

Open Reading Frame 4 Is Not Essential in the Replication and Infection of Genotype 1 Hepatitis E Virus

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design of ORF4-Defective HEV-1 and the In Vitro Transcription of HEV-1 RNA
2.2. Cell Culture and Transfection
2.3. Infection of Mongolian Gerbils and the Sample Collection
2.4. Detection of HEV-1 RNA
2.5. Detection of Anti-HEV IgG Antibodies
2.6. Liver Enzyme Level
2.7. Expression of ORF4-Related Protein
2.8. Western Blot Analysis
3. Results
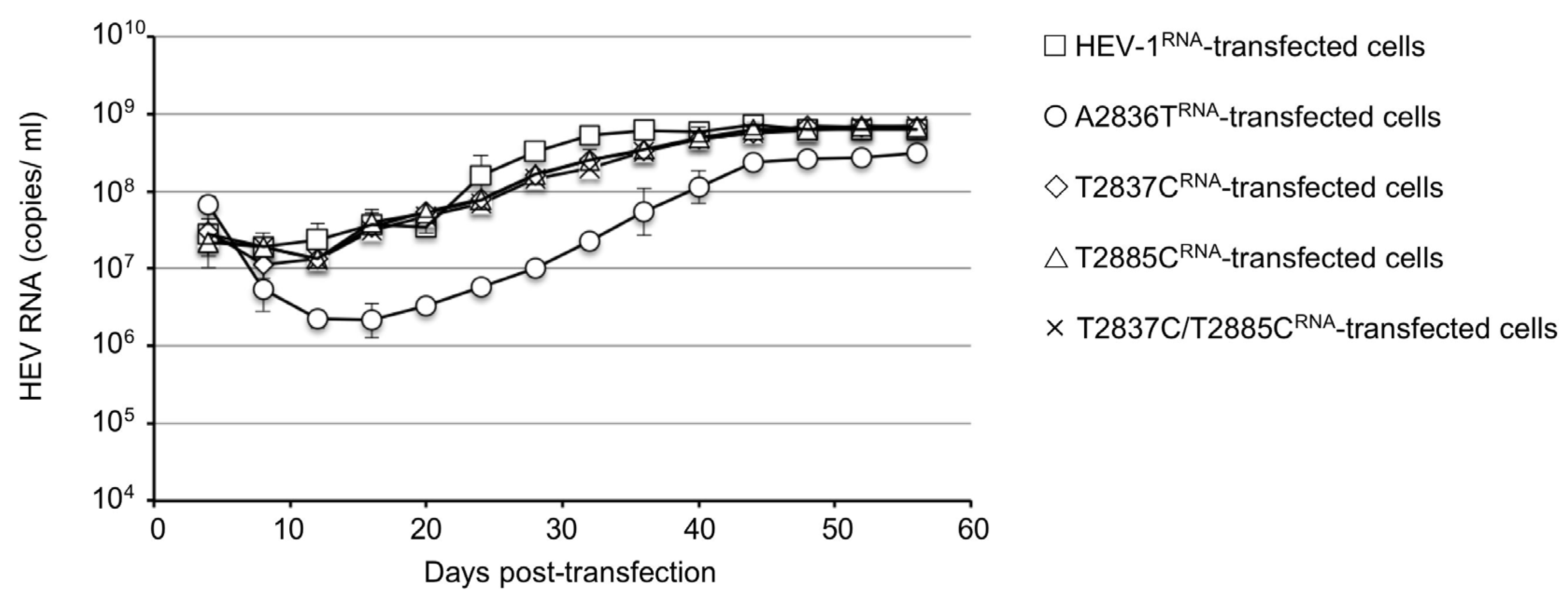
3.1. Generation of Infectious HEV-1 and ORF4-Defective HEV-1s
3.2. Nucleotide Sequence Analyses of ORF4-Defective HEV-1s
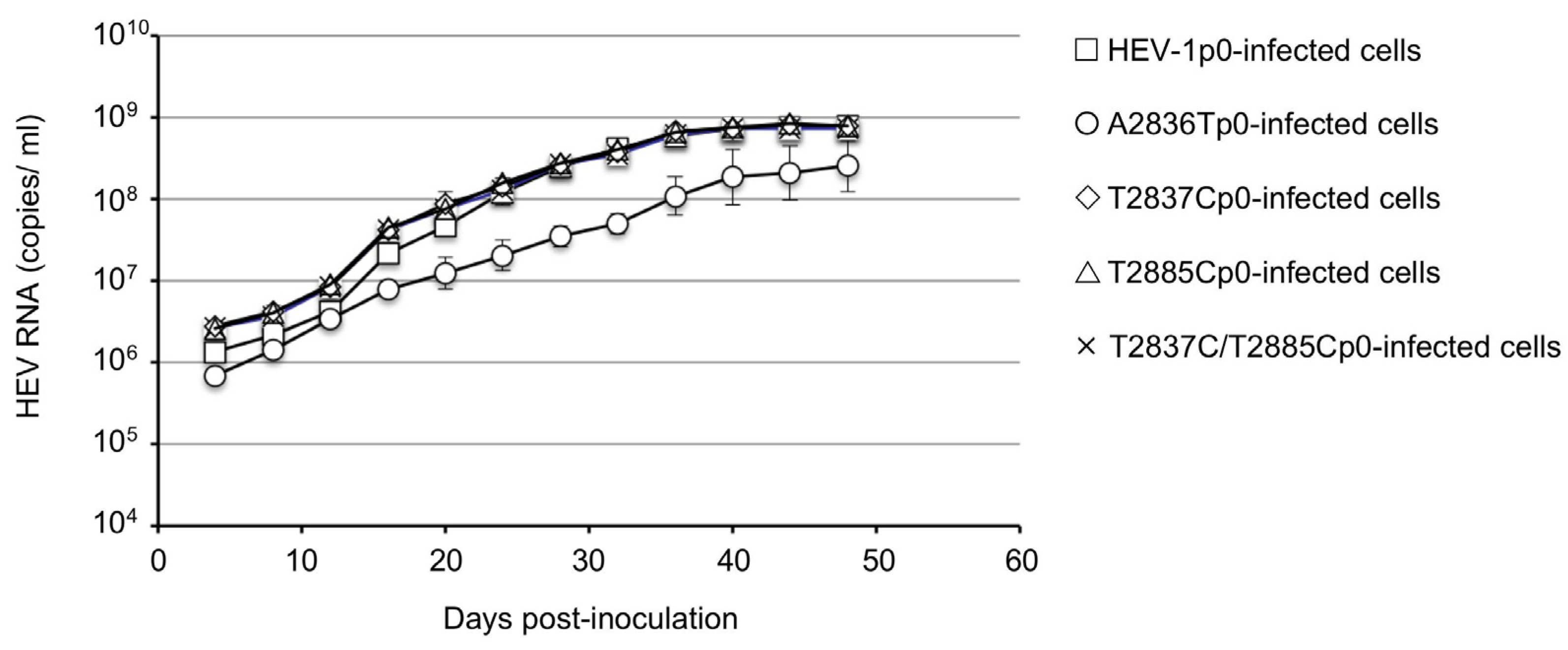
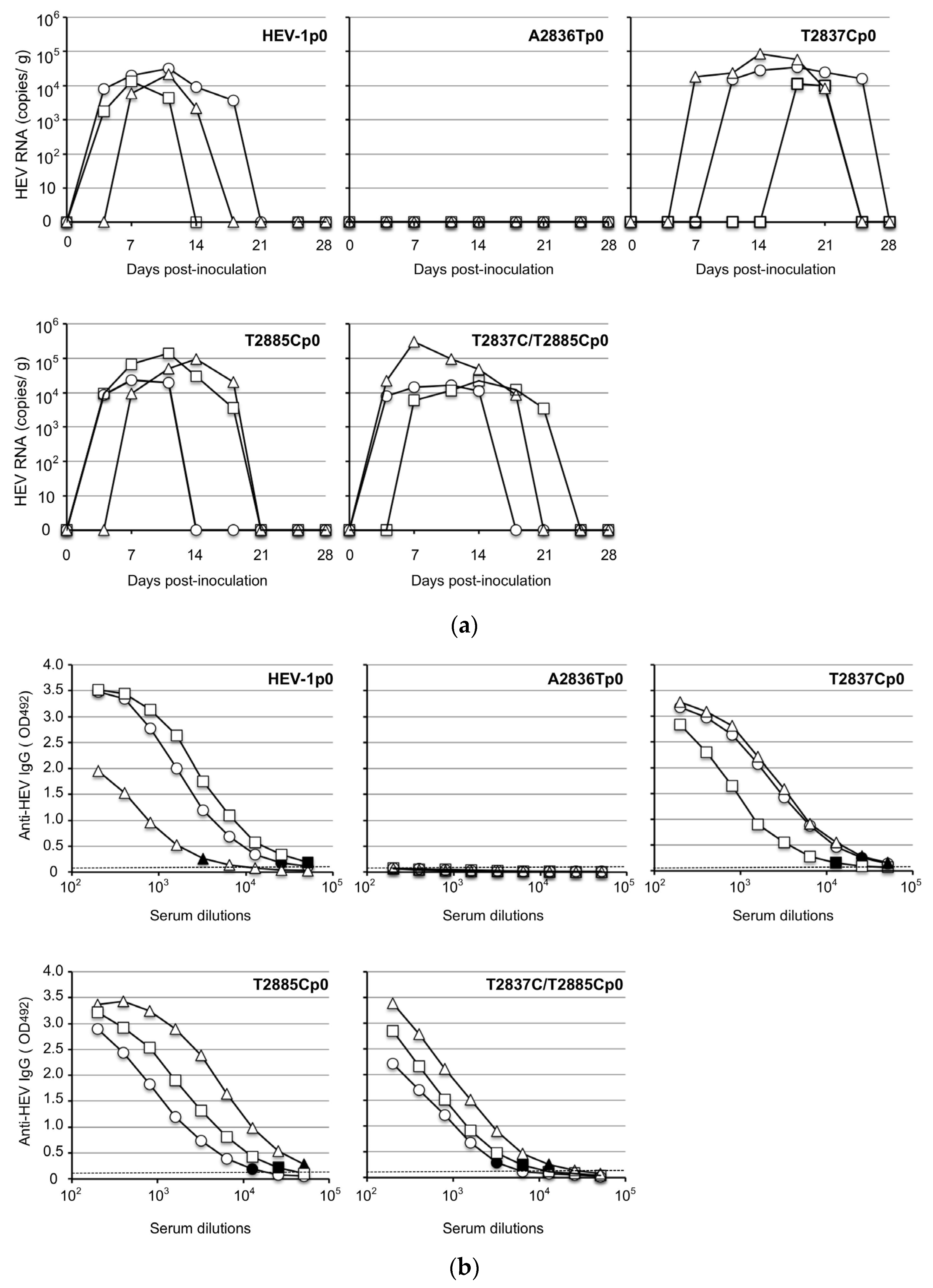
3.3. Infectivity of the ORF4-Defective HEV1s In Vivo
3.4. Detection of Viral Protein in the Wild-type and the ORF4-defective HEV-1s-Infected PLC/PRF/5 Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Purdy, M.A.; Drexler, J.F.; Meng, X.J.; Norder, H.; Okamoto, H.; Van der Poel, W.H.M.; Reuter, G.; de Souza, W.M.; Ulrich, R.G.; Smith, D.B. ICTV Virus Taxonomy Profile: Hepeviridae 2022. J. Gen. Virol. 2022, 103, 001778. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Ambardekar, C.; Lu, Y.; Feng, Z. Distinct Entry Mechanisms for Nonenveloped and Quasi-Enveloped Hepatitis E Viruses. J. Virol. 2016, 90, 4232–4242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.J.; Anderson, D.A.; Arankalle, V.A.; Emerson, S.U.; Harrison, T.J.; Jameel, S.; Okamoto, H. Hepeviridae. In Virus Taxonomy: Ninth Report of the ICTV; King, A.M.A., Michael, J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: London, UK; Academic Press: London, UK, 2012; pp. 1021–1028. [Google Scholar]
- Zhao, C.; Ma, Z.; Harrison, T.J.; Feng, R.; Zhang, C.; Qiao, Z.; Fan, J.; Ma, H.; Li, M.; Song, A.; et al. A novel genotype of hepatitis E virus prevalent among farmed rabbits in China. J. Med. Virol. 2009, 81, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Johne, R.; Heckel, G.; Plenge-Bönig, A.; Kindler, E.; Maresch, C.; Reetz, J.; Schielke, A.; Ulrich, R.G. Novel Hepatitis E Virus Genotype in Norway Rats, Germany. Emerg. Infect. Dis. 2010, 16, 1452–1455. [Google Scholar] [CrossRef] [Green Version]
- Raj, V.S.; Smits, S.L.; Pas, S.D.; Provacia, L.B.; Moorman-Roest, H.; Osterhaus, A.D.; Haagmans, B.L. Novel hepatitis e virus in ferrets, the Netherlands. Emerg. Infect. Dis. 2012, 18, 1369–1370. [Google Scholar] [CrossRef]
- Oliveira-Filho, E.F.; Konig, M.; Thiel, H.J. Genetic variability of HEV isolates: Inconsistencies of current classification. Vet. Microbiol. 2013, 165, 148–154. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Tsang, A.K.; Joseph, M.; Wong, E.Y.; Tang, Y.; Sivakumar, S.; Xie, J.; Bai, R.; et al. New hepatitis E virus genotype in camels, the Middle East. Emerg. Infect. Dis. 2014, 20, 1044–1048. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Cao, K.Y.; Wernery, U.; Schountz, T.; Chiu, T.H.; Tsang, A.K.; Wong, P.C.; Wong, E.Y.; et al. New Hepatitis E Virus Genotype in Bactrian Camels, Xinjiang, China, 2013. Emerg. Infect. Dis. 2016, 22, 2219–2221. [Google Scholar] [CrossRef]
- Sridhar, S.; Yip, C.C.; Wu, S.; Chew, N.F.; Leung, K.H.; Chan, J.F.; Zhao, P.S.; Chan, W.M.; Poon, R.W.; Tsoi, H.W.; et al. Transmission of Rat Hepatitis E Virus Infection to Humans in Hong Kong: A Clinical and Epidemiological Analysis. Hepatology 2021, 73, 10–22. [Google Scholar] [CrossRef]
- Drexler, J.F.; Seelen, A.; Corman, V.M.; Fumie Tateno, A.; Cottontail, V.; Melim Zerbinati, R.; Gloza-Rausch, F.; Klose, S.M.; Adu-Sarkodie, Y.; Oppong, S.K.; et al. Bats Worldwide Carry Hepatitis E Virus-Related Viruses That Form a Putative Novel Genus within the Family Hepeviridae. J. Virol. 2012, 86, 9134–9147. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.F.; Haqshenas, G.; Shivaprasad, H.L.; Guenette, D.K.; Woolcock, P.R.; Larsen, C.T.; Pierson, F.W.; Elvinger, F.; Toth, T.E.; Meng, X.J. Heterogeneity and seroprevalence of a newly identified avian hepatitis e virus from chickens in the United States. J. Clin. Microbiol. 2002, 40, 4197–4202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferri, G.; Vergara, A. Hepatitis E Virus in the Food of Animal Origin: A Review. Foodborne Pathog. Dis. 2021, 18, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Selves, J.; Mansuy, J.M.; Ouezzani, L.; Peron, J.M.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M.; et al. Hepatitis E virus and chronic hepatitis in organ-transplant recipients. N. Engl. J. Med. 2008, 358, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenney, S.P.; Meng, X.J. Hepatitis E Virus Genome Structure and Replication Strategy. Cold Spring Harb. Perspect. Med. 2019, 9, a031724. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, S.; Takahashi, M.; Jirintai; Tanaka, T.; Yamada, K.; Nishizawa, T.; Okamoto, H. A PSAP motif in the ORF3 protein of hepatitis E virus is necessary for virion release from infected cells. J. Gen. Virol. 2011, 92, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Li, T.C.; Ami, Y.; Suzaki, Y.; Yasuda, S.P.; Yoshimatsu, K.; Arikawa, J.; Takeda, N.; Takaji, W. Characterization of full genome of rat hepatitis E virus strain from Vietnam. Emerg. Infect. Dis. 2013, 19, 115–118. [Google Scholar] [CrossRef]
- Li, T.C.; Yang, T.; Ami, Y.; Suzaki, Y.; Shirakura, M.; Kishida, N.; Asanuma, H.; Takeda, N.; Takaji, W. Complete genome of hepatitis E virus from laboratory ferrets. Emerg. Infect. Dis. 2014, 20, 709–712. [Google Scholar] [CrossRef]
- Nair, V.P.; Anang, S.; Subramani, C.; Madhvi, A.; Bakshi, K.; Srivastava, A.; Shalimar; Nayak, B.; Ranjith Kumar, C.T.; Surjit, M. Endoplasmic Reticulum Stress Induced Synthesis of a Novel Viral Factor Mediates Efficient Replication of Genotype-1 Hepatitis E Virus. PLoS Pathog. 2016, 12, e1005521. [Google Scholar] [CrossRef]
- Shafat, Z.; Ahmed, A.; Parvez, M.K.; Parveen, S. Sequence to structure analysis of the ORF4 protein from Hepatitis E virus. Bioinformation 2021, 17, 818–828. [Google Scholar] [CrossRef]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef]
- Li, T.C.; Yamakawa, Y.; Suzuki, K.; Tatsumi, M.; Razak, M.A.; Uchida, T.; Takeda, N.; Miyamura, T. Expression and self-assembly of empty virus-like particles of hepatitis E virus. J. Virol. 1997, 71, 7207–7213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Ami, Y.; Suzaki, Y.; Doan, Y.H.; Muramatsu, M.; Li, T.C. Mongolia Gerbils Are Broadly Susceptible to Hepatitis E Virus. Viruses 2022, 14, 1125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Takeda, N.; Muramatsu, M.; Li, T.C. High Prevalence of Hepatitis E Virus Infection in Imported Cynomolgus Monkeys in Japan. Jpn. J. Infect. Dis. 2019, 72, 429–431. [Google Scholar] [CrossRef] [Green Version]
- Li, T.C.; Suzaki, Y.; Ami, Y.; Dhole, T.N.; Miyamura, T.; Takeda, N. Protection of cynomolgus monkeys against HEV infection by oral administration of recombinant hepatitis E virus-like particles. Vaccine 2004, 22, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Li, T.C.; Zhou, X.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Nakamura, T.; Takeda, N.; Wakita, T. Production of infectious dromedary camel hepatitis E virus by a reverse genetic system: Potential for zoonotic infection. J. Hepatol. 2016, 65, 1104–1111. [Google Scholar] [CrossRef]
- Li, T.C.; Yoshizaki, S.; Yang, T.; Kataoka, M.; Nakamura, T.; Ami, Y.; Yuriko, S.; Takeda, N.; Wakita, T. Production of infectious ferret hepatitis E virus in a human hepatocarcinoma cell line PLC/PRF/5. Virus Res. 2016, 213, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Li, T.C.; Bai, H.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Doan, Y.H.; Takahashi, K.; Mishiro, S.; Takeda, N.; Wakita, T. Genotype 5 Hepatitis E Virus Produced by a Reverse Genetics System Has the Potential for Zoonotic Infection. Hepatol. Commun. 2019, 3, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Li, Y.; Li, Y.; Jin, W.; Duan, S.; Xu, H.; Zhao, Y.; He, Z.; Ami, Y.; Suzaki, Y.; et al. Experimental Cross-Species Transmission of Rat Hepatitis E Virus to Rhesus and Cynomolgus Monkeys. Viruses 2022, 14, 293. [Google Scholar] [CrossRef]
- Yang, F.; Duan, S.; Guo, Y.; Li, Y.; Yoshizaki, S.; Takeda, N.; Wakita, T.; Muramatsu, M.; Zhao, Y.; He, Z.; et al. Current status of hepatitis E virus infection at a rhesus monkey farm in China. Vet. Microbiol. 2019, 230, 244–248. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid | RNA | Nucleotide Position | ORF Positions | Nucleotide Mutations | Amino Acid Positions | Amino Acid Changes |
|---|---|---|---|---|---|---|
| pUC57HEV-1 | HEV-1RNA | |||||
| pUC57A2836T | A2836TRNA | 2836 | ORF4: 1 | ORF4: A to T (Atg to Ttg) | ORF4: 1 | ORF4: Met to Leu |
| ORF1: 2813 | ORF1: A to T (gAt to gTt) | ORF1: 937 | ORF1: Asp to Val | |||
| pUC57T2837C | T2837CRNA | 2837 | ORF4: 2 | ORF4: T to C (aTg to aCg) | ORF4: 1 | ORF4: Met to Thr |
| ORF1: 2814 | ORF1: T to C (gaT to gaC) | ORF1: 937 | ORF1: Non | |||
| pUC57T2885C | T2885CRNA | 2885 | ORF4: 50 | ORF4: T to C (aTg to aCg) | ORF4: 17 | ORF4: Met to Thr |
| ORF1: 2862 | ORF1: T to C (gaT to gaC) | ORF1: 953 | ORF1: Non | |||
| pUC57T2837C/ | T2837C/ T2885CRNA | ORF4: 2 | ORF4: T to C (aTg to aCg) | ORF4: 1 | ORF4: Met to Thr | |
| 2837 | ORF1: 2814 | ORF1: T to C (gaT to gaC) | ORF1: 937 | ORF1: Non | ||
| T2885C | 2885 | ORF4: 50 | ORF4: T to C (aTg to aCg) | ORF4: 17 | ORF4: Met to Thr | |
| ORF1: 2862 | ORF1: T to C (gaT to gaC) | ORF1: 953 | ORF1: Non |
| HEV Strain | Gerbil 1 | Gerbil 2 | Gerbil 3 |
|---|---|---|---|
| HEV-1 | 55 | 49 | 52 |
| T2837C | 57 | 86 | 60 |
| A2836T | 49 | 50 | 54 |
| T2885C | 42 | 56 | 61 |
| T2837C/ T2885C | 41 | 47 | 53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, H.; Ami, Y.; Suzaki, Y.; Doan, Y.H.; Muramatsu, M.; Li, T.-C. Open Reading Frame 4 Is Not Essential in the Replication and Infection of Genotype 1 Hepatitis E Virus. Viruses 2023, 15, 784. https://doi.org/10.3390/v15030784
Bai H, Ami Y, Suzaki Y, Doan YH, Muramatsu M, Li T-C. Open Reading Frame 4 Is Not Essential in the Replication and Infection of Genotype 1 Hepatitis E Virus. Viruses. 2023; 15(3):784. https://doi.org/10.3390/v15030784
Chicago/Turabian StyleBai, Huimin, Yasushi Ami, Yuriko Suzaki, Yen Hai Doan, Masamichi Muramatsu, and Tian-Cheng Li. 2023. "Open Reading Frame 4 Is Not Essential in the Replication and Infection of Genotype 1 Hepatitis E Virus" Viruses 15, no. 3: 784. https://doi.org/10.3390/v15030784
APA StyleBai, H., Ami, Y., Suzaki, Y., Doan, Y. H., Muramatsu, M., & Li, T. -C. (2023). Open Reading Frame 4 Is Not Essential in the Replication and Infection of Genotype 1 Hepatitis E Virus. Viruses, 15(3), 784. https://doi.org/10.3390/v15030784