next Generation Sequencing. J. Comput. Biol. 2017, 24, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Licursi, V.; Conte, F.; Fiscon, G.; Paci, P. MIENTURNET: An interactive web tool for microRNA-target enrichment and network-based analysis. BMC Bioinform. 2019, 20, 545. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal ID | Pre-Infection Sample Used | SIV Inoculum | Group 1 VEH/SIV | Group 2 THC/SIV | Group 3 VEH/SIV/cART | Group 4 THC/SIV /cART | Group 5 THC/no SIV | Post-Infection Sample Collection |
|---|---|---|---|---|---|---|---|---|
| JD66 | NA | SIVmac251 | Yes | 1 & 5 MPI | ||||
| IN24 | NA | SIVmac251 | Yes | 1 & 5 MPI | ||||
| JH47 | NA | SIVmac251 | Yes | 1 & 5 MPI | ||||
| JI45 | NA | SIVmac251 | Yes | 1 & 5 MPI | ||||
| JC85 | NA | SIVmac251 | Yes | 1 & 5 MPI | ||||
| JT80 | NA | SIVmac251 | Yes | 1 & 5 MPI | ||||
| LM56 | NA | SIVmac251 | Yes | 1 & 5 MPI | ||||
| LA88 | NA | SIVmac251 | Yes | 1 & 5 MPI | ||||
| LN60 | NA | SIVmac251 | Yes | 1 & 5 MPI | ||||
| LA55 | NA | SIVmac251 | Yes | 1 & 5 MPI | ||||
| KV50 | NA | SIVmac251 | Yes | 1 & 5 MPI | ||||
| LM85 | NA | SIVmac251 | Yes | 1 & 5 MPI | ||||
| HI78 | NA | SIVmac251 | No | 1 & 5 MPT | ||||
| HN79 | NA | SIVmac251 | No | 1 & 5 MPT | ||||
| HN39 | NA | SIVmac251 | No | 1 & 5 MPT |
| Animal IDs | cART Start Timepoint | cART end Timepoint | Control IgG Start Date | Anti-alpha4beta Antibody Start Date | |
|---|---|---|---|---|---|
| Cohort 2 | |||||
| VEH/SIV/cART | LA88 | 14 DPI or 0.5 MPI | 8 MPI | 7 MPI | |
| LC39 | 14 DPI or 0.5 MPI | 8 MPI | 7 MPI | ||
| LD08 | 14 DPI or 0.5 MPI | 8 MPI | 7 MPI | ||
| LE67 | 14 DPI or 0.5 MPI | 8 MPI | 7 MPI | ||
| THC/SIV/cART | LA55 | 14 DPI or 0.5 MPI | 8 MPI | 7 MPI | |
| LB61 | 14 DPI or 0.5 MPI | 8 MPI | 7 MPI | ||
| LA89 | 14 DPI or 0.5 MPI | 8 MPI | 7 MPI | ||
| KV50 | 14 DPI or 0.5 MPI | 8 MPI | 7 MPI | ||
| Cohort 2 | |||||
| VEH/SIV/cART | LM56 | 14 DPI or 0.5 MPI | 6 MPI | 4 MPI | |
| LH75 | 14 DPI or 0.5 MPI | 6 MPI | 4 MPI | ||
| LN60 | 14 DPI or 0.5 MPI | 6 MPI | 4 MPI | ||
| LC48 | 14 DPI or 0.5 MPI | 6 MPI | 4 MPI | ||
| THC/SIV/cART | LH92 | 14 DPI or 0.5 MPI | 6 MPI | 4 MPI | |
| LI81 | 14 DPI or 0.5 MPI | 6 MPI | 4 MPI | ||
| LM85 | 14 DPI or 0.5 MPI | 6 MPI | 4 MPI | ||
| LJ21 | 14 DPI or 0.5 MPI | 6 MPI | 4 MPI |
| Treatment/ Time | miRNA ID | miRNA Counts | miRNA ID | miRNA Counts |
|---|---|---|---|---|
| VEH/SIV | EVs | ECs | ||
| Pre | mml-miR-26a-5p | 2496 | mml-miR-27a-3p | 9578 |
| mml-let-7a-5p | 2368 | mml-miR-27b-3p | 9435 | |
| mml-let-7c-5p | 2360 | mml-miR-191-5p | 8459 | |
| 1 MPI | mml-miR-26a-5p | 1603 | mml-miR-16-5p | 8264 |
| mml-let-7c-5p | 1588 | mml-miR-191-5p | 8171 | |
| mml-let-7a-5p | 1578 | mml-miR-27a-3p | 6244 | |
| 5 MPI | mml-let-7c-5p | 1483 | mml-miR-27a-3p | 9669 |
| mml-let-7a-5p | 1476 | mml-miR-191-5p | 9617 | |
| mml-miR-26a-5p | 1329 | mml-miR-27b-3p | 9531 | |
| VEH/SIV/cART | EVs | ECs | ||
| Pre | mml-let-7a-5p | 402 | mml-miR-16-5p | 2815 |
| mml-let-7c-5p | 398 | mml-miR-27a-3p | 1692 | |
| mml-miR-26a-5p | 299 | mml-miR-27b-3p | 1674 | |
| 1 MPI | mml-let-7c-5p | 1367 | mml-miR-16-5p | 3389 |
| mml-let-7a-5p | 1353 | mml-miR-451 | 1962 | |
| mml-miR-26a-5p | 651 | mml-miR-27a-3p | 1689 | |
| 5 MPI | mml-let-7c-5p | 560 | mml-miR-16-5p | 2589 |
| mml-let-7a-5p | 550 | mml-miR-451 | 1710 | |
| mml-miR-26a-5p | 290 | mml-miR-191-5p | 1590 | |
| THC/SIV | EVs | ECs | ||
| Pre | mml-miR-26a-5p | 6930 | mml-miR-27a-3p | 10775 |
| mml-let-7a-5p | 5818 | mml-miR-27b-3p | 10656 | |
| mml-let-7c-5p | 5796 | mml-miR-16-5p | 10181 | |
| 1 MPI | mml-let-7a-5p | 711 | mml-miR-191-5p | 5314 |
| mml-let-7c-5p | 709 | mml-miR-16-5p | 3706 | |
| mml-miR-26a-5p | 642 | mml-miR-27a-3p | 3615 | |
| 5 MPI | mml-let-7c-5p | 765 | mml-miR-16-5p | 2269 |
| mml-let-7a-5p | 761 | mml-miR-191-5p | 2222 | |
| mml-miR-26a-5p | 640 | mml-miR-27a-3p | 1577 | |
| THC/SIV/cART | EVs | ECs | ||
| Pre | mml-let-7a-5p | 3856 | mml-miR-451 | 18618 |
| mml-let-7c-5p | 3783 | mml-miR-16-5p | 18182 | |
| mml-miR-26a-5p | 1872 | mml-miR-191-5p | 6051 | |
| 1 MPI | mml-let-7c-5p | 1332 | mml-miR-16-5p | 3031 |
| mml-let-7a-5p | 1321 | mml-miR-191-5p | 2945 | |
| mml-miR-26a-5p | 1073 | mml-miR-27a-3p | 2291 | |
| 5 MPI | mml-let-7a-5p | 1792 | mml-miR-16-5p | 3912 |
| mml-let-7c-5p | 1789 | mml-miR-191-5p | 3267 | |
| mml-miR-26a-5p | 1648 | mml-miR-27a-3p | 2376 | |
| THC/no SIV | EVs | ECs | ||
| Pre | mml-let-7a-5p | 2078 | mml-miR-27a-3p | 14099 |
| mml-let-7c-5p | 2069 | mml-miR-27b-3p | 13775 | |
| mml-miR-26a-5p | 1777 | mml-miR-191-5p | 13511 | |
| 1 MPI | mml-let-7c-5p | 2763 | mml-miR-191-5p | 20060 |
| mml-let-7a-5p | 2757 | mml-miR-16-5p | 19156 | |
| mml-miR-26a-5p | 2240 | mml-miR-27a-3p | 18669 | |
| 5 MPI | mml-let-7a-5p | 1705 | mml-miR-191-5p | 27536 |
| mml-let-7c-5p | 1694 | mml-miR-16-5p | 18294 | |
| mml-miR-26a-5p | 1477 | mml-miR-27a-3p | 17992 | |
| Regulation | FC (log2) | p Value | −log (p Value) | Citation | |
|---|---|---|---|---|---|
| miR-484 | |||||
| 1 MPI | Up | 3.473 | 0.0001 | 4.000 | This study |
| 5 MPI | Up | 2.567 | 0.011 | 1.963 | This study |
| miR-107 | |||||
| 1 MPI | Up | 1.755 | 0.0056 | 2.252 | This study |
| 5 MPI | Up | 1.504 | 0.014 | 1.854 | This study |
| miR-206 | |||||
| 1 MPI | Up | 3.429 | 0.0001 | 4.000 | This study |
| 5 MPI | Up | 2.846 | 0.0014 | 2.854 | This study |
| miR-184 | |||||
| 1 MPI | Up | 1.674 | 0.001 | 3.000 | This study |
| 5 MPI | Up | 2.567 | 0.0001 | 4.000 | This study |
| miR-1260b | |||||
| 1 MPI | Up | 2.772 | 0.0086 | 2.066 | This study |
| 5 MPI | Up | 2.326 | 0.0378 | 1.423 | This study |
| miR-6132 | |||||
| 1 MPI | Up | 3.668 | 0.0001 | 4.000 | This study |
| 5 MPI | Up | 3.201 | 0.0001 | 4.000 | This study |
| Regulation | FC (log2) | p Value | −log (p Value) | Citation | |
|---|---|---|---|---|---|
| miR-342-3p | |||||
| 1 MPI | Down | −3.012 | 0.0105 | 1.979 | This study |
| 5 MPI | Down | −2.010 | 0.0444 | 1.353 | This study |
| miR-100-5p | |||||
| 1 MPI | Down | −1.805 | 0.0067 | 2.174 | This study |
| 5 MPI | Down | −1.657 | 0.0175 | 1.757 | This study |
| miR-181b-5p | |||||
| 1 MPI | Down | −2.266 | 0.0057 | 2.244 | This study |
| 5 MPI | Down | −1.680 | 0.028 | 1.587 | This study |
| Regulation | FC (log2) | p Value | −log (p Value) | Citation | |
|---|---|---|---|---|---|
| miR-676-3p | |||||
| 1 MPI | Down | −1.770 | 0.0222 | 1.654 | This study |
| 5 MPI | Down | −2.771 | 0.0175 | 1.757 | This study |
| miR-574 | |||||
| 1 MPI | Down | −2.858 | 0.0031 | 2.509 | This study |
| 5 MPI | Down | −1.856 | 0.0323 | 1.491 | This study |
| miR-505-5p | |||||
| 1 MPI | Down | −2.736 | 0.0073 | 2.137 | This study |
| 5 MPI | Down | −2.736 | 0.0209 | 1.680 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopcho, S.; McDew-White, M.; Naushad, W.; Mohan, M.; Okeoma, C.M. Alterations in Abundance and Compartmentalization of miRNAs in Blood Plasma Extracellular Vesicles and Extracellular Condensates during HIV/SIV Infection and Its Modulation by Antiretroviral Therapy (ART) and Delta-9-Tetrahydrocannabinol (Δ9-THC). Viruses 2023, 15, 623. https://doi.org/10.3390/v15030623
Kopcho S, McDew-White M, Naushad W, Mohan M, Okeoma CM. Alterations in Abundance and Compartmentalization of miRNAs in Blood Plasma Extracellular Vesicles and Extracellular Condensates during HIV/SIV Infection and Its Modulation by Antiretroviral Therapy (ART) and Delta-9-Tetrahydrocannabinol (Δ9-THC). Viruses. 2023; 15(3):623. https://doi.org/10.3390/v15030623
Chicago/Turabian StyleKopcho, Steven, Marina McDew-White, Wasifa Naushad, Mahesh Mohan, and Chioma M. Okeoma. 2023. "Alterations in Abundance and Compartmentalization of miRNAs in Blood Plasma Extracellular Vesicles and Extracellular Condensates during HIV/SIV Infection and Its Modulation by Antiretroviral Therapy (ART) and Delta-9-Tetrahydrocannabinol (Δ9-THC)" Viruses 15, no. 3: 623. https://doi.org/10.3390/v15030623
APA StyleKopcho, S., McDew-White, M., Naushad, W., Mohan, M., & Okeoma, C. M. (2023). Alterations in Abundance and Compartmentalization of miRNAs in Blood Plasma Extracellular Vesicles and Extracellular Condensates during HIV/SIV Infection and Its Modulation by Antiretroviral Therapy (ART) and Delta-9-Tetrahydrocannabinol (Δ9-THC). Viruses, 15(3), 623. https://doi.org/10.3390/v15030623