
Molecular Detection and Genetic Characterization of Japanese Encephalitis Virus in Animals from 11 Provinces in China

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Samples
2.2. Identification of Japanese Encephalitis Virus and Envelope Protein Sequence Amplification
2.3. Sequence Analysis
2.4. Predicted N-Glycosylation and Phosphorylation Sites in JEV E Protein
2.5. Nucleotide Sequence Accession Numbers
3. Results
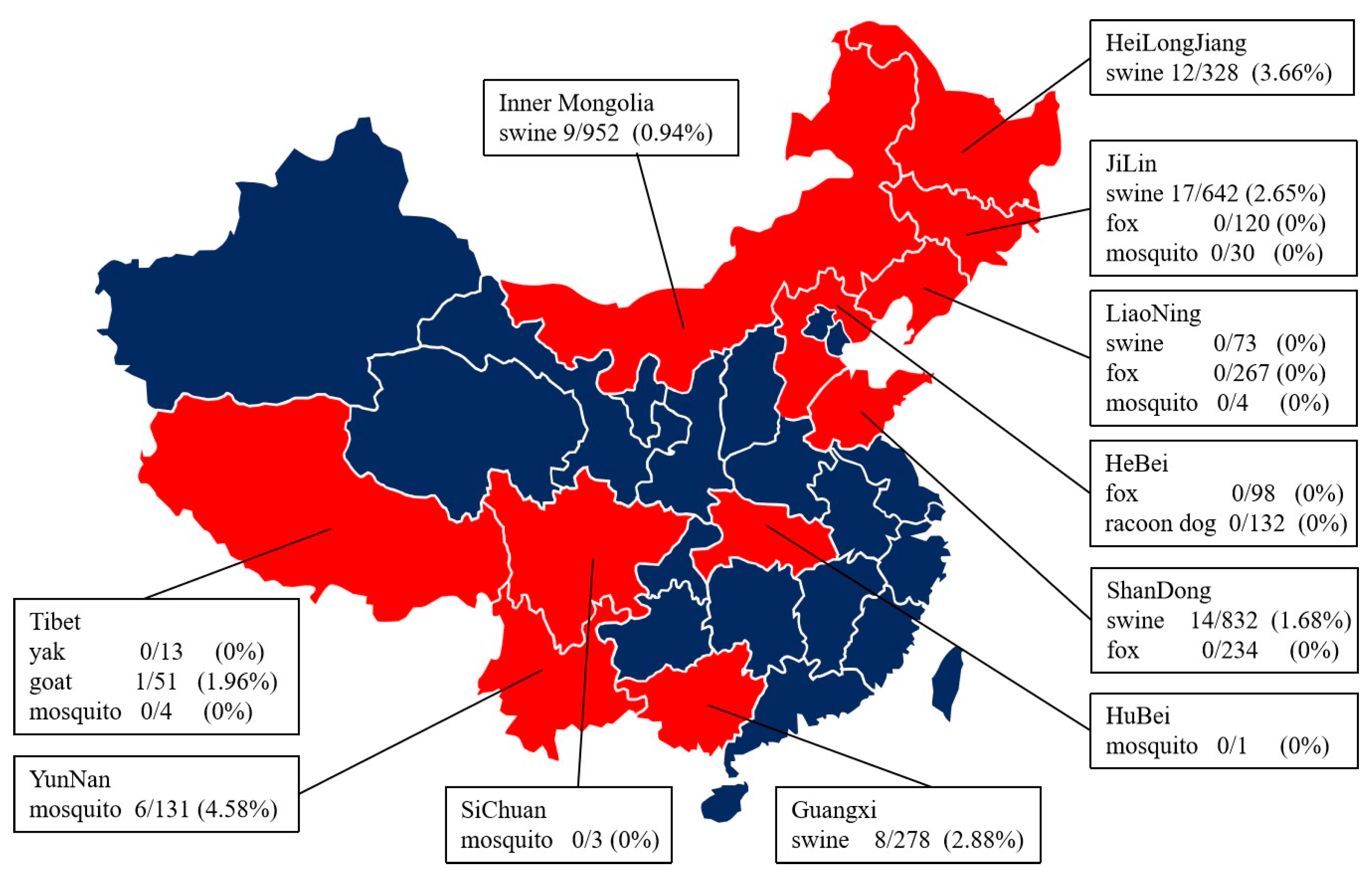
3.1. Samples
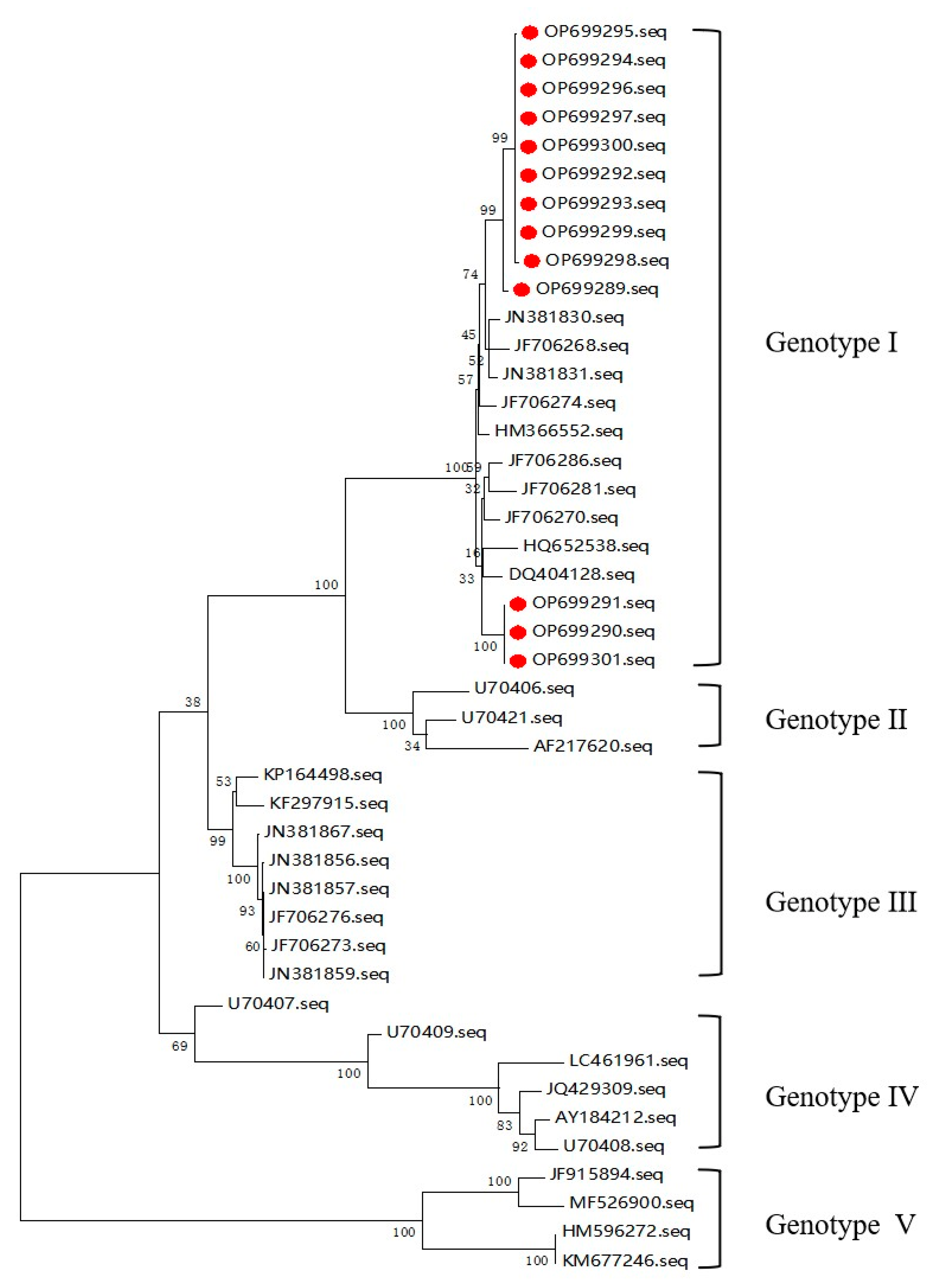
3.2. Phylogenetic Analysis of JEV
3.3. Alterations in Amino Acid Sequences
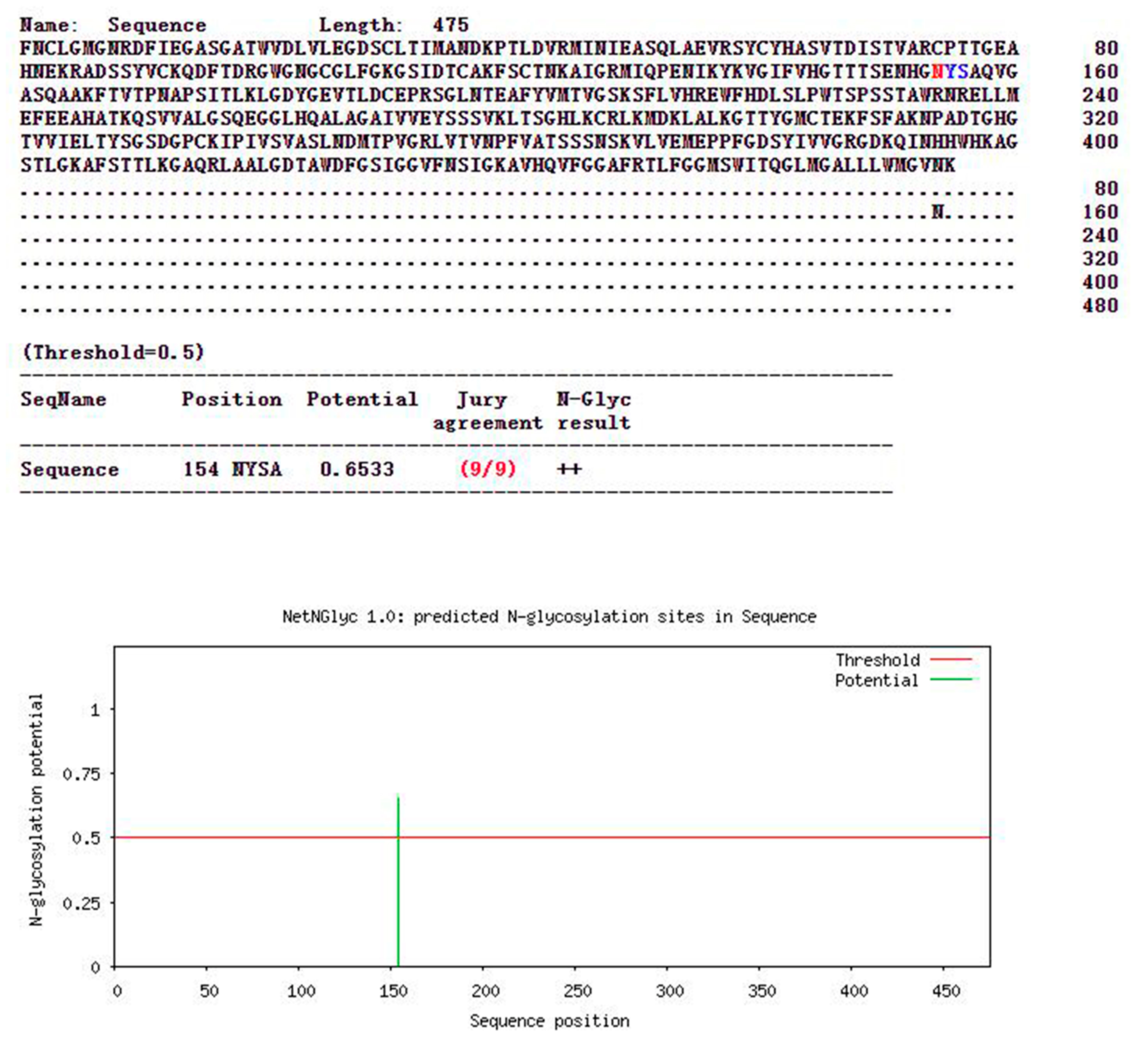
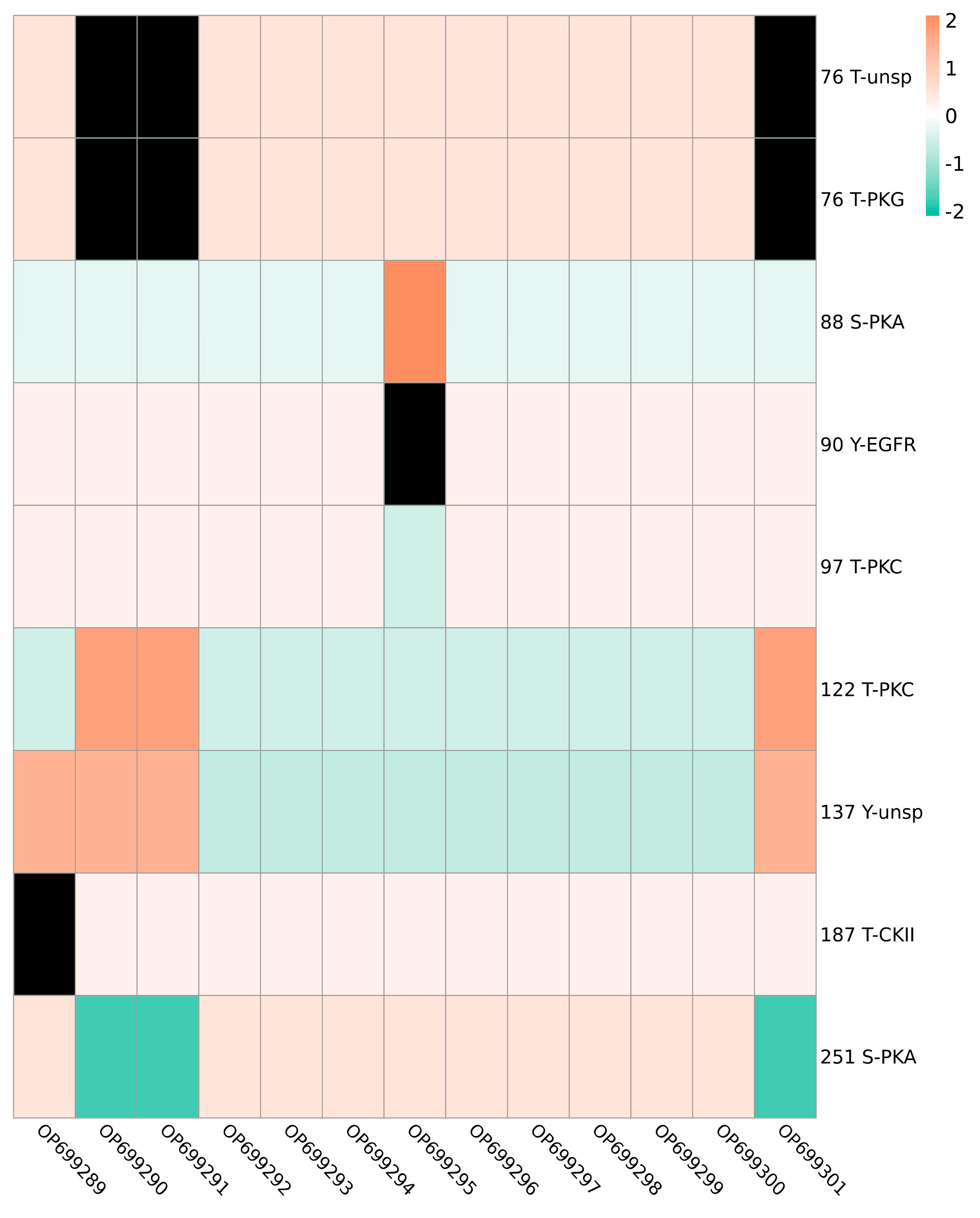
3.4. Glycosylation and Phosphorylation Sites
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Han, N.; Adams, J.; Chen, P.; Guo, Z.-Y.; Zhong, X.-F.; Fang, W.; Li, N.; Wen, L.; Tao, X.-Y.; Yuan, Z.-M.; et al. Comparison of Genotypes I and III in Japanese Encephalitis Virus Reveals Distinct Differences in Their Genetic and Host Diversity. J. Virol. 2014, 88, 11469–11479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Liu, H.; Li, M.; Fu, S.; Liang, G. Insights into the evolutionary history of Japanese encephalitis virus (JEV) based on whole-genome sequences comprising the five genotypes. Virol. J. 2015, 12, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuh, A.J.; Guzman, H.; Tesh, R.B.; Barrett, A.D. Genetic diversity of Japanese encephalitis virus isolates obtained from the Indonesian archipelago between 1974 and 1987. Vector-Borne Zoonotic Dis. 2013, 13, 479–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, L.P.; Bui, T.M.; Phan, N.T. Mechanism of Japanese encephalitis virus genotypes replacement based on human, porcine and mosquito-originated cell lines model. Asian Pac. J. Trop. Med. 2016, 9, 333–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.S.; Hettenbach, S.M.; Park, S.L.; Higgs, S.; Barrett, A.D.; Hsu, W.W.; Harbin, J.N.; Cohnstaedt, L.W.; Vanlandingham, D.L. Differential Infectivities among Different Japanese Encephalitis Virus Genotypes in Culex quinquefasciatus Mosquitoes. PLoS Negl. Trop. Dis. 2016, 10, e0005038. [Google Scholar] [CrossRef] [Green Version]
- Auerswald, H.; Maquart, P.O.; Chevalier, V.; Boyer, S. Mosquito Vector Competence for Japanese Encephalitis Virus. Viruses 2021, 13, 1154. [Google Scholar] [CrossRef]
- Simpson, D.I.; Way, H.J.; Platt, G.S.; Bowen, E.T.; Hill, M.N.; Kamath, S. Arbovirus infections in Sarawak, October 1968–February 1970: GETAH virus isolations from mosquitoes. Trans. R. Soc. Trop. Med. Hyg. 1975, 69, 35–38. [Google Scholar] [CrossRef]
- Ladreyt, H.; Durand, B.; Dussart, P.; Chevalier, V. How Central is the Domestic Pig in the Epidemiological Cycle of Japanese Encephalitis Virus? A Review of Scientific Evidence and Implications for Disease Control. Viruses 2019, 11, 949. [Google Scholar] [CrossRef] [Green Version]
- Le Flohic, G.; Porphyre, V.; Barbazan, P.; Gonzalez, J.P. Review of climate, landscape, and viral genetics as drivers of the Japanese encephalitis virus ecology. PLoS Negl. Trop. Dis. 2013, 7, e2208. [Google Scholar] [CrossRef] [Green Version]
- Ricklin, M.E.; Garcia-Nicolas, O.; Brechbuhl, D.; Python, S.; Zumkehr, B.; Posthaus, H.; Oevermann, A.; Summerfield, A. Japanese encephalitis virus tropism in experimentally infected pigs. Vet. Res. 2016, 47, 34. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, K.L.; Hernandez-Triana, L.M.; Banyard, A.C.; Fooks, A.R.; Johnson, N. Japanese encephalitis virus infection, diagnosis and control in domestic animals. Vet. Microbiol. 2017, 201, 85–92. [Google Scholar] [CrossRef]
- Ladreyt, H.; Auerswald, H.; Tum, S.; Ken, S.; Heng, L.; In, S.; Lay, S.; Top, C.; Ly, S.; Duong, V.; et al. Comparison of Japanese Encephalitis Force of Infection in Pigs, Poultry and Dogs in Cambodian Villages. Pathogens 2020, 9, 719. [Google Scholar] [CrossRef]
- Peiris, J.S.; Amerasinghe, F.P.; Arunagiri, C.K.; Perera, L.P. Japanese encephalitis in Sri Lanka: Comparison of vector and virus ecology in different agro-climatic areas. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 541–548. [Google Scholar] [CrossRef]
- Zhu, Y.; Xu, J.; Lian, S.; Zhang, R.; Hou, J.; Wang, M.; Yan, X. Difference Analysis Between Canine Adenovirus Types 1 And 2. Front. Cell. Infect. Microbiol. 2022, 12, 854876. [Google Scholar] [CrossRef]
- Sun, Y.; Ding, H.; Zhao, F.; Yan, Q.; Li, Y.; Niu, X.; Zeng, W.; Wu, K.; Ling, B.; Fan, S.; et al. Genomic Characteristics and E Protein Bioinformatics Analysis of JEV Isolates from South China from 2011 to 2018. Vaccines 2022, 10, 1303. [Google Scholar] [CrossRef]
- Zhu, Y.; Sun, J.; Yan, M.; Lian, S.; Hu, B.; Lv, S.; Li, Y.; Zhang, Y.; Yan, X. The biological characteristics of the canine adenovirus type 1 from fox and the transcriptome analysis of the infected MDCK cell. Cell Biol. Int. 2021, 45, 936–947. [Google Scholar] [CrossRef]
- Schuh, A.J.; Ward, M.J.; Leigh Brown, A.J.; Barrett, A.D. Dynamics of the emergence and establishment of a newly dominant genotype of Japanese encephalitis virus throughout Asia. J. Virol. 2014, 88, 4522–4532. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Liu, H.; Wang, H.; Fu, S.; Guo, Z.; Liang, G. Southernmost Asia is the source of Japanese encephalitis virus (genotype 1) diversity from which the viruses disperse and evolve throughout Asia. PLoS Negl. Trop. Dis. 2013, 7, e2459. [Google Scholar] [CrossRef] [Green Version]
- Schuh, A.J.; Ward, M.J.; Brown, A.J.; Barrett, A.D. Phylogeography of Japanese encephalitis virus: Genotype is associated with climate. PLoS Negl. Trop. Dis. 2013, 7, e2411. [Google Scholar] [CrossRef] [Green Version]
- Boyer, S.; Durand, B.; Yean, S.; Brengues, C.; Maquart, P.O.; Fontenille, D.; Chevalier, V. Host-Feeding Preference and Diel Activity of Mosquito Vectors of the Japanese Encephalitis Virus in Rural Cambodia. Pathogens 2021, 10, 376. [Google Scholar] [CrossRef]
- Bhattachan, A.; Amatya, S.; Sedai, T.R.; Upreti, S.R.; Partridge, J. Japanese encephalitis in hill and mountain districts, Nepal. Emerg. Infect. Dis. 2009, 15, 1691–1692. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Luo, H.; Rehman, M.U.; Nabi, F.; Li, K.; Lan, Y.; Huang, S.; Zhang, L.; Mehmood, K.; Shahzad, M.; et al. Evidence of JEV in Culex tritaeniorhynchus and pigs from high altitude regions of Tibet, China. J. Vector Borne Dis. 2017, 54, 69. [Google Scholar] [PubMed]
- Impoinvil, D.E.; Baylis, M.; Solomon, T. Japanese encephalitis: On the One Health agenda. Curr. Top. Microbiol. Immunol. 2013, 365, 205–247. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, J.; Chirico, J.; Boqvist, S.; Thu, H.T.V.; Magnusson, U. Occurrence of Japanese encephalitis virus mosquito vectors in relation to urban pig holdings. Am. J. Trop. Med. Hyg. 2012, 87, 1076–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lardeux, F.; Loayza, P.; Bouchite, B.; Chavez, T. Host choice and human blood index of Anopheles pseudopunctipennis in a village of the Andean valleys of Bolivia. Malar. J. 2007, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.W.; Wu, S.C. A functional epitope determinant on domain III of the Japanese encephalitis virus envelope protein interacted with neutralizing-antibody combining sites. J. Virol. 2003, 77, 2600–2606. [Google Scholar] [CrossRef] [Green Version]
- Gromowski, G.D.; Firestone, C.Y.; Whitehead, S.S. Genetic Determinants of Japanese Encephalitis Virus Vaccine Strain SA14-14-2 That Govern Attenuation of Virulence in Mice. J. Virol. 2015, 89, 6328–6337. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yang, H.; Li, Z.; Wang, W.; Lin, H.; Liu, L.; Ni, Q.; Liu, X.; Zeng, X.; Wu, Y.; et al. Envelope Protein Mutations L107F and E138K Are Important for Neurovirulence Attenuation for Japanese Encephalitis Virus SA14-14-2 Strain. Viruses 2017, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Li, X.F.; Ye, Q.; Wang, H.J.; Deng, Y.Q.; Zhu, S.Y.; Zhang, Y.; Li, S.H.; Qin, C.F. Characterization of live-attenuated Japanese encephalitis vaccine virus SA14-14-2. Vaccine 2014, 32, 2675–2681. [Google Scholar] [CrossRef]
- Su, H.L.; Liao, C.L.; Lin, Y.L. Japanese encephalitis virus infection initiates endoplasmic reticulum stress and an unfolded protein response. J. Virol. 2002, 76, 4162–4171. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, P.; Cao, R.; Gu, J. Mutation of putative N-linked glycosylation sites in Japanese encephalitis virus premembrane and envelope proteins enhances humoral immunity in BALB/C mice after DNA vaccination. Virol. J. 2011, 8, 138. [Google Scholar] [CrossRef] [Green Version]
- Dechtawewat, T.; Roytrakul, S.; Yingchutrakul, Y.; Charoenlappanit, S.; Siridechadilok, B.; Limjindaporn, T.; Mangkang, A.; Prommool, T.; Puttikhunt, C.; Songprakhon, P.; et al. Potential Phosphorylation of Viral Nonstructural Protein 1 in Dengue Virus Infection. Viruses 2021, 13, 1393. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Number | Genotype | Origin of Species | Location | Sampling Time |
|---|---|---|---|---|
| JN381830 | I | Mosquito | Henan, China | 2006 |
| JF706268 | I | Mosquito | Yunnan, China | 2009 |
| JN381831 | I | Mosquito | Henan, China | 2004 |
| JF706274 | I | Mosquito | Gansu, China | 2008 |
| HM366552 | I | Mosquito | Guizhou, China | 2006 |
| JF706286 | I | Mosquito | Shandong, China | 2008 |
| JF706281 | I | Mosquito | Yunnan, China | 2005 |
| JF706270 | I | Mosquito | Guangxi, China | 2006 |
| HQ652538 | I | Mosquito | Tibet, China | 2009 |
| DQ404128 | I | Mosquito | Yunnan, China | 1979 |
| U70406 | Ⅱ | Mosquito | Bali, Indonesia | 1981 |
| U70421 | Ⅱ | Mosquito | Kuala Lumpur, Malaysia | 1970 |
| AF217620 | Ⅱ | Human | Australia | 1995 |
| KP164498 | Ⅲ | Swine | India | 2014 |
| KF297915 | Ⅲ | Swine | Guangdong, China | 2009 |
| JN381867 | Ⅲ | Human | Fujian, China | 2002 |
| JN381856 | Ⅲ | Swine | Shanghai, China | 2004 |
| JN381857 | Ⅲ | Mosquito | Guizhou, China | 2004 |
| JF706276 | Ⅲ | Mosquito | Heilongjiang, China | 2002 |
| JF706273 | Ⅲ | Swine | Fujian, China | 2002 |
| JN381859 | Ⅲ | Human | Fujian, China | 2003 |
| U70407 | Ⅳ | Mosquito | Indonesia | 1981 |
| U70409 | Ⅳ | Mosquito | Indonesia | 1981 |
| LC461961 | Ⅳ | Swine | Bali, Indonesia | 2017 |
| JQ429309 | Ⅳ | Mosquito | Bantul, Indonesia | 1981 |
| AY184212 | Ⅳ | Mosquito | Indonesia | 1981 |
| U70408 | Ⅳ | Mosquito | Indonesia | 1981 |
| JF915894 | V | Mosquito | Tibet, China | 2009 |
| MF526900 | V | Mosquito | South Korea | 2015 |
| HM596272 | V | Human | Malaysia | 1952 |
| KM677246 | V | Human | Singapore | 1952 |
| OP699289 | I | Swine | Guangxi, China | 2016 |
| OP699290 | I | Swine | Guangxi, China | 2016 |
| OP699291 | I | Swine | Guangxi, China | 2016 |
| OP699292 | I | Swine | Guangxi, China | 2016 |
| OP699293 | I | Swine | Guangxi, China | 2016 |
| OP699294 | I | Swine | Guangxi, China | 2016 |
| OP699295 | I | Swine | Heilongjiang, China | 2018 |
| OP699296 | I | Swine | Heilongjiang, China | 2018 |
| OP699297 | I | Swine | Heilongjiang, China | 2018 |
| OP699298 | I | Swine | Heilongjiang, China | 2018 |
| OP699299 | I | Swine | Heilongjiang, China | 2018 |
| OP699300 | I | Swine | Jilin, China | 2018 |
| OP699301 | I | Swine | Jilin, China | 2018 |
| 76th Site | 95th Site | 123rd Site | 138th Site | 244th Site | 474th Site | 475th Site | |
|---|---|---|---|---|---|---|---|
| consensus | T | G | S | E | E | N | A |
| OP699289 | S-N | A-K | |||||
| OP699290 | T-M | E-Q | A-K | ||||
| OP699291 | T-M | E-Q | A-K | ||||
| OP699292 | S-N | E-K | A-K | ||||
| OP699293 | S-N | E-K | A-K | ||||
| OP699294 | S-N | E-K | A-K | ||||
| OP699295 | G-D | S-N | E-K | A-K | |||
| OP699296 | S-N | E-K | A-K | ||||
| OP699297 | S-N | E-K | A-K | ||||
| OP699298 | S-N | E-K | N-K | A-R | |||
| OP699299 | S-N | E-K | A-K | ||||
| OP699300 | S-N | E-K | A-K | ||||
| OP699301 | T-M | E-Q | A-K |
| Site | Kinase | Score | Site | Kinase | Score | Site | Kinase | Score |
|---|---|---|---|---|---|---|---|---|
| 40 T | PKC | 0.591 | 210 S | unsp | 0.787 | 344 S | unsp | 0.966 |
| 51 S | DNAPK | 0.564 | 222 S | cdc2 | 0.532 | 349 T | p38MAPK | 0.558 |
| 66 T | PKC | 0.662 | 222 S | DNAPK | 0.531 | 349 T | cdk5 | 0.524 |
| 70 T | PKC | 0.824 | 222 S | PKA | 0.518 | 356 T | PKC | 0.566 |
| 88 S | unsp | 0.985 | 226 T | GSK3 | 0.504 | 363 T | PKC | 0.693 |
| 89 S | PKC | 0.816 | 226 T | PKC | 0.501 | 364 S | unsp | 0.944 |
| 90 Y | unsp | 0.950 | 227 S | unsp | 0.945 | 365 S | PKC | 0.762 |
| 112 S | PKA | 0.545 | 227 S | cdk5 | 0.548 | 368 S | unsp | 0.734 |
| 115 T | PKC | 0.749 | 230 S | PKC | 0.586 | 382 Y | unsp | 0.973 |
| 147 T | unsp | 0.665 | 257 S | DNAPK | 0.621 | 382 Y | INSR | 0.553 |
| 148 T | unsp | 0.627 | 257 S | unsp | 0.532 | 401 S | PKC | 0.773 |
| 148 T | PKC | 0.548 | 277 S | ATM | 0.502 | 401 S | cdc2 | 0.565 |
| 162 S | PKC | 0.666 | 277 S | PKC | 0.818 | 402 T | PKC | 0.786 |
| 162 S | DNAPK | 0.604 | 277 S | unsp | 0.528 | 408 S | PKC | 0.62 |
| 162 S | ATM | 0.539 | 281 T | PKC | 0.575 | 408 S | cdc2 | 0.518 |
| 168 T | PKG | 0.577 | 282 S | PKC | 0.507 | 409 T | cdc2 | 0.537 |
| 183 Y | EGFR | 0.617 | 301 Y | SRC | 0.518 | 410 T | unsp | 0.854 |
| 183 Y | unsp | 0.599 | 305 T | PKC | 0.669 | 410 T | PKC | 0.737 |
| 187 T | unsp | 0.738 | 309 S | PKC | 0.717 | 429 S | CKI | 0.618 |
| 194 S | unsp | 0.954 | 317 T | unsp | 0.933 | 460 T | DNAPK | 0.554 |
| 202 Y | unsp | 0.655 | 327 T | CKI | 0.569 | 460 T | cdc2 | 0.55 |
| 205 T | PKC | 0.853 | 329 S | unsp | 0.994 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, G.; Gao, Y.; Shi, N.; Zhang, S.; Xiao, P.; Zhang, J.; Xie, C.; Ha, Z.; Feng, S.; Li, C.; et al. Molecular Detection and Genetic Characterization of Japanese Encephalitis Virus in Animals from 11 Provinces in China. Viruses 2023, 15, 625. https://doi.org/10.3390/v15030625
Zhao G, Gao Y, Shi N, Zhang S, Xiao P, Zhang J, Xie C, Ha Z, Feng S, Li C, et al. Molecular Detection and Genetic Characterization of Japanese Encephalitis Virus in Animals from 11 Provinces in China. Viruses. 2023; 15(3):625. https://doi.org/10.3390/v15030625
Chicago/Turabian StyleZhao, Guanyu, Yan Gao, Ning Shi, Shiheng Zhang, Pengpeng Xiao, Jiaqi Zhang, Changzhan Xie, Zhuo Ha, Sheng Feng, Chenghui Li, and et al. 2023. "Molecular Detection and Genetic Characterization of Japanese Encephalitis Virus in Animals from 11 Provinces in China" Viruses 15, no. 3: 625. https://doi.org/10.3390/v15030625
APA StyleZhao, G., Gao, Y., Shi, N., Zhang, S., Xiao, P., Zhang, J., Xie, C., Ha, Z., Feng, S., Li, C., Zhang, X., Xie, Y., Yu, N., Zhang, H., Bi, J., & Jin, N. (2023). Molecular Detection and Genetic Characterization of Japanese Encephalitis Virus in Animals from 11 Provinces in China. Viruses, 15(3), 625. https://doi.org/10.3390/v15030625