
Response to Bovine Viral Diarrhea Virus in Heifers Vaccinated with a Combination of Multivalent Modified Live and Inactivated Viral Vaccines

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. BVDV Strains
2.3. Isolation, Culture, and Preparation of Peripheral Blood Mononuclear Cell (PBMC) for Flow Cytometry
2.4. Antigen and Mitogen Stimulation
2.5. Preparation of PBMC for Flow Cytometry
2.6. Virus Neutralization (VN) Assay
2.7. Data Analysis
3. Results
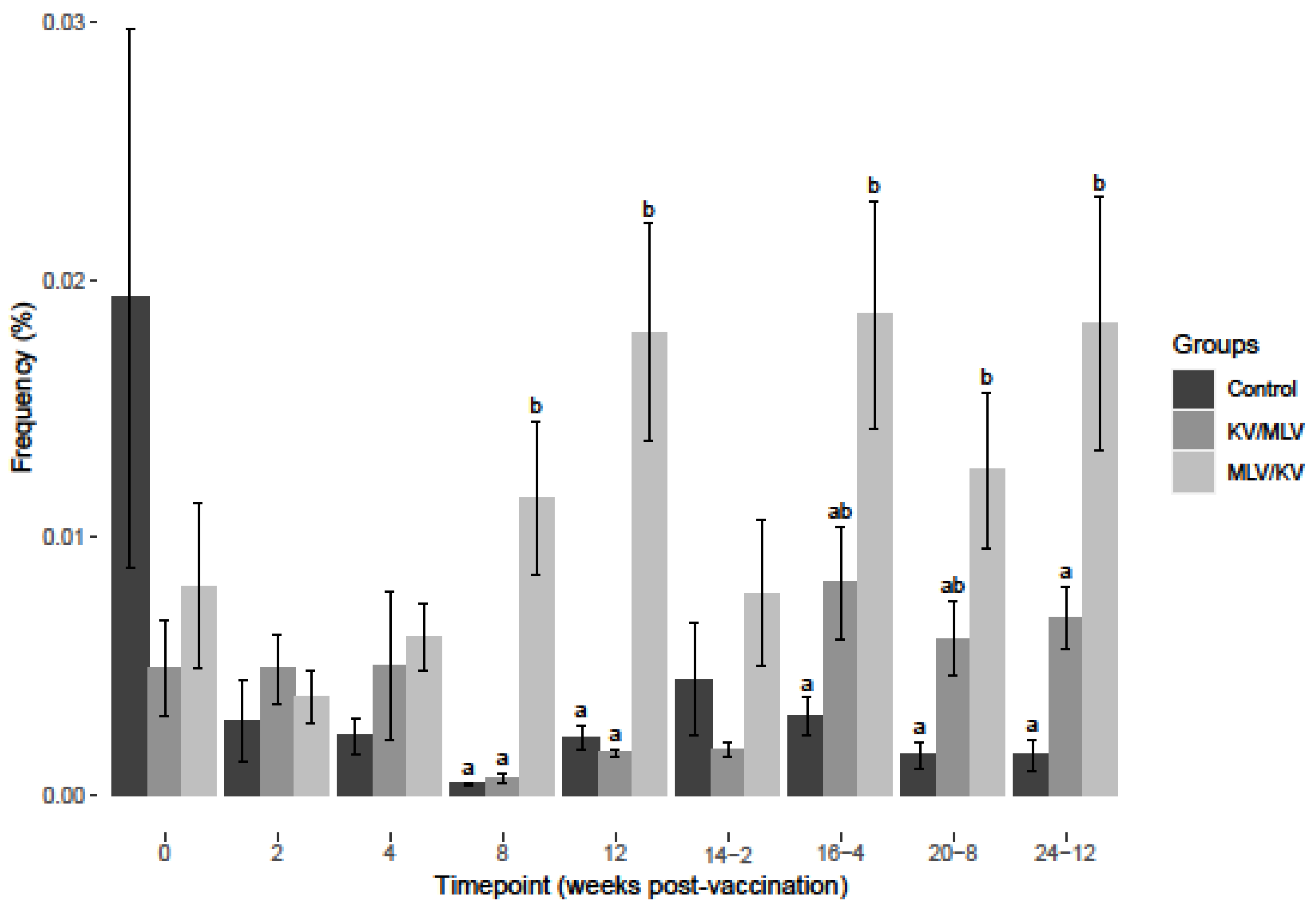
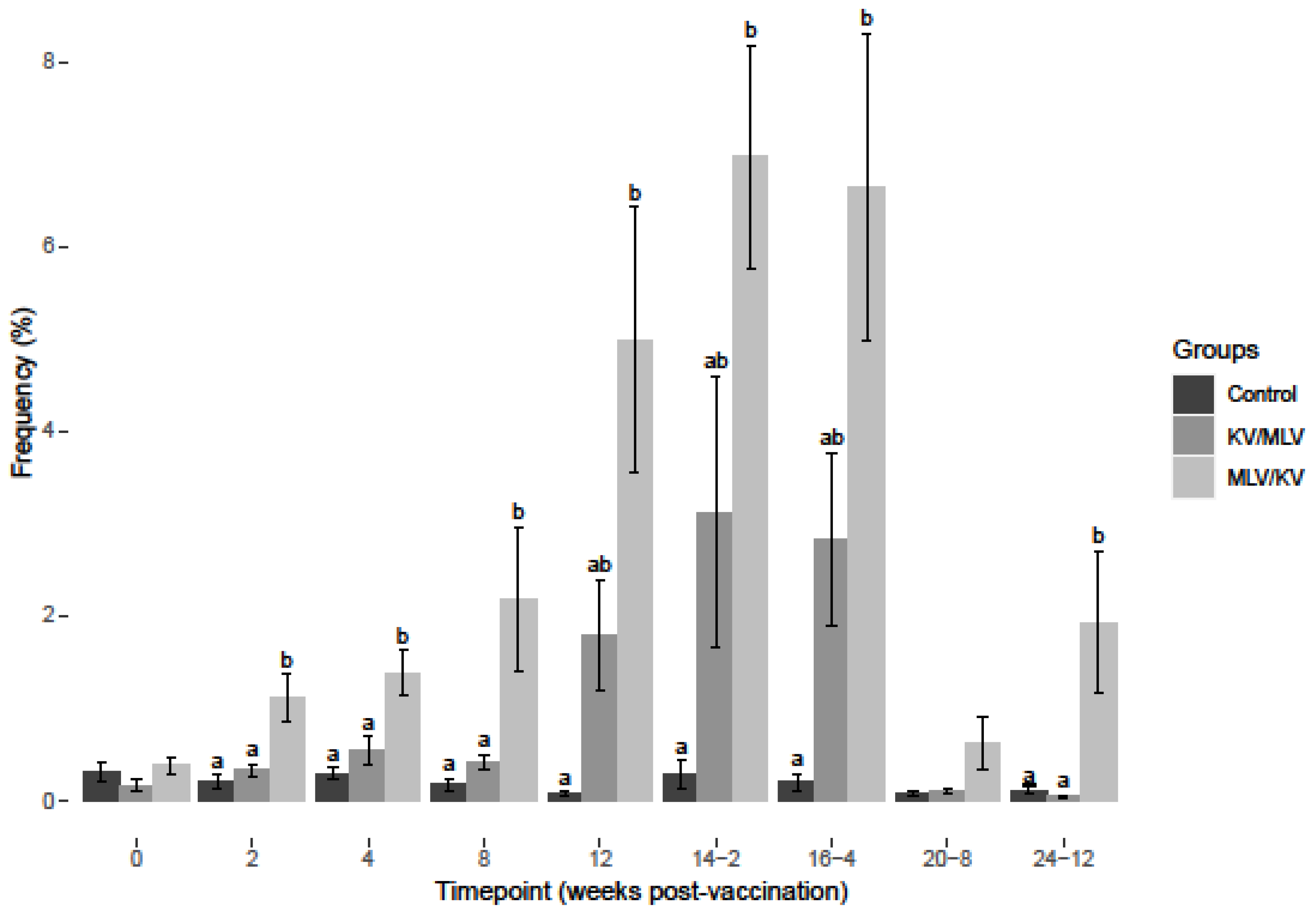
3.1. BVDV-2a Results
3.2. BVDV-1a Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kelling, C.L. Evolution of bovine viral diarrhea virus vaccines. Vet. Clin. Food Anim. Pract. 2004, 20, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Newcomer, B.W.; Givens, D. Diagnosis and control of viral diseases of reproductive importance: Infectious bovine rhinotracheitis and bovine viral diarrhea. Vet. Clin. Food Anim. Pract. 2016, 32, 425–441. [Google Scholar] [CrossRef]
- Newcomer, B.W.; Walz, P.H.; Givens, M.D.; Wilson, A.E. Efficacy of bovine viral diarrhea virus vaccination to prevent reproductive disease: A meta-analysis. Theriogenology 2015, 83, 360–365.e1. [Google Scholar] [CrossRef] [PubMed]
- Brownlie, J.; Clarke, M.C.; Hoopetr, L.B.; Bell, G.D. Protection of the bovine fetus from bovine viral diarrhoea virus by means of a new inactivated vaccine. Vet. Rec. 1995, 137, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Chase, C.C.L.; Fulton, R.W.; O’tole, D.; Gillette, B.; Daly, F.R.; Perry, G.; Clement, T. Bovine herpesvirus 1 modified live virus vaccines for cattle reproduction: Balancing protection with undesired effects. Vet. Microbiol. 2017, 206, 69–77. [Google Scholar] [CrossRef]
- Walz, P.H.; Givens, M.D.; Rodning, S.P.; Riddell, K.P.; Brodersen, B.W.; Scruggs, D.; Short, T.; Grotelueschen, D. Evaluation of reproductive protection against bovine viral diarrhea virus and bovine herpesvirus-1 afforded by annual revaccination with modified-live viral or combination modified-live/killed viral vaccines after primary vaccination with modified-live viral vaccine. Vaccine 2017, 35, 1046–1054. [Google Scholar]
- Royan, G. Comparison of the BVDV, BHV-1, and BRSV anamnestic response to modified-live or inactivated vaccines in calves previously vaccinated with a modified-live virus vaccine. Bov. Pract. 2009, 43, 44–50. [Google Scholar]
- Frey, H.-R.; Eicken, K.; Grummer, B.; Kenklies, S.; Oguzoglu, T.C.; Moennig, V. Foetal protection against bovine virus diarrhoea virus after two-step vaccination. J. Vet. Med. Ser. B 2002, 49, 489–493. [Google Scholar] [CrossRef]
- Dubovi, E.J.; Gröhn, Y.T.; A Brunner, M.; A Hertl, J. Response to modified live and killed multivalent viral vaccine in regularly vaccinated, fresh dairy cows. Vet. Ther. Res. Appl. Vet. Med. 2000, 1, 49–58. [Google Scholar]
- Moennig, V.; Becher, P. Control of bovine viral diarrhea. Pathogens 2018, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Moennig, V.; Eicken, K.; Flebbe, U.; Frey, H.-R.; Grummer, B.; Haas, L.; Greiser-Wilke, I.; Liess, B. Implementation of two-step vaccination in the control of bovine viral diarrhoea (BVD). Prev. Vet. Med. 2005, 72, 109–114. [Google Scholar] [CrossRef]
- Rajput, M.K.; Darweesh, M.F.; Braun, L.J.; Mansour, S.M.; Chase, C.C. Comparative humoral immune response against cytopathic or non-cytopathic bovine viral diarrhea virus infection. Res. Vet. Sci. 2020, 129, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Stevens, E.; Zemmerman, A.D.; Butterbaugh, R.E.; Barling, K.; Scholz, D.; Rhoades, J.; Chas, C.C. The induction of a cell-mediated immune response to bovine viral diarrhea virus with an adjuvanted inactivated vaccine. Vet. Ther. Res. Appl. Vet. Med. 2009, 10, E1–E8. [Google Scholar]
- Platt, R.; Widel, P.W.; Kesl, L.D.; Roth, J.A. Comparison of humoral and cellular immune responses to a pentavalent modified live virus vaccine in three age groups of calves with maternal antibodies, before and after BVDV type 2 challenge. Vaccine 2009, 27, 4508–4519. [Google Scholar] [CrossRef]
- Platt, R.; Kesl, L.; Guidarini, C.; Wang, C.; Roth, J.A. Comparison of humoral and T-cell-mediated immune responses to a single dose of Bovela® live double deleted BVDV vaccine or to a field BVDV strain. Vet. Immunol. Immunopathol. 2017, 187, 20–27. [Google Scholar] [CrossRef]
- Platt, R.; Coutu, C.; Meinert, T.; Roth, J.A. Humoral and T cell-mediated immune responses to bivalent killed bovine viral diarrhea virus vaccine in beef cattle. Vet. Immunol. Immunopathol. 2008, 122, 8–15. [Google Scholar] [CrossRef]
- Falkenberg, S.M.; Dassanayake, R.P.; Neil, D.J.; Walz, P.H.; Casas, E.; Ridpath, F.J.; Roth, J. Measuring CMI responses using the PrimeFlow RNA assay: A new method of evaluating BVDV vaccination response in cattle. Vet. Immunol. Immunopathol. 2020, 221, 110024. [Google Scholar] [CrossRef]
- Falkenberg, S.M.; Dassanayake, R.P.; Terhaar, B.; Ridpath, J.F.; Neill, J.D.; Roth, J.A. Evaluation of Antigenic Comparisons Among BVDV Isolates as it Relates to Humoral and Cell Mediated Responses. Front. Vet. Sci. 2021, 8, 642. [Google Scholar] [CrossRef]
- Mosena, A.C.S.; Falkenberg, S.M.; Ma, H.; Casas, E.; Dassanayake, R.P.; Walz, P.H.; Canal, C.W.; Neill, J.D. Multivariate analysis as a method to evaluate antigenic relationships between BVDV vaccine and field strains. Vaccine 2020, 38, 5764–5772. [Google Scholar] [CrossRef] [PubMed]
- Bauermann, F.V.; Flores, F.E.; Flakenberg, S.M.; Weibelne, J.F.R. Lack of evidence for the presence of emerging HoBi-like viruses in North American fetal bovine serum lots. J. Vet. Diagn. Investig. 2014, 26, 10–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolin, S.R.; Matthews, P.J.; Ridpath, J.F. Methods for detection and frequency of contamination of fetal calf serum with bovine viral diarrhea virus and antibodies against bovine viral diarrhea virus. J. Vet. Diagn. Investig. 1991, 3, 199–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolin, S.R.; Ridpath, J. Assessment of protection from systemic infection or disease afforded by low to intermediate titers of passively acquired neutralizing antibody against bovine viral diarrhea virus in calves. Am. J. Vet. Res. 1995, 56, 755–759. [Google Scholar] [PubMed]
- Ridpath, J.F. Phylogenetic, antigenic and clinical characterization of type 2 BVDV from North America. Vet. Microbiol. 2000, 77, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Riddell, K.P.; Newcomer, B.W.; Neill, J.D.; Falkenberg, S.M.; Cortese, V.S.; Scruggs, D.W.; Short, T.H. Comparison of reproductive protection against bovine viral diarrhea virus provided by multivalent viral vaccines containing inactivated fractions of bovine viral diarrhea virus 1 and 2. Vaccine 2018, 36, 3853–3860. [Google Scholar]
- Gorbalenya, A.E.; Krupovic, M.; Mushegian, A.; Kropinski, A.M.; Siddell, S.G.; Varsani, A.; Adams, M.J.; Davison, A.J.; Dutilh, B.E.; Harrach, B.; et al. The new scope of virus taxonomy: Partitioning the virosphere into 15 hierarchical ranks. Nat. Microbiol. 2020, 5, 668–674. [Google Scholar]
- Van Anne, T.R.; Rinehart, C.L.; Buterbaugh, R.E.; Bauer, M.J.; Young, A.J.; Blaha, M.L.; Klein, A.L.; Chase, C.C.L. Cell-mediated and humoral immune responses to bovine herpesvirus type 1 and bovine viral diarrhea virus in calves following administration of a killed-virus vaccine and bovine herpesvirus type 1 challenge. Am. J. Vet. Res. 2018, 79, 1166–1178. [Google Scholar] [CrossRef]
- Platt, R.; Burdett, W.; Roth, J.A. Roth, Induction of antigen-specific T-cell subset activation to bovine respiratory disease viruses by a modified-live virus vaccine. Am. J. Vet. Res. 2006, 67, 1179–1184. [Google Scholar] [CrossRef]
- Ohmann, H.B.; Babiuk, L. Influence of interferons α11 and γ and of tumour necrosis factor on persistent infection with bovine viral diarrhoea virus in vitro. J. Gen. Virol. 1988, 69, 1399–1403. [Google Scholar] [CrossRef]
- Shannon, I.; White, L.C.; Yang, H.; Nayak, J.L. Differences in Influenza-Specific CD4 T-Cell Mediated Immunity Following Acute Infection Versus Inactivated Vaccination in Children. J. Infect. Dis. 2021, 223, 2164–2173. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, L.; Nordling, D.; Andersson, M.S.; Sheldon, B.; Qvarnstrom, A. Infectious diseases, reproductive effort and the cost of reproduction in birds. Philosophical Transactions of the Royal Society of London. Ser. B Biol. Sci. 1994, 346, 323–331. [Google Scholar]
- Downs, C.; Adelman, J.; Demas, G. Mechanisms and methods in ecoimmunology: Integrating within-organism and between-organism processes. Am. Zool. 2014, 54, 340–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardia, D.R. Tree swallows trade off immune function and reproductive effort differently across their range. Ecology 2005, 86, 2040–2046. [Google Scholar] [CrossRef] [Green Version]
- Oordt, D.A.C.V.; Taff, C.C.; A Ryan, T.; Vitousek, M.N. Timing of Breeding Reveals a Trade-Off between Immune Investment and Life History in Tree Swallows. Integr. Comp. Biol. 2022, 62, 1629–1639. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody and Probes | Cell Marker | Clone | Isotype | Fluorochromes |
|---|---|---|---|---|
| * CD2 | T and NK cells | MUC2A | IgG2a | BV421 |
| CD25 | IL-2 receptor/activation | LCTB2A | IgG3 | BUV395 |
| CD335 | NK cells | ASK1 | IgG1 | BV711 |
| CD4 PrimeFlow probe | T cell subset | AF568 | ||
| CD8α PrimeFlow probe | T cell subset | AF488 | ||
| IFN-γ PrimeFlow probe | IFN-γ mRNA/stimulation | AF750 | ||
| ** BVDV PrimeFlow probe | BVDV viral RNA | AF647 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falkenberg, S.M.; Dassanayake, R.P.; Crawford, L.; Sarlo Davila, K.; Boggiatto, P. Response to Bovine Viral Diarrhea Virus in Heifers Vaccinated with a Combination of Multivalent Modified Live and Inactivated Viral Vaccines. Viruses 2023, 15, 703. https://doi.org/10.3390/v15030703
Falkenberg SM, Dassanayake RP, Crawford L, Sarlo Davila K, Boggiatto P. Response to Bovine Viral Diarrhea Virus in Heifers Vaccinated with a Combination of Multivalent Modified Live and Inactivated Viral Vaccines. Viruses. 2023; 15(3):703. https://doi.org/10.3390/v15030703
Chicago/Turabian StyleFalkenberg, Shollie M., Rohana P. Dassanayake, Lauren Crawford, Kaitlyn Sarlo Davila, and Paola Boggiatto. 2023. "Response to Bovine Viral Diarrhea Virus in Heifers Vaccinated with a Combination of Multivalent Modified Live and Inactivated Viral Vaccines" Viruses 15, no. 3: 703. https://doi.org/10.3390/v15030703
APA StyleFalkenberg, S. M., Dassanayake, R. P., Crawford, L., Sarlo Davila, K., & Boggiatto, P. (2023). Response to Bovine Viral Diarrhea Virus in Heifers Vaccinated with a Combination of Multivalent Modified Live and Inactivated Viral Vaccines. Viruses, 15(3), 703. https://doi.org/10.3390/v15030703