
Role of Histamine and Related Signaling in Kaposi’s Sarcoma-Associated Herpesvirus Pathogenesis and Oncogenesis




 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Reagents and Infection Protocols
2.2. Cell Proliferation and Soft Agar Assays
2.3. RT-qPCR
2.4. Western Blot
2.5. KS Tumor Tissues from HIV+ Patients and Immunohistochemistry
2.6. PEL Xenograft Models
2.7. Statistical Analysis
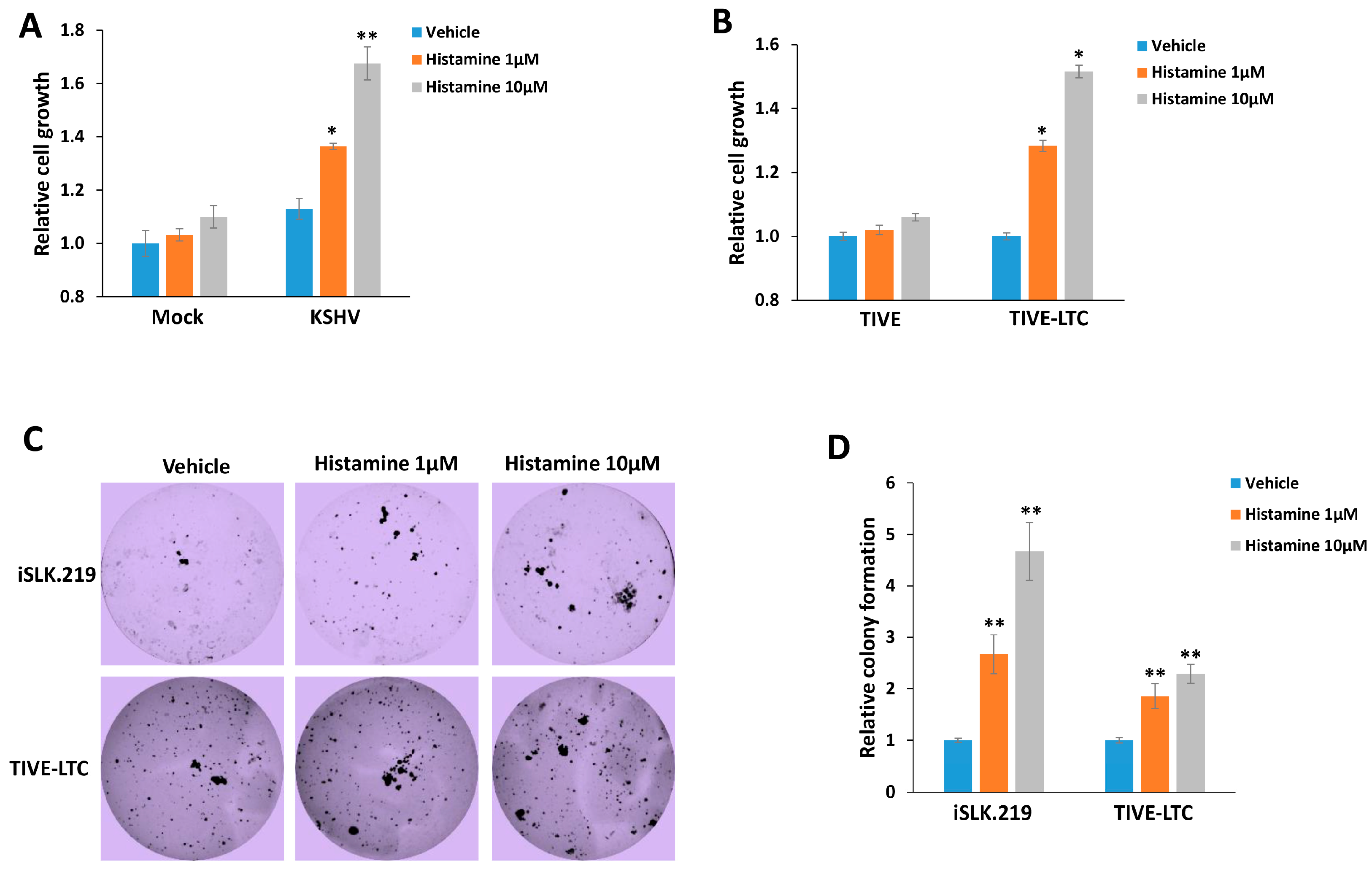
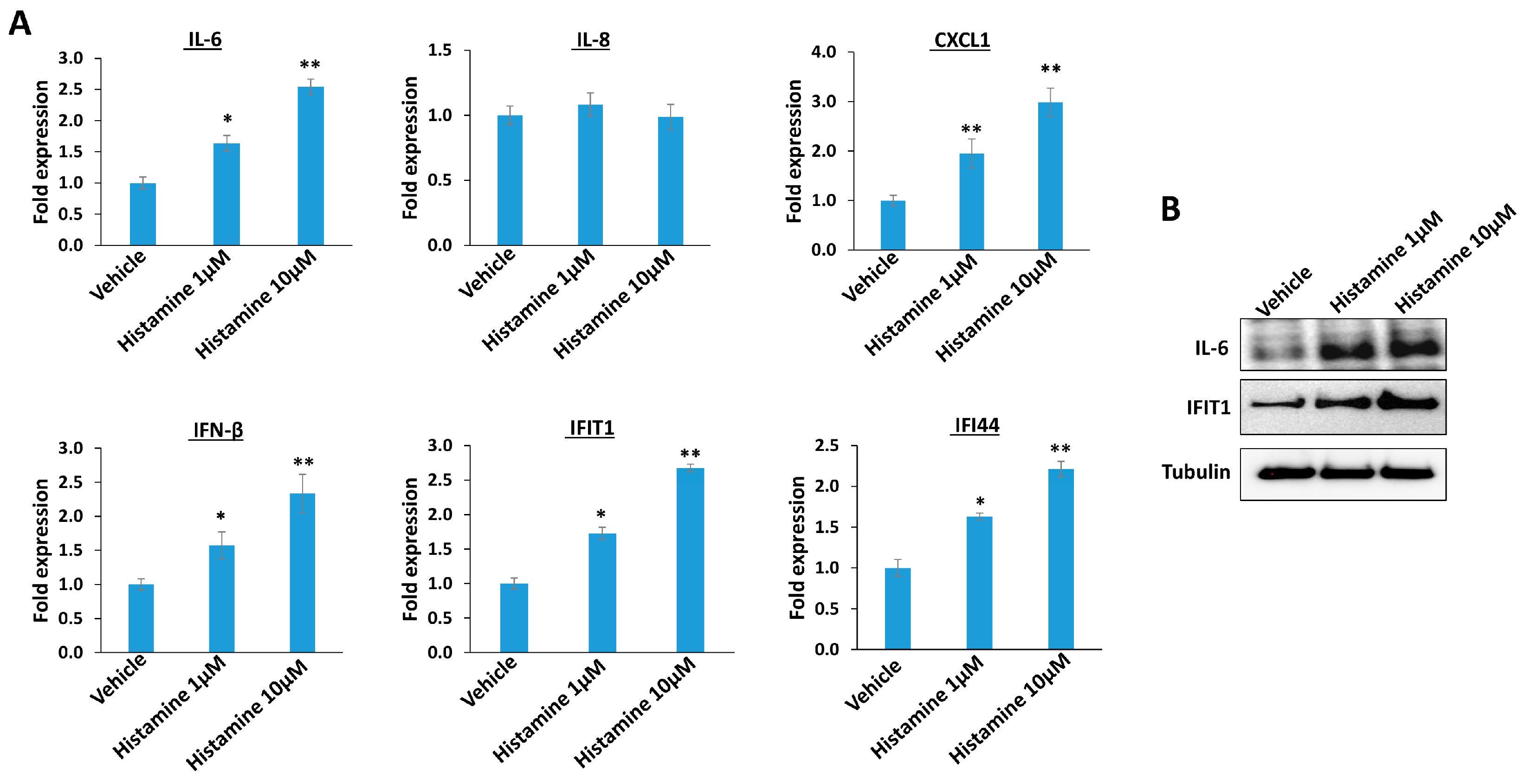
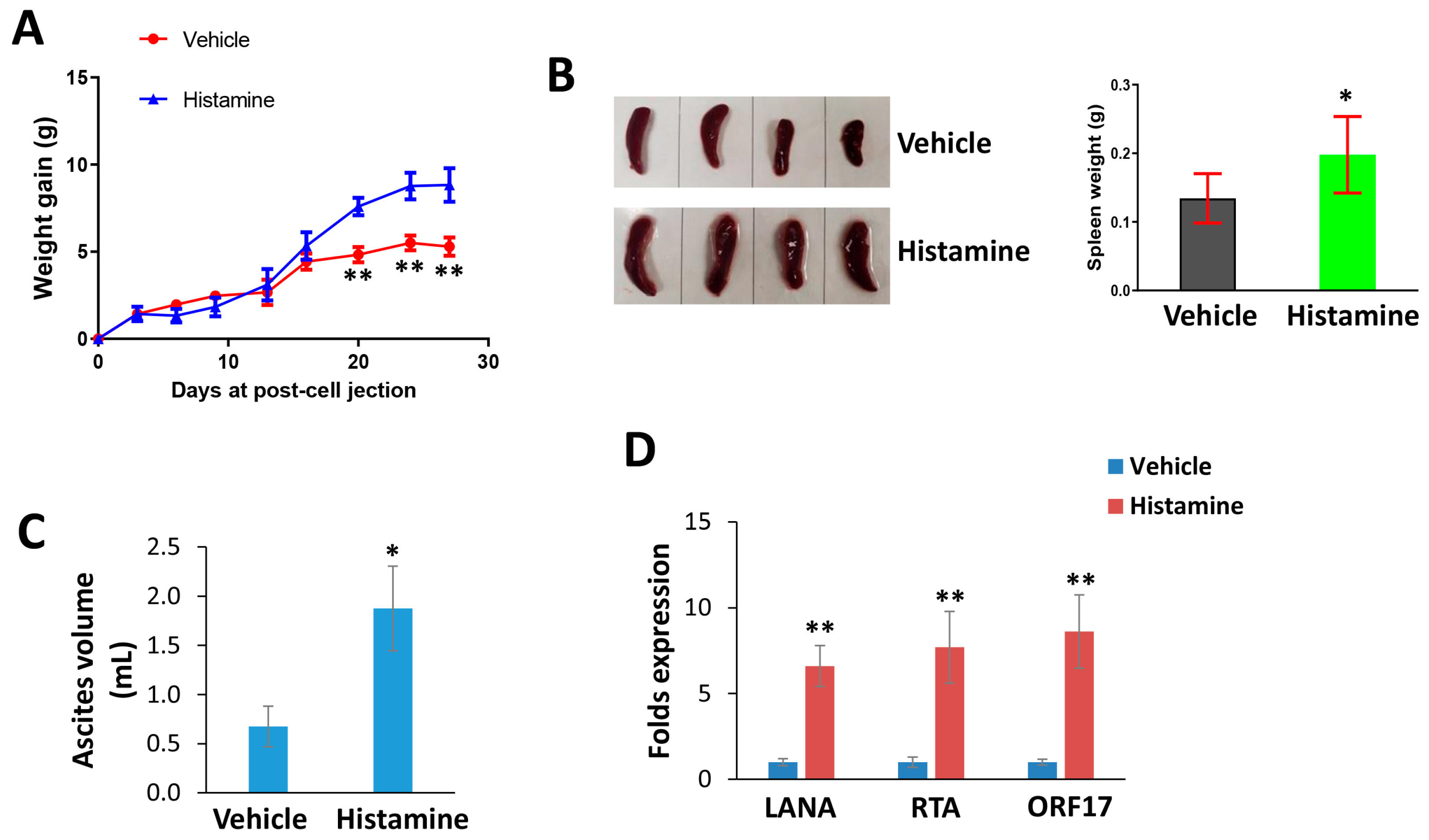
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chang, Y.; Cesarman, E.; Pessin, M.S.; Lee, F.; Culpepper, J.; Knowles, D.M.; Moore, P.S. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi’s sarcoma. Science 1994, 266, 1865–1869. [Google Scholar] [CrossRef]
- Dupin, N. Update on oncogenesis and therapy for Kaposi sarcoma. Curr. Opin. Oncol. 2020, 32, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, F.J.; Hoffman, L.J.; Liegey-Dougall, A. Reactivation of and Primary Infection with Human Herpesvirus 8 among Solid-Organ Transplant Recipients. J. Infect. Dis. 2002, 185, 1238–1243. [Google Scholar] [CrossRef] [PubMed]
- Luppi, M.; Barozzi, P.; Santagostino, G.; Trovato, R.; Schulz, T.F.; Marasca, R.; Bottalico, D.; Bignardi, L.; Torelli, G. Molecular evidence of organ-related transmission of Kaposi sarcoma-associated herpesvirus or human herpesvirus-8 in transplant patients. Blood 2000, 96, 3279–3281. [Google Scholar] [CrossRef]
- Ariza-Heredia, E.J.; Razonable, R.R. Human Herpes Virus 8 in Solid Organ Transplantation. Transplantation 2011, 92, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Faustino-Rocha, A.I.; Gama, A.; Neuparth, M.J.; Oliveira, P.A.; Ferreira, R.; Ginja, M. Mast Cells in Mammary Carcinogenesis: Host or Tumor Supporters? Anticancer Res. 2017, 37, 1013–1021. [Google Scholar]
- Xie, H.; He, S.H. Roles of histamine and its receptors in allergic and inflammatory bowel diseases. World J. Gastroenterol. 2005, 11, 2851–2857. [Google Scholar] [CrossRef]
- Ayers, L.W.; Barbachano-Guerrero, A.; McAllister, S.C.; Ritchie, J.A.; Asiago-Reddy, E.; Bartlett, L.C.; Cesarman, E.; Wang, D.; Rochford, R.; Martin, J.N.; et al. Mast Cell Activation and KSHV Infection in Kaposi Sarcoma. Clin. Cancer Res. 2018, 24, 5085–5097. [Google Scholar] [CrossRef]
- Chen, J.; Dai, L.; Goldstein, A.; Zhang, H.; Tang, W.; Forrest, J.C.; Post, S.R.; Chen, X.; Qin, Z. Identification of new antiviral agents against Kaposi’s sarcoma-associated herpesvirus (KSHV) by high-throughput drug screening reveals the role of histamine-related signaling in promoting viral lytic reactivation. PLoS Pathog. 2019, 15, e1008156. [Google Scholar] [CrossRef]
- Myoung, J.; Ganem, D. Generation of a doxycycline-inducible KSHV producer cell line of endothelial origin: Maintenance of tight latency with efficient reactivation upon induction. J. Virol. Methods 2011, 174, 12–21. [Google Scholar] [CrossRef]
- An, F.Q.; Folarin, H.M.; Compitello, N.; Roth, J.; Gerson, S.L.; McCrae, K.R.; Fakhari, F.D.; Dittmer, D.P.; Renne, R. Long-term-infected telomerase-immortalized endothelial cells: A model for Kaposi’s sarcoma-associated herpesvirus latency in vitro and in vivo. J. Virol. 2006, 80, 4833–4846. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Freitas, E.; Sullivan, R.; Mohan, S.; Bacelieri, R.; Branch, D.; Romano, M.; Kearney, P.; Oates, J.; Plaisance, K.; et al. Upregulation of xCT by KSHV-Encoded microRNAs Facilitates KSHV Dissemination and Persistence in an Environment of Oxidative Stress. PLoS Pathog. 2010, 6, e1000742. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Del Valle, L.; Miley, W.; Whitby, D.; Ochoa, A.C.; Flemington, E.K.; Qin, Z. Transactivation of human endogenous retrovirus K (HERV-K) by KSHV promotes Kaposi’s sarcoma development. Oncogene 2018, 37, 4534–4545. [Google Scholar] [CrossRef]
- Dai, L.; Trillo-Tinoco, J.; Cao, Y.; Bonstaff, K.; Doyle, L.; Del Valle, L.; Whitby, D.; Parsons, C.; Reiss, K.; Zabaleta, J.; et al. Targeting HGF/c-MET induces cell cycle arrest, DNA damage, and apoptosis for primary effusion lymphoma. Blood 2015, 126, 2821–2831. [Google Scholar] [CrossRef]
- Dai, L.; Cao, Y.; Jiang, W.; Zabaleta, J.; Liu, Z.; Qiao, J.; Qin, Z. KSHV co-infection down-regulates HPV16 E6 and E7 from cervical cancer cells. Oncotarget 2017, 8, 35792–35803. [Google Scholar] [CrossRef]
- Barrett, L.; Chen, J.; Dai, L.; Plaisance-Bonstaff, K.; Del Valle, L.; Qin, Z. Role of Interleukin-1 Family Members and Signaling Pathways in KSHV Pathogenesis. Front. Cell. Infect. Microbiol. 2020, 10, 587929. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Lan, K. Activation and counteraction of antiviral innate immunity by KSHV: An update. Sci. Bull. 2018, 63, 1223–1234. [Google Scholar] [CrossRef]
- Dong, H.; Zhang, W.; Zeng, X.; Hu, G.; Zhang, H.; He, S.; Zhang, S. Histamine Induces Upregulated Expression of Histamine Receptors and Increases Release of Inflammatory Mediators from Microglia. Mol. Neurobiol. 2014, 49, 1487–1500. [Google Scholar] [CrossRef]
- Bakker, R.A.; Schoonus, S.B.; Smit, M.J.; Timmerman, H.; Leurs, R. Histamine H(1)-receptor activation of nuclear factor-kappa B: Roles for G beta gamma- and G alpha(q/11)-subunits in constitutive and agonist-mediated signaling. Mol. Pharmacol. 2001, 60, 1133–1142. [Google Scholar] [CrossRef]
- Nizamutdinova, I.T.; Dusio, G.F.; Gasheva, O.Y.; Skoog, H.; Tobin, R.; Peddaboina, C.; Meininger, C.J.; Zawieja, D.C.; Newell-Rogers, M.K.; Gashev, A.A. Mast cells and histamine are triggering the NF-kappaB-mediated reactions of adult and aged perilymphatic mesenteric tissues to acute inflammation. Aging 2016, 8, 3065–3090. [Google Scholar] [CrossRef]
- Seifert, R.; Strasser, A.; Schneider, E.H.; Neumann, D.; Dove, S.; Buschauer, A. Molecular and cellular analysis of human histamine receptor subtypes. Trends Pharmacol. Sci. 2013, 34, 33–58. [Google Scholar] [CrossRef] [PubMed]
- Cricco, G.P.; Davio, C.A.; Martin, G.; Engel, N.; Fitzsimons, C.P.; Bergoc, R.M.; Rivera, E.S. Histamine as an autocrine growth factor in experimental mammary carcinomas. Agents Actions 1994, 43, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Stoyanov, E.; Uddin, M.; Mankuta, D.; Dubinett, S.M.; Levi-Schaffer, F. Mast cells and histamine enhance the proliferation of non-small cell lung cancer cells. Lung Cancer 2012, 75, 38–44. [Google Scholar] [CrossRef]
- Aoki, Y.; Yarchoan, R.; Braun, J.; Iwamoto, A.; Tosato, G. Viral and cellular cytokines in AIDS-related malignant lymphomatous effusions. Blood 2000, 96, 1599–1601. [Google Scholar] [CrossRef] [PubMed]
- Oksenhendler, E.; Carcelain, G.; Aoki, Y.; Boulanger, E.; Maillard, A.; Clauvel, J.P.; Agbalika, F. High levels of human herpesvirus 8 viral load, human interleukin-6, interleukin-10, and C reactive protein correlate with exacerbation of multicentric castleman disease in HIV-infected patients. Blood 2000, 96, 2069–2073. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Lee, J.; Kang, S.K.; Wirth, D.; Yoo, S.M.; Park, C.; Lee, M.S. CXCL1 confers a survival advantage in Kaposi’s sarcoma-associated herpesvirus-infected human endothelial cells through STAT3 phosphorylation. J. Med. Virol. 2023, 95. [Google Scholar] [CrossRef]
- Gonzalez-Perez, A.C.; Stempel, M.; Chan, B.; Brinkmann, M.M. One Step Ahead: Herpesviruses Light the Way to Understanding Interferon-Stimulated Genes (ISGs). Front. Microbiol. 2020, 11, 124. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Song, J.; Plaisance-Bonstaff, K.; Mu, S.; Post, S.R.; Dai, L.; Qin, Z. Role of Histamine and Related Signaling in Kaposi’s Sarcoma-Associated Herpesvirus Pathogenesis and Oncogenesis. Viruses 2023, 15, 1011. https://doi.org/10.3390/v15041011
Chen J, Song J, Plaisance-Bonstaff K, Mu S, Post SR, Dai L, Qin Z. Role of Histamine and Related Signaling in Kaposi’s Sarcoma-Associated Herpesvirus Pathogenesis and Oncogenesis. Viruses. 2023; 15(4):1011. https://doi.org/10.3390/v15041011
Chicago/Turabian StyleChen, Jungang, Jiao Song, Karlie Plaisance-Bonstaff, Shengyu Mu, Steven R. Post, Lu Dai, and Zhiqiang Qin. 2023. "Role of Histamine and Related Signaling in Kaposi’s Sarcoma-Associated Herpesvirus Pathogenesis and Oncogenesis" Viruses 15, no. 4: 1011. https://doi.org/10.3390/v15041011
APA StyleChen, J., Song, J., Plaisance-Bonstaff, K., Mu, S., Post, S. R., Dai, L., & Qin, Z. (2023). Role of Histamine and Related Signaling in Kaposi’s Sarcoma-Associated Herpesvirus Pathogenesis and Oncogenesis. Viruses, 15(4), 1011. https://doi.org/10.3390/v15041011