
Characterization of Three Different Endolysins Effective against Gram-Negative Bacteria

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Media
2.2. Bacteriophage Cultures
2.3. Phage DNA Isolation and In Silico Analysis
2.4. Cloning of Three Endolysin Genes for Expression
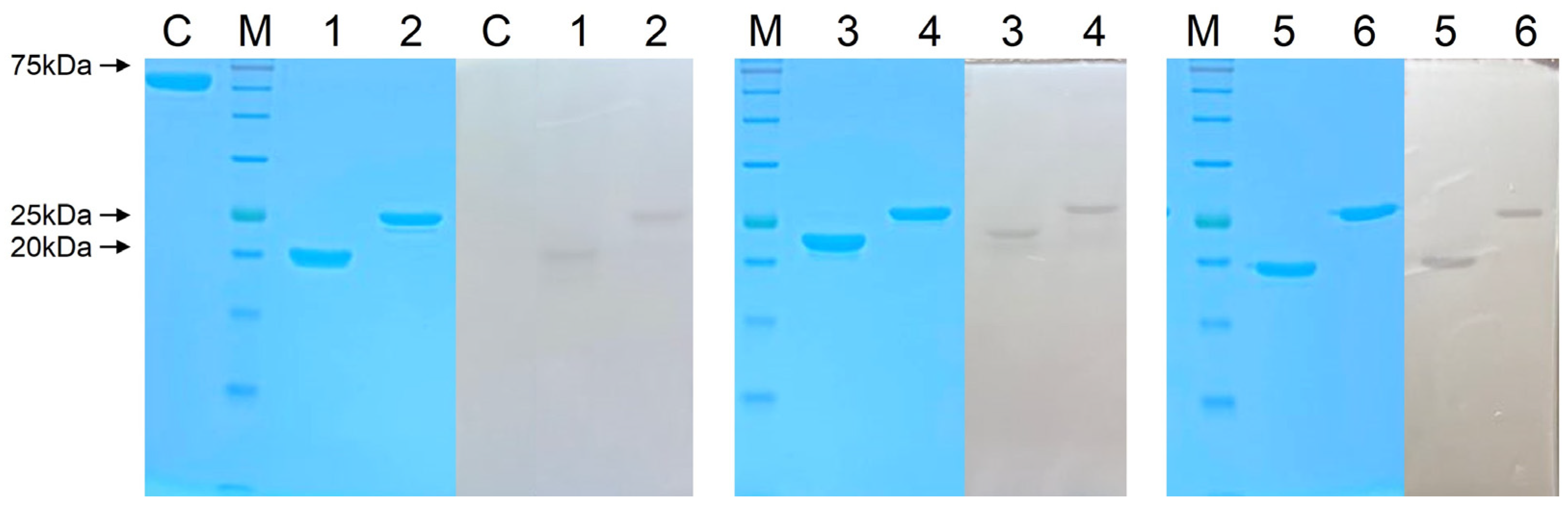
2.5. Overexpression and Purification of Endolysins
2.6. Zymogram Assay
2.7. Colony Forming Unit (CFU) Reduction Assay
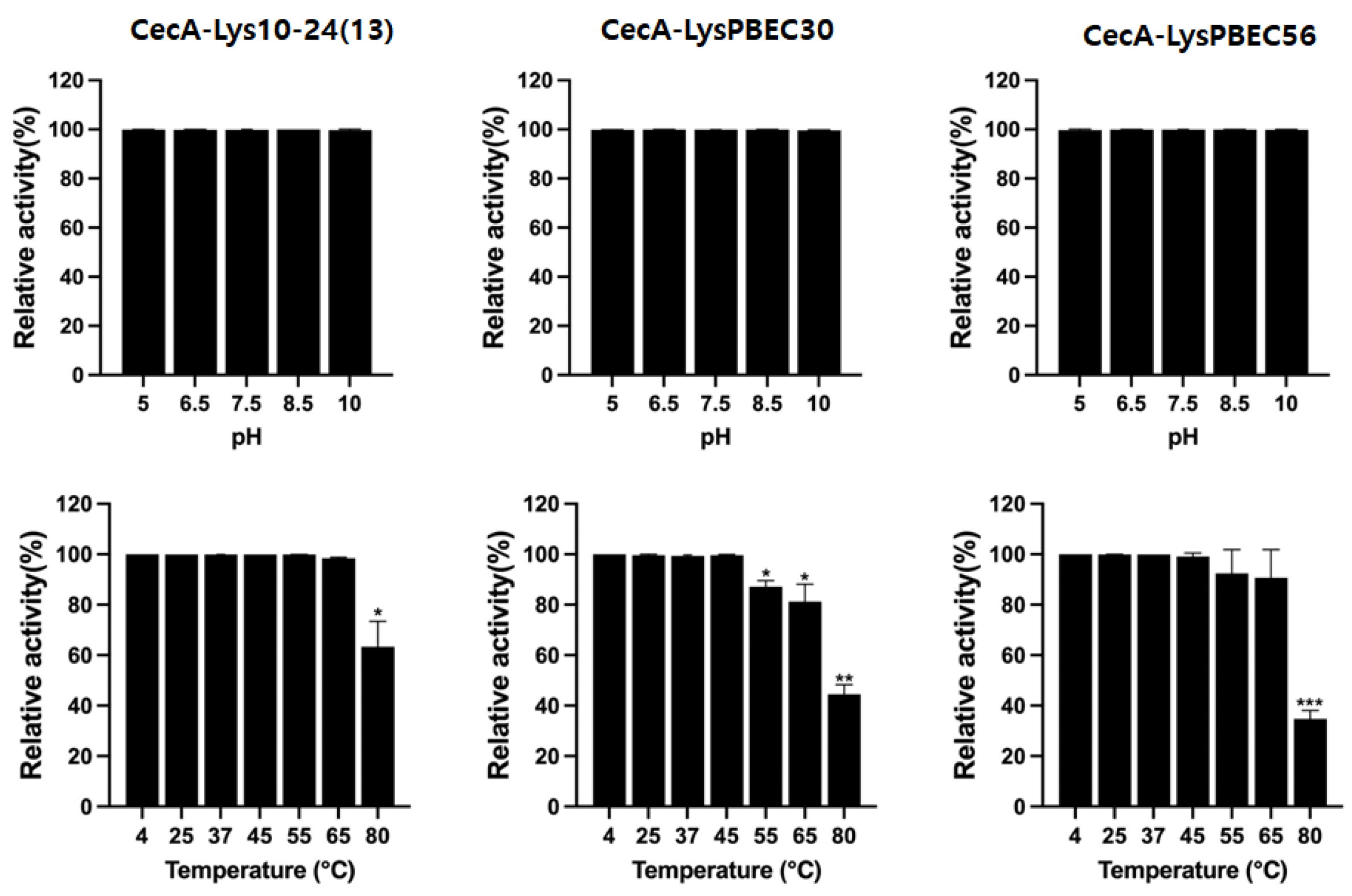
2.8. Stability Test of Endolysins
2.9. Determination of Minimum Inhibitory Concentration (MIC)
2.10. Galleria Mellonella Infection Model
2.11. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Antimicrobial Resistance Collaborators. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Kong, L.; Gao, H.; Cheng, X.; Wang, X. A Review of Current Bacterial Resistance to Antibiotics in Food Animals. Front. Microbiol. 2022, 13, 822689. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Huang, W.E.; Yang, Q. Clinical Perspective of Antimicrobial Resistance in Bacteria. Infect. Drug Resist. 2022, 15, 735. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181. [Google Scholar] [CrossRef]
- Miethke, M.; Pieroni, M.; Weber, T.; Brönstrup, M.; Hammann, P.; Halby, L.; Arimondo, P.B.; Glaser, P.; Aigle, B.; Bode, H.B.; et al. Towards the sustainable discovery and development of new antibiotics. Nature 2021, 5, 726–749. [Google Scholar] [CrossRef] [PubMed]
- Dutescu, I.A.; Hillier, S.A. Encouraging the Development of New Antibiotics: Are Financial Incentives the Right Way Forward? A Systematic Review and Case Study. Infect. Drug Resist. 2021, 14, 415–434. [Google Scholar] [CrossRef]
- Kashani, H.H.; Schmelcher, M.; Sabzalipoor, H.; Hosseini, E.S.; Moniri, R. Recombinant Endolysins as Potential Therapeutics against Antibiotic-Resistant Staphylococcus aureus: Current Status of Research and Novel Delivery Strategies. Clin. Microbiol. Rev. 2017, 31, e00071. [Google Scholar]
- Murray, E.; Draper, L.A.; Ross, R.P.; Hill, C. The Advantages and Challenges of using Endolysins in a Clinical Setting. Viruses 2021, 13, 680. [Google Scholar] [CrossRef]
- Rahman, M.u.; Wang, W.; Sun, Q.; Shah, J.A.; Li, C.; Sun, Y.; Li, Y.; Zhang, B.; Chen, W.; Wang, S. Endolysin, a Promising Solution against Antimicrobial Resistance. Antibiotics 2021, 10, 1277. [Google Scholar] [CrossRef]
- Young, R. Phage lysis: Three steps, three choices, one outcome. J. Microbiol. 2014, 52, 243–258. [Google Scholar] [CrossRef]
- Lee, C.; Kim, J.; Son, B.; Ryu, S. Development of Advanced Chimeric Endolysin to Control Multidrug-Resistant Staphylococcus aureus through Domain Shuffling. ACS Infect. Dis. 2021, 7, 2081. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, Y.; Wang, J.; Zhao, Y.; Zhong, Q.; Li, G.; Fu, Z.; Lu, S. Phage Endolysin LysP108 showed Promising Antibacterial Potential against Methicillin-Resistant Staphylococcus aureus. Front. Cell Infect. Microbiol. 2021, 11, 668430. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Lim, J.; Chang, H.; Park, J. Characteristics and Lytic Activity of Phage-Derived Peptidoglycan Hydrolase, LysSAP8, as a Potent Alternative Biocontrol Agent for Staphylococcus aureus. J. Microbiol. Biotechnol. 2019, 29, 1916–1924. [Google Scholar] [CrossRef]
- Kim, N.; Park, W.B.; Cho, J.E.; Choi, Y.J.; Choi, S.J.; Jun, S.Y.; Kang, C.K.; Song, K.; Choe, P.G.; Bang, J.; et al. Effects of Phage Endolysin SAL200 Combined with Antibiotics on Staphylococcus aureus Infection. Antimicrob. Agents Chemother. 2018, 62, e00731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Mi, Z.; Yin, X.; Fan, H.; An, X.; Zhang, Z.; Chen, J.; Tong, Y. Characterization of Enterococcus faecalis Phage IME-EF1 and its Endolysin. PLoS ONE 2013, 8, e80435. [Google Scholar] [CrossRef]
- Son, B.; Yun, J.; Lim, J.; Shin, H.; Heu, S.; Ryu, S. Characterization of LysB4, an Endolysin from the Bacillus cereus-Infecting Bacteriophage B4. BMC Microbiol. 2012, 12, 33. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Feng, C.; Ren, J.; Zhuang, X.; Zhang, Y.; Zhu, Y.; Dong, K.; He, P.; Guo, X.; Qin, J. A Novel Antimicrobial Endolysin, LysPA26, against Pseudomonas aeruginosa. Front. Microbiol. 2017, 8, 293. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Hu, K.; Xie, Y.; Liu, Y.; Mu, D.; Guo, H.; Zhang, Z.; Zhang, Y.; Chang, D.; Shi, Y. A Novel Phage PD-6A3, and its Endolysin Ply6A3, with Extended Lytic Activity against Acinetobacter baumannii. Front. Microbiol. 2018, 9, 3302. [Google Scholar] [CrossRef] [Green Version]
- Plotka, M.; Kapusta, M.; Dorawa, S.; Kaczorowska, A.; Kaczorowski, T. Ts2631 Endolysin from the Extremophilic Thermus Scotoductus Bacteriophage vB_Tsc2631 as an Antimicrobial Agent against Gram-Negative Multidrug-Resistant Bacteria. Viruses 2019, 11, 657. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Lee, D.; Jin, J.; Kim, J. Antimicrobial Activity of LysSS, a Novel Phage Endolysin, against Acinetobacter baumannii and Pseudomonas aeruginosa. J. Glob. Antimicrob. Rsist. 2020, 22, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.M.; Gondil, V.S.; Li, C.; Jiang, M.; Li, J.; Yu, J.; Wei, H.; Yang, H. A Novel Acinetobacter baumannii Bacteriophage Endolysin LysAB54 with High Antibacterial Activity against Multiple Gram-Negative Microbes. Front. Cell. Infect. Microbiol. 2021, 11, 637313. [Google Scholar] [CrossRef] [PubMed]
- Briers, Y.; Walmagh, M.; Lavigne, R.; Grymonprez, B.; Biebl, M.; Pirnay, J.; Defraine, V.; Michiels, J.; Cenens, W.; Aertsen, A.; et al. Art-175 is a Highly Efficient Antibacterial against Multidrug-Resistant Strains and Persisters of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2014, 58, 3774–3784. [Google Scholar] [CrossRef] [Green Version]
- Defraine, V.; Schuermans, J.; Grymonprez, B.; Govers, S.K.; Aertsen, A.; Fauvart, M.; Michiels, J.; Lavigne, R.; Briers, Y. Efficacy of Artilysin Art-175 against Resistant and Persistent Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 3480–3488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelkader, K.; Gutiérrez, D.; Tamés-Caunedo, H.; Ruas-Madiedo, P.; Safaan, A.; Khairalla, A.S.; Gaber, Y.; Dishisha, T.; Briers, Y. Engineering a Lysin with Intrinsic Antibacterial Activity (LysMK34) by Cecropin A Fusion Enhances its Antibacterial Properties against Acinetobacter baumannii. Appl. Environ. Microbiol. 2022, 88, e0151521. [Google Scholar] [CrossRef] [PubMed]
- Antonova, N.P.; Vasina, D.V.; Lendel, A.M.; Usachev, E.V.; Makarov, V.V.; Gintsburg, A.L.; Tkachuk, A.P.; Gushchin, V.A. Broad Bactericidal Activity of the Myoviridae Bacteriophage Lysins LysAm24, LysECD7, and LysSi3 against Gram-Negative ESKAPE Pathogens. Viruses 2019, 11, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.; Hong, J.; Jung, Y.; Ha, J.; Kim, H.; Myung, H.; Song, M. Bactericidal Effect of Cecropin A Fused Endolysin on Drug-Resistant Gram-Negative Pathogens. J. Microbiol. Biotechnol. 2022, 32, 816–823. [Google Scholar] [CrossRef]
- Lim, J.; Jang, J.; Myung, H.; Song, M. Eradication of drug-resistant Acinetobacter baumannii by cell-penetrating peptide fused endolysin. J. Microbiol. 2022, 60, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Kim, Y.D.; Jang, J.; Kim, M.S.; Song, M.; Myung, H. Combination Effect of Engineered Endolysin EC340 with Antibiotics. Front. Microbiol. 2022, 13, 821936. [Google Scholar] [CrossRef]
- Vázquez, R.; García, E.; García, P. Sequence-Function Relationships in Phage-Encoded Bacterial Cell Wall Lytic Enzymes and Their Implications for Phage-Derived Product Design. J. Virol. 2021, 95, e0032121. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Briers, Y. Lysins Breaking Down the Walls of Gram-Negative Bacteria, no Longer a no-Go. Curr. Opin. Biotechnol. 2021, 68, 15–22. [Google Scholar] [CrossRef]
- Heselpoth, R.D.; Euler, C.W.; Schuch, R.; Fischetti, V.A. Lysocins: Bioengineered Antimicrobials that Deliver Lysins Across the Outer Membrane of Gram-Negative Bacteria. Antimicrob. Agents Chemother. 2019, 63, e00342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvestro, L.; Weiser, J.N.; Axelsen, P.H. Antibacterial and Antimembrane Activities of Cecropin A in Escherichia coli. Antimicrob. Agents Chemother. 2000, 44, 602–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vudumula, U.; Adhikari, M.D.; Ojha, B.; Goswami, S.; Das, G.; Ramesh, A. Tuning the Bactericidal Repertoire and Potency of Quinoline-Based Amphiphiles for Enhanced Killing of Pathogenic Bacteria. RSC Adv. 2012, 2, 3864. [Google Scholar] [CrossRef]
- Kim, S.; Patel, D.; Park, S.; Slusky, J.; Klauda, J.; Widmalm, G.; Im, W. Bilayer Properties of Lipid A from various Gram-Negative Bacteria. Biophys. J. 2016, 111, 1750–1760. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.; Hu, X.; Liao, M.; Fa, K.; Ciumac, D.; Clifton, L.A.; Sani, M.; King, S.M.; Maestro, A.; Separovic, F.; et al. Structural Disruptions of the Outer Membranes of Gram-Negative Bacteria by Rationally Designed Amphiphilic Antimicrobial Peptides. ACS Appl. Mater. Interfaces 2021, 13, 16062. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MIC (μg/mL) | ||
|---|---|---|---|
| CecA-Lys10-24(13) | CecA-LysPBEC30 | CecA-LysPBEC56 | |
| E. coli | |||
| ATCC 8739 | 8 | 16 | 16 |
| K-12 MG1655 | 4 | 32 | 4 |
| ATCC 25922 | 4 | 16 | 8 |
| ATCC 700927 | >64 | >64 | >64 |
| F611 | >64 | >64 | >64 |
| F703 | 64 | >64 | >64 |
| F716 | 16 | 32 | 32 |
| F852 | 16 | >64 | 32 |
| F859 | 8 | 64 | 64 |
| F862 | 4 | 4 | 16 |
| F906 | 32 | >64 | 64 |
| P. aeruginosa | |||
| PAO1 | 8 | 32 | 32 |
| ATCC 15522 | 8 | >64 | 16 |
| ATCC 15692 | 8 | 32 | 16 |
| ATCC 13388 | 4 | 4 | 8 |
| ATCC 10145 | >64 | >64 | >64 |
| ATCC 9027 | 64 | >64 | 64 |
| ATCC 27853 | 16 | >64 | 16 |
| F147 | 16 | 32 | 32 |
| F102 | 32 | >64 | >64 |
| F265 | >64 | >64 | >64 |
| F388 | 16 | 16 | 16 |
| F341 | 16 | 32 | 32 |
| A. baumannii | |||
| ATCC 19606 | 4 | 4 | 8 |
| ATCC 17978 | 32 | 16 | 32 |
| KACC 13090 | 8 | 32 | 16 |
| KACC 14233 | 16 | 32 | 32 |
| K3680 | 16 | 64 | 16 |
| K643 | 16 | 64 | 64 |
| F4 | >64 | 32 | 64 |
| F15 | 16 | 64 | 16 |
| F65 | 64 | 64 | 32 |
| F66 | 16 | 8 | 16 |
| F67 | 8 | 16 | 16 |
| F68 | 32 | 64 | 64 |
| F69 | 64 | 64 | 64 |
| K. pneumoniae | |||
| KCTC 2208 | 4 | 4 | 4 |
| KCTC 2296 | 8 | 8 | 16 |
| KCTC 2246 | 16 | >64 | 16 |
| ATCC 700603 | 16 | >64 | >64 |
| F120 | 32 | 16 | 64 |
| F147 | 16 | >64 | 64 |
| F126 | 8 | 8 | 16 |
| F85 | 32 | >64 | >64 |
| E. aerogenes | |||
| F276 | 16 | >64 | >64 |
| E. cloacae | |||
| ATCC 13047 | 8 | 8 | 32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, T.-H.; Hong, H.-W.; Kim, M.S.; Song, M.; Myung, H. Characterization of Three Different Endolysins Effective against Gram-Negative Bacteria. Viruses 2023, 15, 679. https://doi.org/10.3390/v15030679
Jeong T-H, Hong H-W, Kim MS, Song M, Myung H. Characterization of Three Different Endolysins Effective against Gram-Negative Bacteria. Viruses. 2023; 15(3):679. https://doi.org/10.3390/v15030679
Chicago/Turabian StyleJeong, Tae-Hwan, Hye-Won Hong, Min Soo Kim, Miryoung Song, and Heejoon Myung. 2023. "Characterization of Three Different Endolysins Effective against Gram-Negative Bacteria" Viruses 15, no. 3: 679. https://doi.org/10.3390/v15030679
APA StyleJeong, T. -H., Hong, H. -W., Kim, M. S., Song, M., & Myung, H. (2023). Characterization of Three Different Endolysins Effective against Gram-Negative Bacteria. Viruses, 15(3), 679. https://doi.org/10.3390/v15030679