
A Conserved Acidic Residue in the C-Terminal Flexible Loop of HIV-1 Nef Contributes to the Activity of SERINC5 and CD4 Downregulation
 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids
2.2. Cells Lines
2.3. Viruses and Infectivity Assay
2.4. FACS Analyses
2.5. Western Blotting
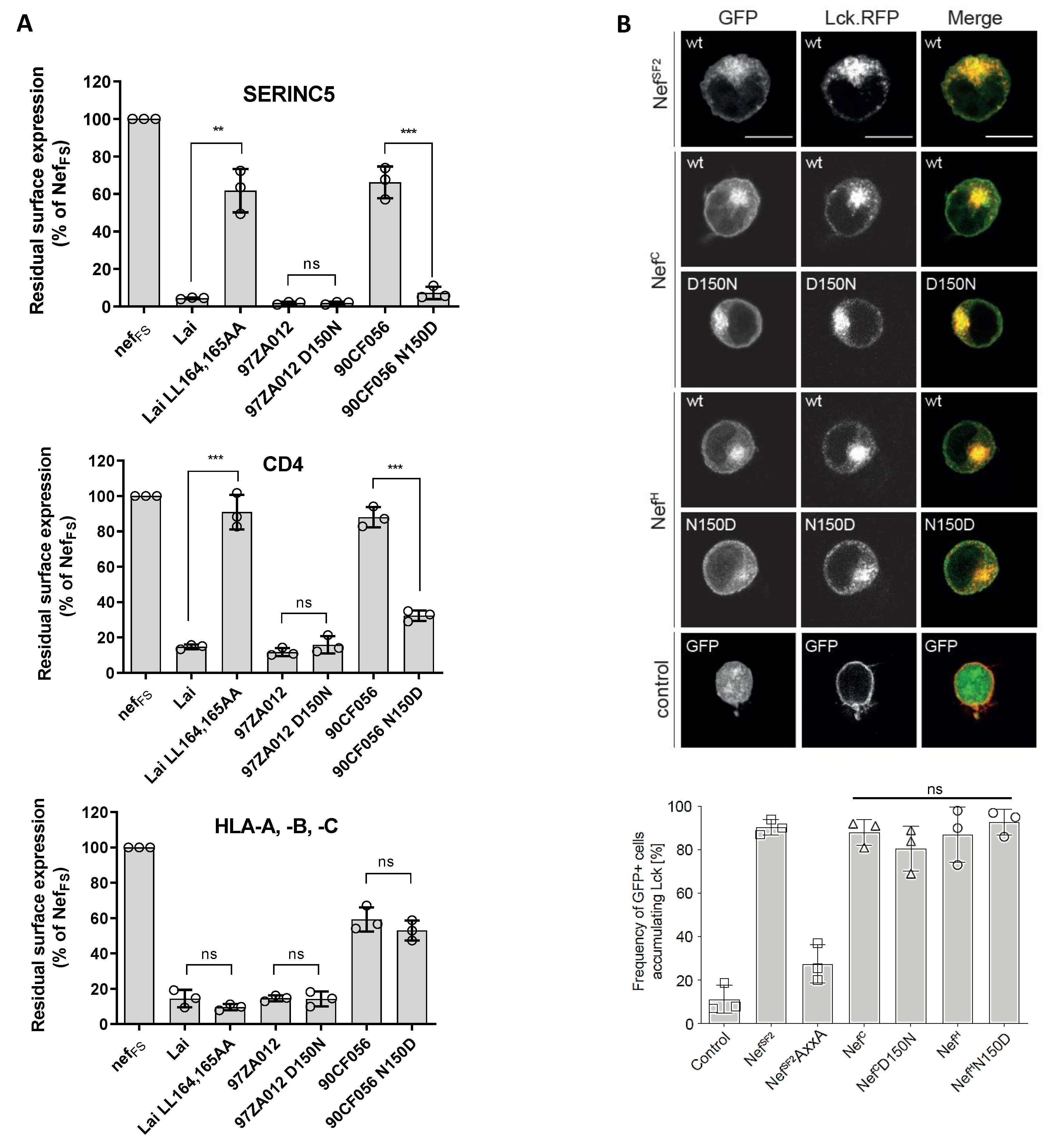
2.6. Lck Retargeting Assay
2.7. BiFC
3. Results
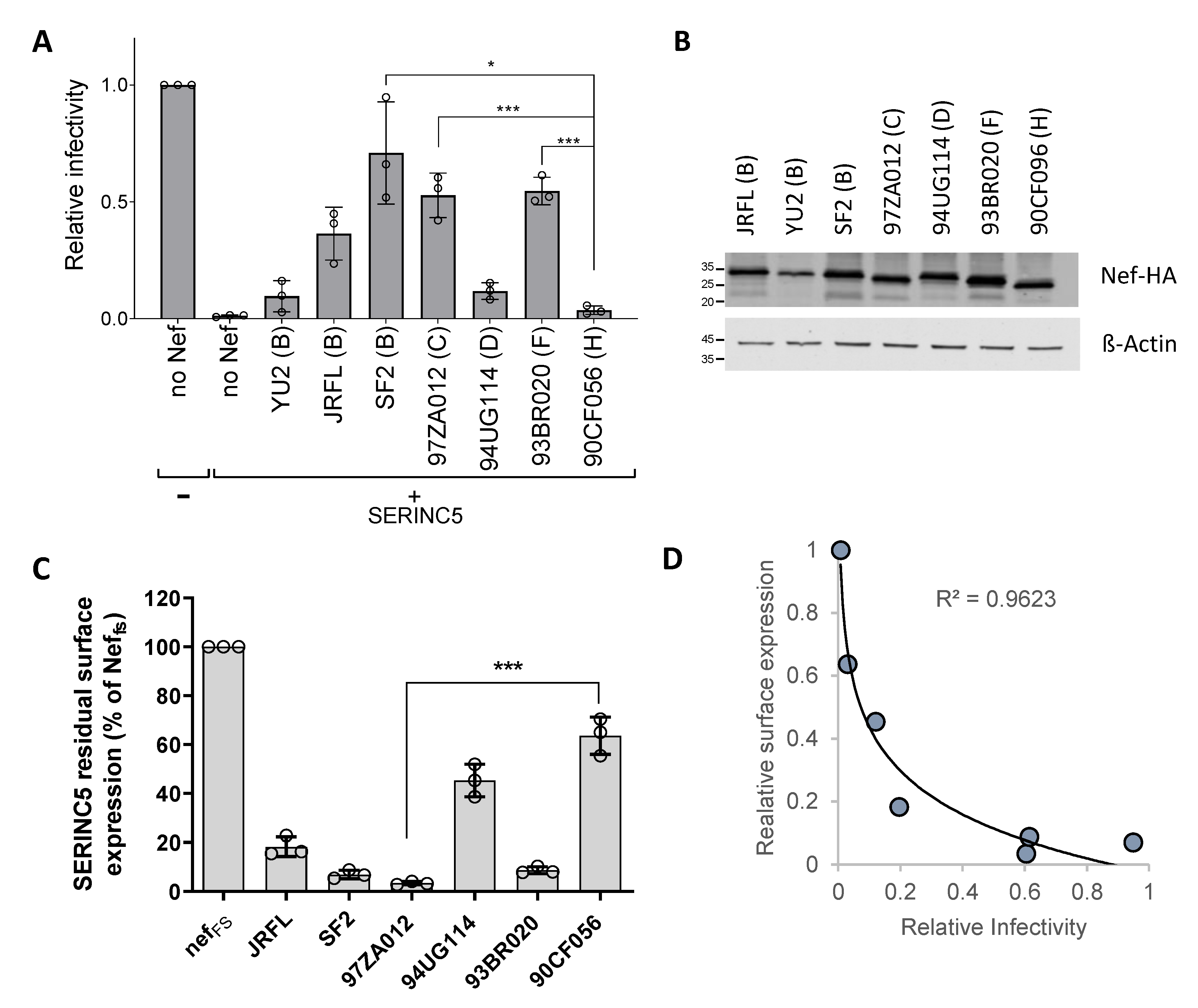
3.1. The Ability of Nef Proteins to Counteract SERINC5 Varies in Different HIV-1 Isolates
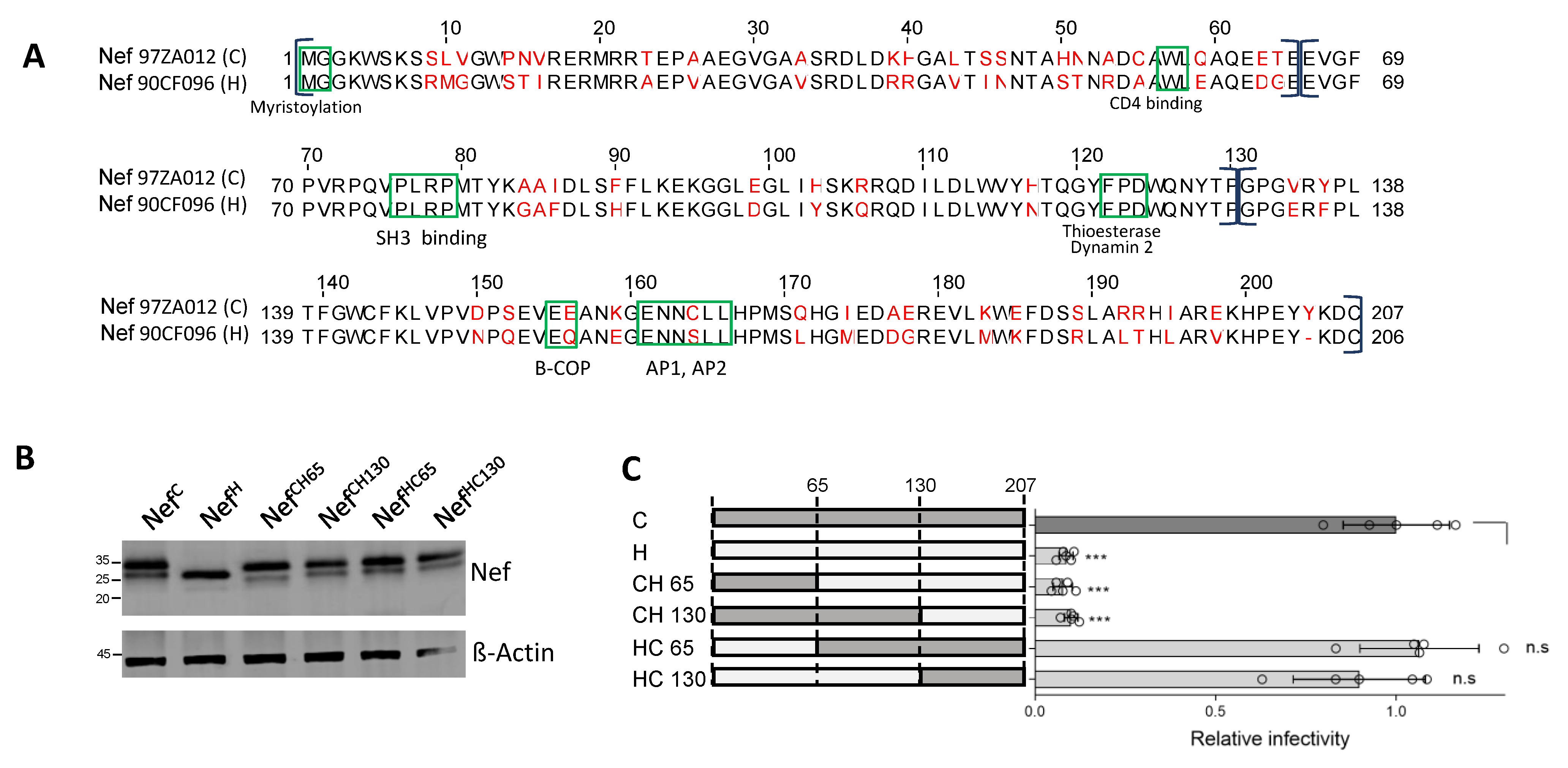
3.2. A Region at the C-Terminus of Nef and Upstream the Leucine-Based Motif Contributes to the Activity against SERINC5
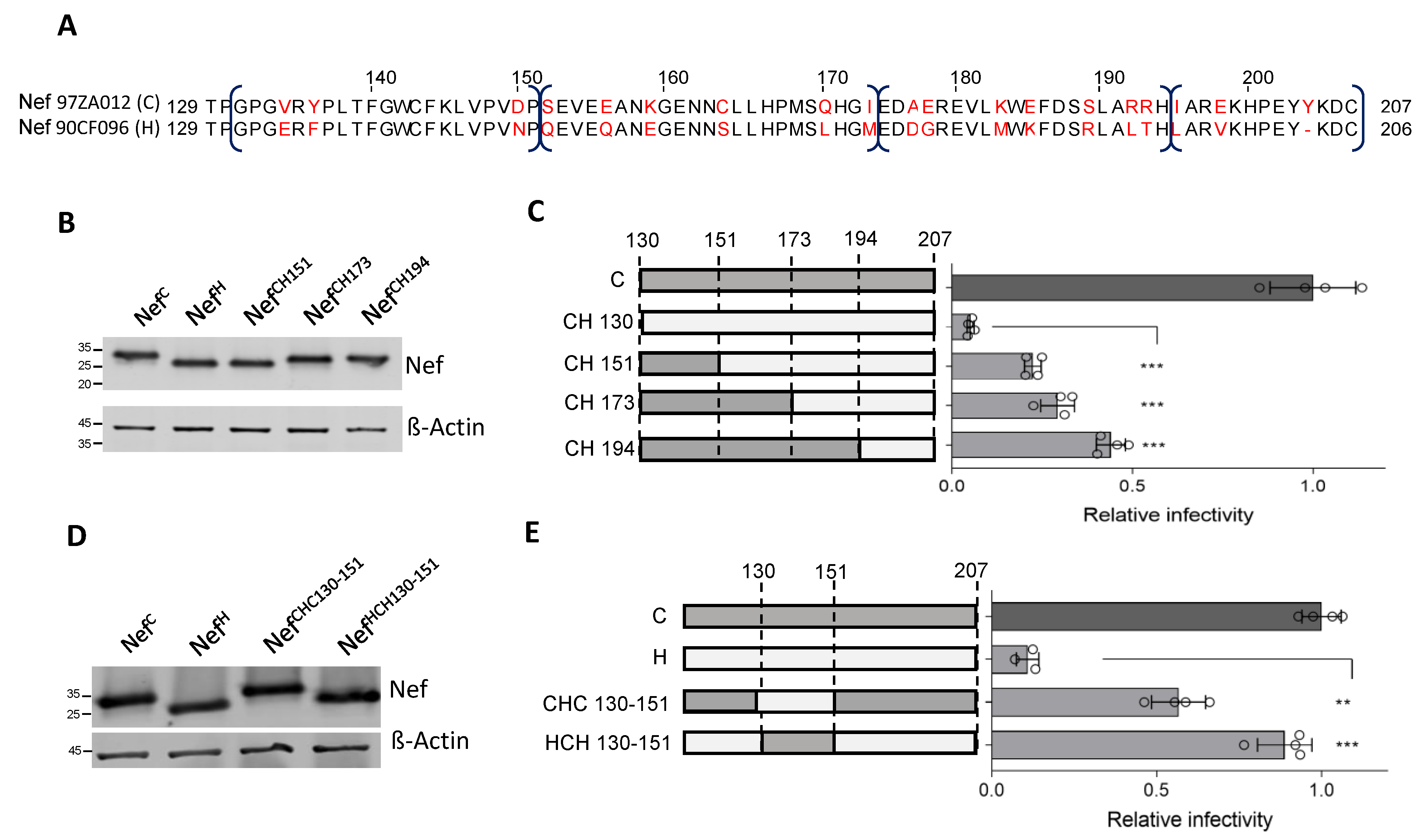
3.3. A 21 aa Stretch of Nef Upstream the Di-Leucine Motif Is Crucial for the Activity against SERINC5
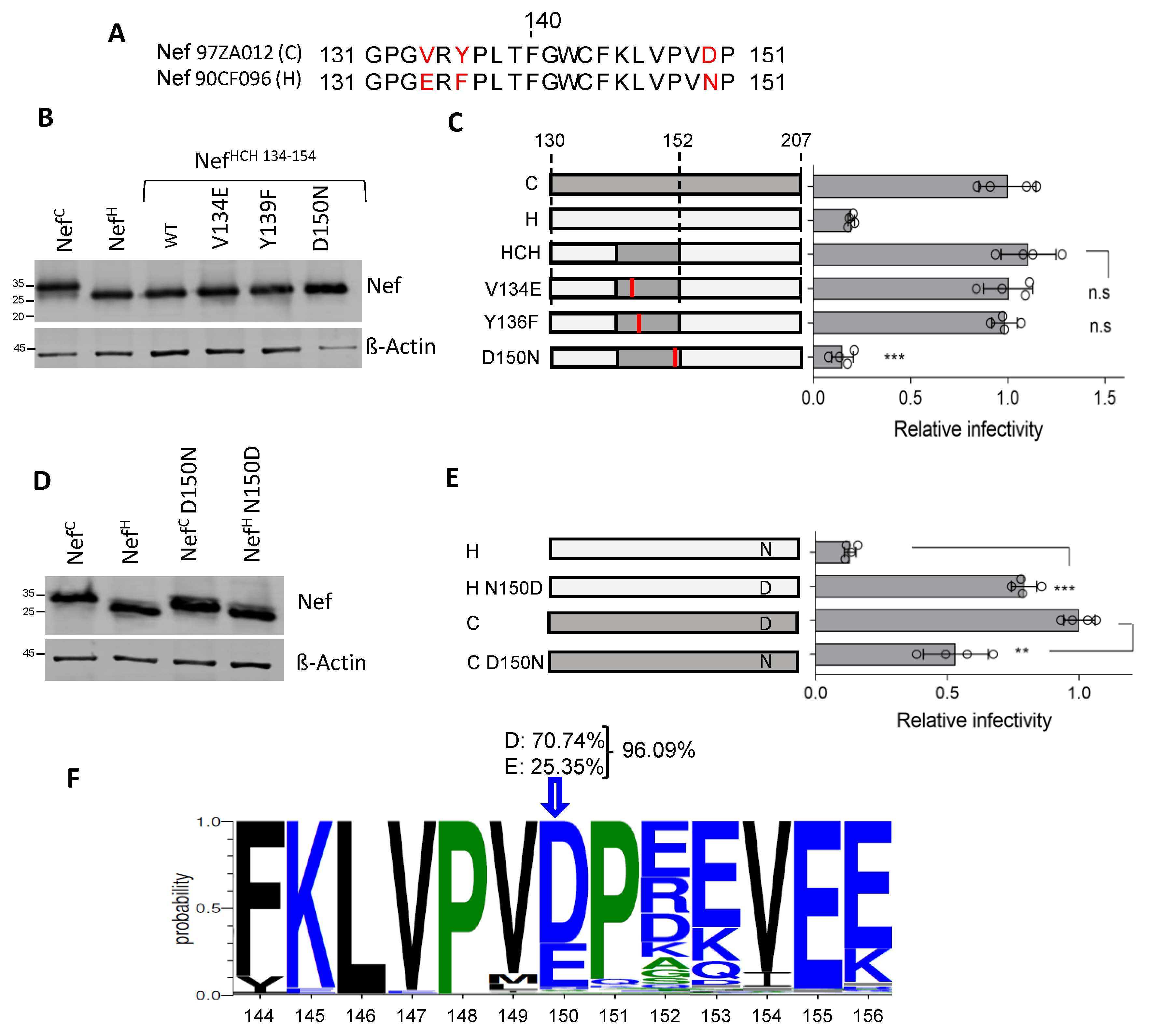
3.4. Residue 150 Is Crucial for the Activity of Nef Clade H on Infectivity
3.5. Residue 150 Is Crucial for the Nef Downregulation Activity of SERINC5 and CD4
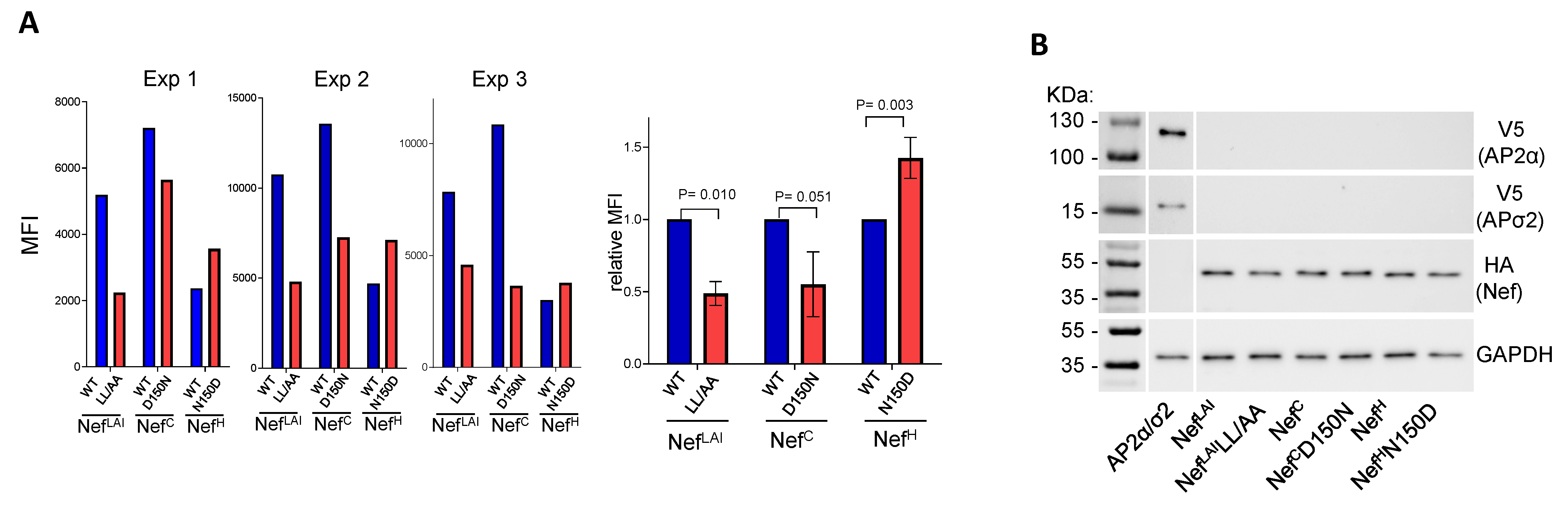
3.6. Residue 150 Is Important for Nef-AP2 Interaction
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kestler, H.W.; Ringler, D.J.; Mori, K.; Panicali, D.L.; Sehgal, P.K.; Daniel, M.D.; Desrosiers, R.C. Importance of the Nef Gene for Maintenance of High Virus Loads and for Development of AIDS. Cell 1991, 65, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Deacon, N.J.; Tsykin, A.; Solomon, A.; Smith, K.; Ludford-Menting, M.; Hooker, D.J.; McPhee, D.A.; Greenway, A.L.; Ellett, A.; Chatfield, C.; et al. Genomic Structure of an Attenuated Quasi Species of HIV-1 from a Blood Transfusion Donor and Recipients. Science 1995, 270, 988–991. [Google Scholar] [CrossRef]
- Kirchhoff, F.; Greenough, T.C.; Brettler, D.B.; Sullivan, J.L.; Desrosiers, R.C. Absence of Intact Nef Sequences in a Long-Term Survivor with Nonprogressive HIV-1 Infection. N. Engl. J. Med. 1995, 332, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Abraham, L.; Fackler, O.T. HIV-1 Nef: A Multifaceted Modulator of T Cell Receptor Signaling. Cell Commun. Signal. 2012, 10, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, E.A.; daSilva, L.L.P. HIV-1 Nef: Taking Control of Protein Trafficking. Traffic 2016, 17, 976–996. [Google Scholar] [CrossRef] [Green Version]
- Buffalo, C.Z.; Iwamoto, Y.; Hurley, J.H.; Ren, X. How HIV Nef Proteins Hijack Membrane Traffic to Promote Infection. J. Virol. 2019, 93, e01322-19. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Singh, R.; Homann, S.; Yang, H.; Guatelli, J.; Xiong, Y. Structural Basis of Evasion of Cellular Adaptive Immunity by HIV-1 Nef. Nat. Struct. Mol. Biol. 2012, 19, 701–706. [Google Scholar] [CrossRef]
- Kwon, Y.; Kaake, R.M.; Echeverria, I.; Suarez, M.; Karimian Shamsabadi, M.; Stoneham, C.; Ramirez, P.W.; Kress, J.; Singh, R.; Sali, A.; et al. Structural Basis of CD4 Downregulation by HIV-1 Nef. Nat. Struct. Mol. Biol. 2020, 27, 822–828. [Google Scholar] [CrossRef]
- Rosa, A.; Chande, A.; Ziglio, S.; De Sanctis, V.; Bertorelli, R.; Goh, S.L.; McCauley, S.M.; Nowosielska, A.; Antonarakis, S.E.; Luban, J.; et al. HIV-1 Nef Promotes Infection by Excluding SERINC5 from Virion Incorporation. Nature 2015, 526, 212–217. [Google Scholar] [CrossRef] [Green Version]
- Usami, Y.; Wu, Y.; Göttlinger, H.G. SERINC3 and SERINC5 Restrict HIV-1 Infectivity and Are Counteracted by Nef. Nature 2015, 526, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Sood, C.; Marin, M.; Chande, A.; Pizzato, M.; Melikyan, G.B. SERINC5 Protein Inhibits HIV-1 Fusion Pore Formation by Promoting Functional Inactivation of Envelope Glycoproteins. J. Biol. Chem. 2017, 292, 6014–6026. [Google Scholar] [CrossRef] [Green Version]
- Ward, A.E.; Kiessling, V.; Pornillos, O.; White, J.M.; Ganser-Pornillos, B.K.; Tamm, L.K. HIV-Cell Membrane Fusion Intermediates Are Restricted by Serincs as Revealed by Cryo-Electron and TIRF Microscopy. J. Biol. Chem. 2020, 295, 15183–15195. [Google Scholar] [CrossRef]
- Heigele, A.; Kmiec, D.; Regensburger, K.; Langer, S.; Peiffer, L.; Stürzel, C.M.; Sauter, D.; Peeters, M.; Pizzato, M.; Learn, G.H.; et al. The Potency of Nef-Mediated SERINC5 Antagonism Correlates with the Prevalence of Primate Lentiviruses in the Wild. Cell Host Microbe 2016, 20, 381–391. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Xiong, R.; Zhou, T.; Su, P.; Zhang, X.; Qiu, X.; Li, H.; Li, S.; Yu, C.; Wang, B.; et al. HIV-1 Nef Antagonizes SERINC5 Restriction by Downregulation of SERINC5 via the Endosome/Lysosome System. J. Virol. 2018, 92, e00196-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trautz, B.; Pierini, V.; Wombacher, R.; Stolp, B.; Chase, A.J.; Pizzato, M.; Fackler, O.T. Antagonism of the SERINC5 Particle Infectivity Restriction by HIV-1 Nef Involves Counteraction of Virion-Associated Pools of the Restriction Factor. J. Virol. 2016; JVI.01246-16. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.W.; Alsahafi, N.; Kuang, X.T.; Swann, S.A.; Toyoda, M.; Göttlinger, H.; Walker, B.D.; Ueno, T.; Finzi, A.; Brumme, Z.L.; et al. Natural HIV-1 Nef Polymorphisms Impair SERINC5 Downregulation Activity. Cell Rep. 2019, 29, 1449–1457.e5. [Google Scholar] [CrossRef] [PubMed]
- Pizzato, M.; Helander, A.; Popova, E.; Calistri, A.; Zamborlini, A.; Palù, G.; Göttlinger, H.G. Dynamin 2 Is Required for the Enhancement of HIV-1 Infectivity by Nef. Proc. Natl. Acad. Sci. USA 2007, 104, 6812–6817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzato, M.; Popova, E.; Göttlinger, H.G. Nef Can Enhance the Infectivity of Receptor-Pseudotyped Human Immunodeficiency Virus Type 1 Particles. J. Virol. 2008, 82, 10811–10819. [Google Scholar] [CrossRef] [Green Version]
- Shu, S.T.; Emert-Sedlak, L.A.; Smithgall, T.E. Cell-Based Fluorescence Complementation Reveals a Role for HIV-1 Nef Protein Dimerization in AP-2 Adaptor Recruitment and CD4 Co-Receptor Down-Regulation. J. Biol. Chem. 2017, 292, 2670–2678. [Google Scholar] [CrossRef] [Green Version]
- Pye, V.E.; Rosa, A.; Bertelli, C.; Struwe, W.B.; Maslen, S.L.; Corey, R.; Liko, I.; Hassall, M.; Mattiuzzo, G.; Ballandras-Colas, A.; et al. A Bipartite Structural Organization Defines the SERINC Family of HIV-1 Restriction Factors. Nat. Struct. Mol. Biol. 2020, 27, 78–83. [Google Scholar] [CrossRef]
- de Sousa-Pereira, P.; Abrantes, J.; Bauernfried, S.; Pierini, V.; Esteves, P.J.; Keppler, O.T.; Pizzato, M.; Hornung, V.; Fackler, O.T.; Baldauf, H.-M. The Antiviral Activity of Rodent and Lagomorph SERINC3 and SERINC5 Is Counteracted by Known Viral Antagonists. J. Gen. Virol. 2019, 100. [Google Scholar] [CrossRef]
- Ananth, S.; Morath, K.; Trautz, B.; Tibroni, N.; Shytaj, I.L.; Obermaier, B.; Stolp, B.; Lusic, M.; Fackler, O.T. Multifunctional Roles of the N-Terminal Region of HIV-1SF2Nef Are Mediated by Three Independent Protein Interaction Sites. J. Virol. 2019, 94, e01398-19. [Google Scholar] [CrossRef] [PubMed]
- Mangasarian, A.; Trono, D. The Multifaceted Role of HIV Nef. Res. Virol. 1997, 148, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Rudolph, J.M.; Abraham, L.; Habermann, A.; Haller, C.; Krijnse-Locker, J.; Fackler, O.T. HIV-1 Nef Compensates for Disorganization of the Immunological Synapse by Inducing Trans-Golgi Network–Associated Lck Signaling. Blood 2012, 119, 786–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, I.; Li, S.; Li, R.; Chai, Q.; Zhang, L.; Wang, B.; Yu, C.; Zheng, Y.-H. The Retroviral Accessory Proteins S2, Nef, and GlycoMA Use Similar Mechanisms for Antagonizing the Host Restriction Factor SERINC5. J. Biol. Chem. 2019, 294, 7013–7024. [Google Scholar] [CrossRef] [PubMed]
- Craig, H.M.; Pandori, M.W.; Guatelli, J.C. Interaction of HIV-1 Nef with the Cellular Dileucine-Based Sorting Pathway Is Required for CD4 down-Regulation and Optimal Viral Infectivity. Proc. Natl. Acad. Sci. USA 1998, 95, 11229–11234. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.W.; Mwimanzi, F.M.; Mann, J.K.; Bwana, M.B.; Lee, G.Q.; Brumme, C.J.; Hunt, P.W.; Martin, J.N.; Bangsberg, D.R.; Ndung’u, T.; et al. Variation in HIV-1 Nef Function within and among Viral Subtypes Reveals Genetically Separable Antagonism of SERINC3 and SERINC5. PLoS Pathog. 2020, 16, e1008813. [Google Scholar] [CrossRef]
- Obermaier, B.; Ananth, S.; Tibroni, N.; Pierini, V.; Shytaj, I.L.; Diaz, R.S.; Lusic, M.; Fackler, O.T. Patient-Derived HIV-1 Nef Alleles Reveal Uncoupling of CD4 Downregulation and SERINC5 Antagonism Functions of the Viral Pathogenesis Factor. J. Acquir. Immune Defic. Syndr. 2020, 85, e23. [Google Scholar] [CrossRef] [PubMed]
- Kruize, Z.; van Nuenen, A.C.; van Wijk, S.W.; Girigorie, A.F.; van Dort, K.A.; Booiman, T.; Kootstra, N.A. Nef Obtained from Individuals with HIV-1 Vary in Their Ability to Antagonize SERINC3- and SERINC5-Mediated HIV-1 Restriction. Viruses 2021, 13, 423. [Google Scholar] [CrossRef] [PubMed]
- Mumby, M.J.; Johnson, A.L.; Trothen, S.M.; Edgar, C.R.; Gibson, R.; Stathopulos, P.B.; Arts, E.J.; Dikeakos, J.D. An Amino Acid Polymorphism within the HIV-1 Nef Dileucine Motif Functionally Uncouples Cell Surface CD4 and SERINC5 Downregulation. J. Virol. 2021, 95, e00588-21. [Google Scholar] [CrossRef]
- Ren, X.; Park, S.Y.; Bonifacino, J.S.; Hurley, J.H. How HIV-1 Nef Hijacks the AP-2 Clathrin Adaptor to Downregulate CD4. eLife 2014, 3, e01754. [Google Scholar] [CrossRef]
- Schaefer, M.R.; Wonderlich, E.R.; Roeth, J.F.; Leonard, J.A.; Collins, K.L. HIV-1 Nef Targets MHC-I and CD4 for Degradation Via a Final Common β-COP–Dependent Pathway in T Cells. PLoS Pathog. 2008, 4, e1000131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal, A.; Sabbaj, S.; Edwards, B.H.; Ritter, D.; Perkins, C.; Tang, J.; Szinger, J.J.; Weiss, H.; Goepfert, P.A.; Korber, B.; et al. T Cell Responses in HIV Type 1-Infected Adolescent Minorities Share Similar Epitope Specificities with Whites Despite Significant Differences in HLA Class I Alleles. AIDS Res. Hum. Retrovir. 2003, 19, 1017–1026. [Google Scholar] [CrossRef]
- Kebba, A.; Kaleebu, P.; Rowland, S.; Ingram, R.; Whitworth, J.; Imami, N.; Gotch, F. Distinct Patterns of Peripheral HIV-1-Specific Interferon-γ Responses in Exposed HIV-1-Seronegative Individuals. J. Infect. Dis. 2004, 189, 1705–1713. [Google Scholar] [CrossRef] [Green Version]
- Kitano, M.; Kobayashi, N.; Kawashima, Y.; Akahoshi, T.; Nokihara, K.; Oka, S.; Takighuchi, M. Identification and Characterization of HLA-B*5401-Restricted HIV-1-Nef and Pol-Specific CTL Epitopes. Microbe. Infect. 2008, 10, 764–772. [Google Scholar] [CrossRef]
- Turnbull, E.L.; Wong, M.; Wang, S.; Wei, X.; Jones, N.A.; Conrod, K.E.; Aldam, D.; Turner, J.; Pellegrino, P.; Keele, B.F.; et al. Kinetics of Expansion of Epitope-Specific T Cell Responses during Primary HIV-1 Infection. J. Immunol. 2009, 182, 7131–7145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.; Chorvinsky, E.; Albihani, S.; Cruz, C.R.; Jones, R.B.; Shpall, E.J.; Margolis, D.M.; Ambinder, R.F.; Bollard, C.M. HIV-Specific T Cells Generated from Naive T Cells Suppress HIV In Vitro and Recognize Wide Epitope Breadths. Mol. Ther. 2018, 26, 1435–1446. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Ganusov, V.V. Defining Kinetic Properties of HIV-Specific CD8+ T-Cell Responses in Acute Infection. Microorganisms 2019, 7, 69. [Google Scholar] [CrossRef] [Green Version]
- Bugembe, D.L.; Ekii, A.O.; Ndembi, N.; Serwanga, J.; Kaleebu, P.; Pala, P. Computational MHC-I Epitope Predictor Identifies 95% of Experimentally Mapped HIV-1 Clade A and D Epitopes in a Ugandan Cohort. BMC Infect. Dis. 2020, 20, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, J.A.; Zhou, S.; Xu, Y.; Moeser, M.J.; MacMillan, D.R.; Council, O.; Kirchherr, J.; Sung, J.M.; Roan, N.R.; Adimora, A.A.; et al. The HIV-1 Latent Reservoir Is Largely Sensitive to Circulating T Cells. eLife 2020, 9, e57246. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Firrito, C.; Bertelli, C.; Rosa, A.; Chande, A.; Ananth, S.; van Dijk, H.; Fackler, O.T.; Stoneham, C.; Singh, R.; Guatelli, J.; et al. A Conserved Acidic Residue in the C-Terminal Flexible Loop of HIV-1 Nef Contributes to the Activity of SERINC5 and CD4 Downregulation. Viruses 2023, 15, 652. https://doi.org/10.3390/v15030652
Firrito C, Bertelli C, Rosa A, Chande A, Ananth S, van Dijk H, Fackler OT, Stoneham C, Singh R, Guatelli J, et al. A Conserved Acidic Residue in the C-Terminal Flexible Loop of HIV-1 Nef Contributes to the Activity of SERINC5 and CD4 Downregulation. Viruses. 2023; 15(3):652. https://doi.org/10.3390/v15030652
Chicago/Turabian StyleFirrito, Claudia, Cinzia Bertelli, Annachiara Rosa, Ajit Chande, Swetha Ananth, Hannah van Dijk, Oliver T. Fackler, Charlotte Stoneham, Rajendra Singh, John Guatelli, and et al. 2023. "A Conserved Acidic Residue in the C-Terminal Flexible Loop of HIV-1 Nef Contributes to the Activity of SERINC5 and CD4 Downregulation" Viruses 15, no. 3: 652. https://doi.org/10.3390/v15030652
APA StyleFirrito, C., Bertelli, C., Rosa, A., Chande, A., Ananth, S., van Dijk, H., Fackler, O. T., Stoneham, C., Singh, R., Guatelli, J., & Pizzato, M. (2023). A Conserved Acidic Residue in the C-Terminal Flexible Loop of HIV-1 Nef Contributes to the Activity of SERINC5 and CD4 Downregulation. Viruses, 15(3), 652. https://doi.org/10.3390/v15030652