
Tizoxanide Antiviral Activity on Dengue Virus Replication

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. Determination of Viremia and Protein Concentration in Dengue-Virus-Infected Cells
2.4. Viral Titration by Plaque Assay
2.5. Pre-Treatment Test
2.6. Assay of Inhibition of Virus Adsorption
2.7. Virus Penetration Inhibition Assay
2.8. Virucidal Activity
2.9. Post-Infection Assay
2.10. Dose–Response Inhibition Curve
2.11. Virus Release Assay
2.12. Real-Time Polymerase Chain Reaction via One-Step Reverse Transcription
2.13. Statistical Analysis
2.14. Protein Digestion to Proteomics Analysis
2.15. Liquid Chromatography-Tandem Mass Spectrometry (LC/MS/MS) Analysis
2.16. Protein Identification and Analysis
3. Results
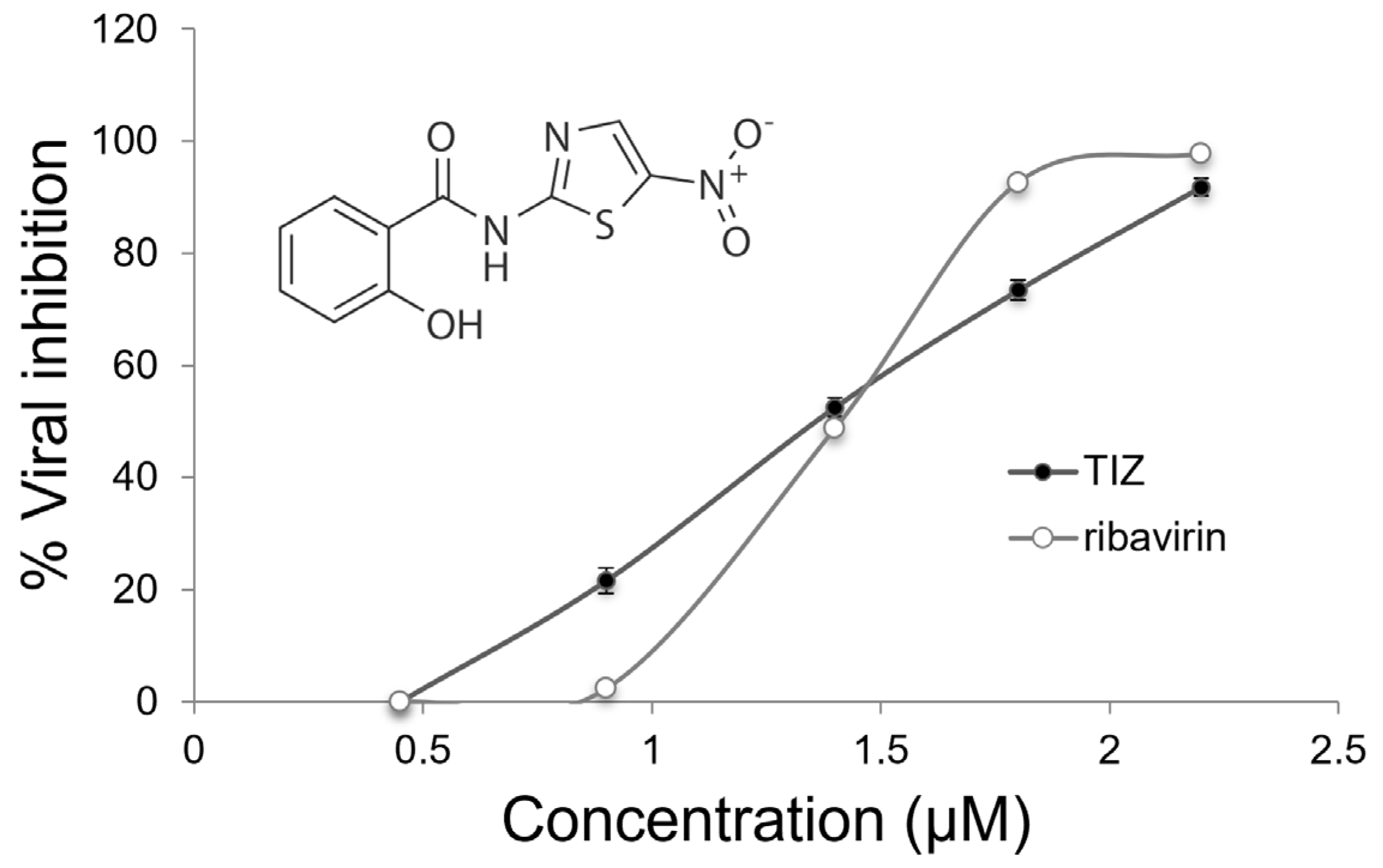
3.1. Antiviral Activity
3.2. Antiviral Effects against DENV-2
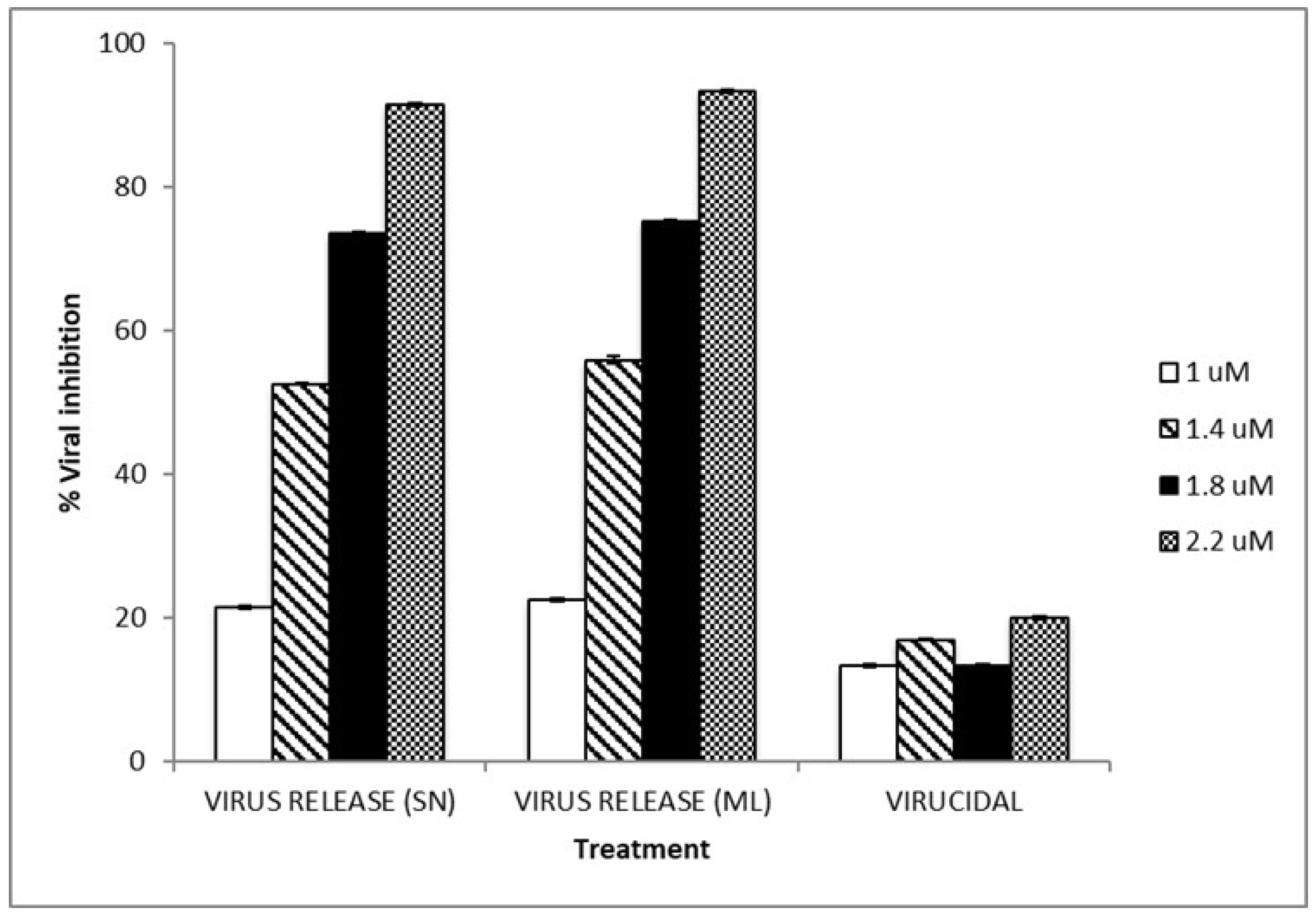
3.3. TIZ Inhibits Viral Infection
3.4. Loss of Infectivity of the Viral Particles by TIZ
3.5. Infectivity of Viral Particles and RT-PCR
3.6. Proteomics Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pierson, T.C.; Diamond, M.S. Degrees of maturity: The complex structure and biology of flaviviruses. Curr. Opin. Virol. 2012, 2, 168–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Racz, M.L. Epidemiologia das infecções virais. In Microbiologia; Trabulsi, L.R., Alterthum, F., Eds.; Atheneu: São Paulo, Brazil, 2008; pp. 573–576. [Google Scholar]
- Salles, T.S.; Sá-Guimarães, T.D.E.; De Alvarenga, E.S.L.; Guimarães-Ribeiro, V.; De Meneses, M.D.F.; De Castro-Salles, P.F.; Dos Santos, C.R.; Melo, A.C.D.A.; Soares, M.R.; Ferreira, D.F.; et al. History, epidemiology and diagnostics of dengue in the American and Brazilian contexts: A review. Parasit. Vectors 2018, 11, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, M.I.; Richardson, J.H.; Sanchez-Vargas, I.; Olson, K.E.; Beaty, B.J. Dengue virus type 2: Replication and tropisms in orally infected Aedes aegypti mosquitoes. BCM Microbiol. 2007, 7, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamond, M.S.; Pierson, T.C. Molecular Insight into Deng ue Virus Pathogenesis and Its Implications for Disease Control. Cell 2015, 162, 488–492. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Dengue Vaccine: WHO Position Paper–September 2018; Weekly Epidemiological Record; World Health Organization: Geneva, Switzerland, 2018; Volume 93, pp. 457–476. [Google Scholar]
- Brownlee, K.A.; Hamre, D.A. Studies on chemotherapy of vaccínia vírus 1. An experimental design for testing antiviral agents. J. Bacteriol. 1951, 61, 127–134. [Google Scholar] [CrossRef] [Green Version]
- De Clercq, E.; Li, G. Approved Antiviral Drugs over the Past 50 Years. Clin. Microbiol. Rev. 2016, 29, 695–747. [Google Scholar] [CrossRef] [Green Version]
- De Clercq, E. Recent highlights in the development of new antiviral drugs. Curr. Opin. Microbiol. 2005, 8, 552–560. [Google Scholar] [CrossRef]
- Antonelli, G.; Turriziani, O. Antiviral therapy: Old and current issues. Int. Antimicrob. Agents 2012, 40, 95–102. [Google Scholar] [CrossRef]
- Wigg, M.D. Antivirais. In Introdução à Virologia Humana, 2nd ed.; Santos, N.S.O., Romanos, M.T.V., Wigg, M.D., Eds.; Guanabara Koogan: São Paulo, Brazil, 2008; pp. 117–146. [Google Scholar]
- Kühl, N.; Graf, D.; Bock, J.; Behnam, M.A.M.; Leuthold, M.M.; Klein, C.D. A New Class of Dengue and West Nile Virus Protease Inhibitors with Submicromolar Activity in Reporter Gene DENV-2 Protease and Viral Replication Assays. J. Med. Chem. 2020, 63, 8179–8197. [Google Scholar] [CrossRef]
- Kaptein, S.J.F.; Goethals, O.; Kiemel, D.; Marchand, A.; Kesteleyn, B.; Bonfanti, J.-F.; Bardiot, D.; Stoops, B.; Jonckers, T.H.M.; Dallmeier, K.; et al. A pan-serotype dengue virus inhibitor targeting the NS3–NS4B interaction. Nature 2021, 598, 504–509. [Google Scholar] [CrossRef]
- Panda, K.; Alagarasu, K.; Patil, P.; Agrawal, M.; More, A.; Kumar, N.; Mainkar, P.; Parashar, D.; Cherian, S. In Vitro Antiviral Activity of α-Mangostin against Dengue Virus Serotype-2 (DENV-2). Molecules 2021, 26, 3016. [Google Scholar] [CrossRef] [PubMed]
- Ansori, A.N.M.; Fadholly, A.; Proboningrat, A.; Hayaza, S.; Susilo, R.J.K.; Naw, S.W.; Posa, G.A.V.; Yusrizal, Y.F.; Sibero, M.T.; Sucipto, T.H.; et al. In Vitro Antiviral Activity of Pinus merkusii (Pinaceae) Stem Bark and Cone against Dengue Virus Type-2 (DENV-2). Res. J. Pharm. Technol. 2021, 14, 3705–3708. [Google Scholar] [CrossRef]
- Euanorasetr, J.; Intra, B.; Thunmrongsiri, N.; Limthongkul, J.; Ubol, S.; Anuegoonpipat, A.; Kurosu, T.; Ikuta, K.; Nihira, T.; Panbangred, W. In vitro antiviral activity of spirotetronate compounds against dengue virus serotype 2. J. Gen. Appl. Microbiol. 2019, 65, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Forrest, J.C.; Zhang, X. A screen of the NIH collection small molecule library identifies potential antiviral drugs. Antivir. Res. 2015, 114, 1–10. [Google Scholar] [CrossRef]
- Rossignol, J.F. Nitazoxanide, the first of thiazolide, in the treatment of a broad range of DNA and RNA viruses. Antivir. Res. 2014, 110, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Gekonge, B.; Badin, M.C.; Montaner. L.J. Short Communication: Nitazoxanide inhibits HIV viral replication in monocyte derived macrophages. AIDS Res. Hum. Retrovir. 2015, 31, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.A.; Blackburn, K.; Migowski, E.; Goshe, M.B.; Brown, D.T.; Ferreira, D.F.; Soares, M.R. Quantitative proteomic analysis of the tizoxanide effect in vero cells. Sci. Rep. 2020, 10, 14733. [Google Scholar] [CrossRef] [PubMed]
- Rocco, P.R.; Silva, P.L.; Cruz, F.F.; Melo-Junior, M.A.C.; Tierno, P.F.; Moura, M.A.; De Oliveira, L.F.G.; Lima, C.C.; Dos Santos, E.A.; Junior, W.F.; et al. Early use of nitazoxanide in mild COVID-19 disease: Randomised, placebo-controlled trial. Eur. Respir. J. 2021, 58, 2003725. [Google Scholar] [CrossRef]
- Chang, J.; Schul, W.; Butters, T.D.; Yip, A.; Liu, B.; Goh, A.; Lakshminarayana, S.B.; Alonzi, D.; Reinkensmeier, G.; Pan, X.; et al. Combination of α-glucosidase inhibitor and ribavirin for the treatment of dengue virus infection in vitro and in vivo. Antivir. Res. 2011, 89, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Dulbecco, R.; Vogt., M. Some problems of animal virology as studied by the plaque technique. Cold Spring Harb. Symp. Quant. Biol. 1953, 18, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.M.; Cheng, H.Y.; Lin, T.C.; Chiang, L.C.; Lin, C.C. Acetone, ethanol and methanol extracts of Phyllanthus urinaria inhibit HSV-2 infection in vivo. Antivir. Res. 2005, 67, 24–30. [Google Scholar] [CrossRef]
- Zhu, W.; Chiu, L.C.M.; Ooi, V.E.C.; Chang, P.K.S.; Ang, P.O., Jr. Antiviral property and mode of action of a sulphated polysaccharide from Sargassum patens against herpes simplex virus type 2. Int. J. Antimicrob. Agents 2004, 24, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-T.; Hsu, J.T.-A.; Hsieh, H.-P.; Lin, P.-H.; Chen, T.-C.; Kao, C.-L.; Lee, C.-N.; Chang, S.-Y. Anti-HSV activity of digitoxin and its possible mechanisms. Antivir. Res. 2008, 79, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.Y.; Lin, C.C.; Lin, T.C. Antiherpes simplex virus type 2 activity of casuarinin from the bark of Terminalia arjuna Linn. Antivir. Res. 2002, 55, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.E.; Kuster, R.M.; Yamamoto, K.A.; Salles, T.S.; Campos, R.; Meneses, M.D.F.; Soares, M.R.; Ferreira, D.F. Quercetin and quercetin e-O-glycosides from Bauhinia longifolia (Bong.) Steud. show anti-Mayaro virus activity. Paras. Vectors. 2014, 7, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized ppb-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A peptide search engine intefrated intro the MaxQuant environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef]
- Torres-Flores, J.M.; Reyes-Sandoval, A.; Salazar, M.I. Dengue Vaccines: An Update. BioDrugs 2022, 36, 325–336. [Google Scholar] [CrossRef]
- Guzman, M.G.; Gubler, D.J.; Izquierdo, A.; Martinez, E.; Halstead, S.B. Dengue infection. Nat. Rev. Dis. Prim. 2016, 2, 16055. [Google Scholar] [CrossRef] [PubMed]
- Philipson, L. Attachment of viruses to cell receptors and penetration of viruses into cells. In Textbook of Medical Virology; Lycke, E., Norrby, E., Eds.; Butterworth & Co.: London, UK, 1983; pp. 51–56. [Google Scholar] [CrossRef]
- Rossignol, J.F.; Elfert, A.; El-Gohary, Y.; Elfert, A.; Keeffe, E.B. Thiazolides, a new class of anti-influenza molecules targeting viral hemagglutinin at post-translational level. J. Biol. Chem. 2009, 284, 29798–29808. [Google Scholar] [CrossRef] [Green Version]
- La Frazia, S.; Ciucci, A.; Arnoldi, F.; Coira, M.; Gianferretti, P.; Angelini, M.; Belardo, G.; Burrone, O.R.; Rossignol, J.-F.; Santoro, M.G. Thiazolides, a new class of antiviral agents effective against rotavirus infection, target viral morphogenesis inhibiting viroplasm formation. J. Virol. 2013, 87, 11096–11106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelegyna, L.; Hautala, T.; Seppänen, M.; Adebayo, A.; Sullivan, K.E.; Icenogle, J. Inhibition of rubella virus replication by the broad-spectrum drug nitazoxanide in cell culture and in a patient with a primary immune deficiency. Antivir. Res. 2017, 147, 58–66. [Google Scholar]
- Shi, Z.; Wei, J.; Deng, X.; Li, S.; Qiu, Y.; Shao, D.; Li, B.; Zhang, K.; Xue, F.; Wang, X.; et al. Nitazoxanide inhibits the replication of Japanese encephalitis virus in cultured cells and in a mouse model. Virol. J. 2014, 11, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elazar, M.; Liu, M.; McKenna, S.A.; Liu, P.; Gehrig, E.A.; Puglisi, J.D.; Rossignol, J.; Glenn, J.S. The anti-hepatitis C agent nitazoxanide induces phosphorylation of eukaryotic initiation factor 2á via protein kinase activated by double-stranded RNA activation. Gastroenterology 2009, 137, 1827–1835. [Google Scholar] [CrossRef] [PubMed]
- Clemens, M.J. PKR—A protein kinase regulated by double-stranded RNA. Int. J. Biochem. Cell Biol. 1997, 29, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Ashiru, O.; Howe, J.D.; Butters, T.D. Nitazoxanide, an antiviral thiazolide, deplets ATP-sensitive intracellular Ca2+ stores. Virology 2014, 462–463, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Béthune, J.; Wieland, F.; Moelleken, J. COPI-mediated transport. J. Membr. Biol. 2006, 211, 65–79. [Google Scholar] [CrossRef]
- Miller, S.; Krijnse-Locker, J. Modification of intracellular membrane structures for virus replication. Nat. Rev. Microbiol. 2008, 6, 363–374. [Google Scholar] [CrossRef]
- Korolchuk, V.I.; Menzies, F.M.; Rubinsztein, D.C. Mechanisms of cross-talk between the ubiquitin-proteasome and autophagy-lysosome systems. FEBS Lett. 2010, 584, 1393–1398. [Google Scholar] [CrossRef] [Green Version]
- Monteith, G.R.; Mcandrew, D.; Faddy, H.M.; Roberts-thomson, S.J. Calcium and cancer: Targeting Ca2+ transport. Nat. Rev. Cancer 2007, 7, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Dally, S.; Bredoux, R.; Corvazier, E.; Andersen, J.P.; Clausen, J.D.; Dode, L.; Fanchaouy, M.; Gelebart, P.; Monceau, V.; Del Monte, F.; et al. Ca2+-ATPases in non-failing and failing heart: Evidence for a novel cardiac sarco/endoplasmic reticulum Ca2+-APTase 2 isoform (SERCA2c). Biochem. J. 2006, 395, 249–258. [Google Scholar] [CrossRef]
- Kaufman, R.J. Double-stranded RNA-activated protein kinase mediates virus-induced apoptosis: A new role for an old actor. Proc. Natl. Acad. Sci. USA 1999, 96, 11693–11695. [Google Scholar] [CrossRef] [Green Version]
- Schröer, M. Endoplasmic reticulum stress responses. Cell Mol. Life Sci. 2008, 65, 862–894. [Google Scholar] [CrossRef] [PubMed]
- Caspersen, C.; Pedersen, P.S.; Treiman, M. The sarco/endoplasmic reticulum calcium-ATPase 2b is an edoplasmic reticulum stress-inducible protein. J. Biol. Chem. 2000, 275, 22363–22372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.C.; Zhou, Y.; Yang, C.Z.; Xiong, D.S. A review of ERGIC-53: Its structure, functions, regulation and relations with diseases. Histol. Histopathol. 2009, 24, 1193–1204. [Google Scholar] [CrossRef]
- Appenzeller-Herzog, C.; Hanuri, H.P. The ER-Golgi intermediate compartment (ERGIC): In search of its identity and function. J. Cell Sci. 2006, 119, 2173–2183. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.Y.; Kawasaki, N.; Hu, D.; Tozawa, H.; Matsumoto, N.; Yamamoto, K. Subcellular localization of ERGIC-53 under endoplasmic reticulum stress condition. Glycobiology 2012, 22, 1709–1720. [Google Scholar] [CrossRef] [Green Version]
- Earp, L.J.; Delos, S.E.; Park, H.E.; White, J.M. The many mechanisms of viral membrane fusion proteins. Curr. Top. Microbiol. Immunol. 2005, 285, 25–66. [Google Scholar] [CrossRef]
- Klaus, J.P.; Eisenhauer, P.; Russo, J.; Mason, A.B.; Do, D.; King, B.; Taatjes, D.; Cornillez-Ty, C.; Boyson, J.E.; Thali, M.; et al. The intracelular cargo receptor ERGIC-53 is required for the production of infectious aernavirus, coronavirus, and filovirus particles. Cell Host Microbe 2013, 14, 522–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiasny, K.; Fritz, R.; Panger, L.K.; Heinz, F.X. Molecular mechanisms of flavivirus membrane fusion. Amino Acids 2011, 41, 1159–1163. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Rape, M. Building ubiquitin chains: E2 enzymes at work. Nat. Rev. Mol. Cell Biol. 2009, 10, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wijk, S.J.; Timmers, H.T. The family of ubiquitin-conjugating enzymes (E2s): Deciding between life and death of proteins. FASEB J. 2010, 24, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.L.; Zhou, H.; Pastushok, L.; Moraes, T.; McKenna, S.; Ziola, B.; Ellison, M.J.; Dixit, V.M.; Xiao, W. Distinct regulation of Ubc13 functions by the two ubiquitin-conjugating enzyme variants Mms2 and Uev1A. J. Cell Biol. 2005, 170, 745–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzic, J.; Marinovic-Terzic, I.; Ikeda, F.; Dikic, I. Ubiquitin signals in the NF-κβ pathway. Biochem. Soc. Trans. 2007, 35, 942–945. [Google Scholar] [CrossRef]
- Nawa, M. Effects of bafilomycin on Japanese encephalitis virus in C6/36 mosquito cells. Arch. Virol. 1998, 143, 1555–1568. [Google Scholar] [CrossRef]
- Nawa, M.; Takasaki, T.; Yamada, K.I.; Kurane, I.; Akatsuka, T. Interference in Japanese encephalitis virus infection of Vero cells by a cationic amphphilic drug, chlorpromazine. J. Gen. Virol. 2003, 84, 1737–1741. [Google Scholar] [CrossRef]
- Chu, J.J.H.; Leong, P.W.H.; Ng, M.L. Analysis of the endocytic pathway mediating the infectious entry of mosquito-borne flavivirus West Nile into Aedes albopictus mosquito (C6/36) cells. Virology 2006, 349, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, M.N.; Sukumaran, B.; Pal, U.; Agaisse, H.; Murray, J.L.; Hodge, T.W.; Fikrig, E. Rab 5 is required for the cellular entry of dengue and West Nile viruses. J. Virol. 2007, 81, 4881–4885. [Google Scholar] [CrossRef] [Green Version]
- Acosta, E.G.; Castilla, V.; Damonte, E.B. Funcional entry of dengue virus into Aedes albopictus mosquito cells is dependent on clathrin-mediated endocytosis. J. Gen. Virol. 2008, 89 Pt 2, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Acosta, E.G.; Castilla, V.; Damonte, E.B. Alternative infectious entry pathways for dengue virus serotypes into mammalian cells. Cell Microbiol. 2009, 11, 1533–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| CC50 a | CC90 a | IC50 b | IC90 b | SI50 c | |
|---|---|---|---|---|---|
| TIZ | 6.67 ± 0.30 | 2.63 ± 0.11 | 1.38 ± 0.04 | 2.11 ± 0.07 | 4.8 |
| ribavirin | 79692 ± 2390 | 1601 ± 40 | 409 ± 14 | 819 ± 31 | 194.6 |
| (uM) | PFU/mL a | SD b | %VI | Ct | Ct 1:10 | Ct 1:100 | Ct 1:1000 |
|---|---|---|---|---|---|---|---|
| Supernatant | |||||||
| 0 | 1.02 × 106 | 2.40 × 104 | - | 16.2 | 18.5 | 22.4 | 25.6 |
| 1 | 7.97 × 105 | 1.30 × 104 | 21.6 | 16.5 | 18.6 | 22 | 25.6 |
| 1.4 | 4.83 × 105 | 8.50 × 103 | 52.5 | 16.4 | 18.9 | 22.2 | 25.4 |
| 1.8 | 2.70 × 105 | 4.10 × 103 | 73.4 | 17.1 | 19.8 | 23.2 | 26.3 |
| 2.2 | 8.33 × 104 | 6.20 × 103 | 91.8 | 18.1 | 21.3 | 24.2 | 27.5 |
| Monolayers | |||||||
| 0 | 8.68 × 105 | 1.40 × 104 | - | 16.9 | 19.5 | 22.3 | 25.4 |
| 1 | 6.70 × 105 | 1.50 × 104 | 22.8 | 17.3 | 20.1 | 22.5 | 25.3 |
| 1.4 | 3.78 × 105 | 2.00 × 104 | 56.4 | 18.0 | 20.6 | 23.7 | 26.3 |
| 1.8 | 2.10 × 105 | 8.20 × 103 | 75.8 | 18.9 | 21.0 | 24.1 | 27.2 |
| 2.2 | 6.00 × 104 | 8.20 × 103 | 93.1 | 20.5 | 22.1 | 25.1 | 28.6 |
| Fasta Headers | Protein Description | # Peptides | t-Test p-Value | Fold Change | Biological Process | Higher Expression | |
|---|---|---|---|---|---|---|---|
| Control | TIZ | ||||||
| gi|635066390 | dynactin subunit 2 | 3 | 0.012 | 0.2 | Cytoskeleton, intracellular trafficking, secretion, and vesicular transport | x | |
| gi|635142324 | Ras-related protein Rab-9A | 2 | 0.014 | 0.4 | Intracellular trafficking, secretion, and vesicular transport | x | |
| gi|635116140 | Calumenin | 3 | 0.001 | 0.5 | Metabolism | x | |
| gi|635014920 | nucleobindin-2 | 2 | 0.049 | 0.7 | Nuclear protein | x | |
| gi|635016929 | phosphatidylinositol-binding clathrin assembly protein | 3 | 0.013 | 0.7 | Intracellular trafficking, secretion, and vesicular transport | x | |
| gi|635090518 | integrin beta-4 | 2 | 0.030 | 0.8 | Signal transduction mechanisms; cell adhesion | x | |
| gi|635089234 | AP-2 complex subunit beta | 9 | 0.031 | 0.8 | Intracellular trafficking, secretion, and vesicular transport | x | |
| gi|635102142 | galectin-1 | 7 | 0.047 | 0.9 | Cell cycle control, cell division, chromosome partitioning | x | |
| gi|635090268 | septin-9 | 4 | 0.039 | 0.9 | Cell cycle control, cell division, chromosome partitioning | x | |
| gi|635080465 | exportin-1 | 11 | 0.049 | 0.9 | Nuclear protein | x | |
| gi|635015756 | 26S proteasome non-ATPase regulatory subunit 13 | 10 | 0.007 | 1.1 | Postranslational modification; protein turnover | x | |
| gi|635016430 | 40S ribosomal protein S3 | 14 | 0.043 | 1.1 | Cell cycle control, cell division and chromosome partitioning; Translation, ribosomal and biogenesis | x | |
| gi|635023078 | T-complex protein 1 subunit theta | 23 | 0.003 | 1.1 | Postranslational modification; protein turnover; chaperones | x | |
| gi|635067296 | ubiquitin-conjugating enzyme E2 N/N-like | 7 | 0.011 | 1.1 | Postranslational modification; protein turnover; chaperones; | x | |
| gi|635147631 | ADP/ATP translocase 2 | 7 | 0.012 | 1.1 | Cell cycle control, cell division, chromosome partitioning | x | |
| gi|635116374 | aldose reductase | 14 | 0.047 | 1.2 | Metabolism | x | |
| gi|635068162 | sarcoplasmic/endoplasmic reticulum calcium ATPase 2 | 9 | 0.002 | 1.2 | Inorganic ion transpot and metabolism | x | |
| gi|635080331 | actin-related protein 2 | 3 | 0.030 | 1.2 | Cytoskeleton | x | |
| gi|635018272 | coatomer subunit delta | 7 | 0.014 | 1.3 | Intracellular trafficking, secretion, and vesicular transport | x | |
| gi|635104202 | T-complex protein 1 subunit gamma | 20 | 0.018 | 1.3 | Postranslational modification; protein turnover; chaperones | x | |
| gi|635098341 | protein ERGIC-53 | 5 | 0.049 | 1.3 | Intracellular trafficking, secretion, and vesicular transport | x | |
| gi|635074604 | serine palmitoyltransferase 1 | 5 | 0.028 | 1.4 | Metabolism | x | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamoto, K.A.; Blackburn, K.; Goshe, M.B.; Brown, D.T.; Migoswski, E.; Campanhon, I.B.; Moreira, M.F.; Ferreira, D.F.; Soares, M.R. Tizoxanide Antiviral Activity on Dengue Virus Replication. Viruses 2023, 15, 696. https://doi.org/10.3390/v15030696
Yamamoto KA, Blackburn K, Goshe MB, Brown DT, Migoswski E, Campanhon IB, Moreira MF, Ferreira DF, Soares MR. Tizoxanide Antiviral Activity on Dengue Virus Replication. Viruses. 2023; 15(3):696. https://doi.org/10.3390/v15030696
Chicago/Turabian StyleYamamoto, Kristie A., Kevin Blackburn, Michael B. Goshe, Dennis T. Brown, Edimilson Migoswski, Isabele B. Campanhon, Monica F. Moreira, Davis F. Ferreira, and Marcia R. Soares. 2023. "Tizoxanide Antiviral Activity on Dengue Virus Replication" Viruses 15, no. 3: 696. https://doi.org/10.3390/v15030696
APA StyleYamamoto, K. A., Blackburn, K., Goshe, M. B., Brown, D. T., Migoswski, E., Campanhon, I. B., Moreira, M. F., Ferreira, D. F., & Soares, M. R. (2023). Tizoxanide Antiviral Activity on Dengue Virus Replication. Viruses, 15(3), 696. https://doi.org/10.3390/v15030696