
The Lytic Activity of Bacteriophage ZCSE9 against Salmonella enterica and Its Synergistic Effects with Kanamycin

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Cultures and Preparations
2.2. Isolation, Purification, and Amplification of the Phage
2.3. Host Range and Lytic Profile
2.4. Transmission Electron Microscopy (TEM)
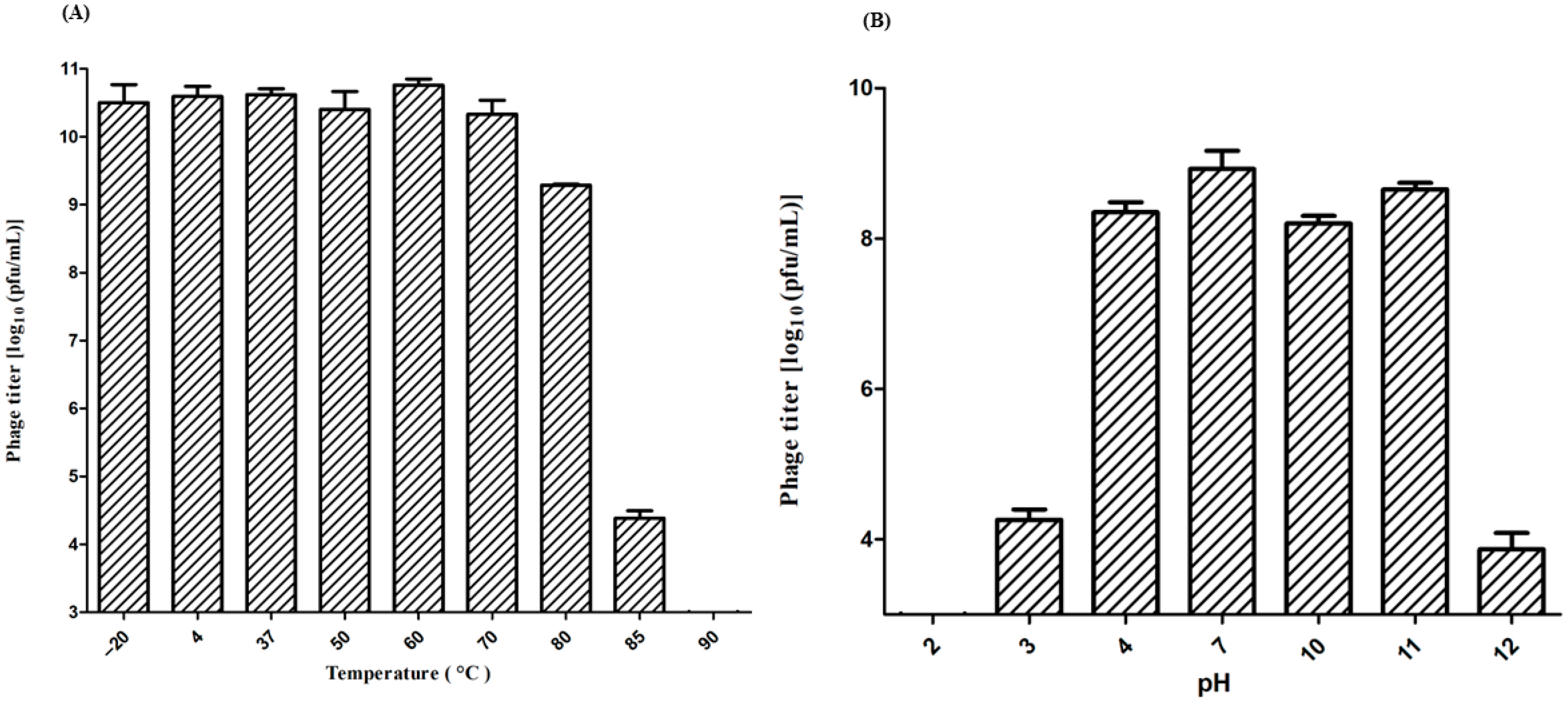
2.5. Physicochemical Stability
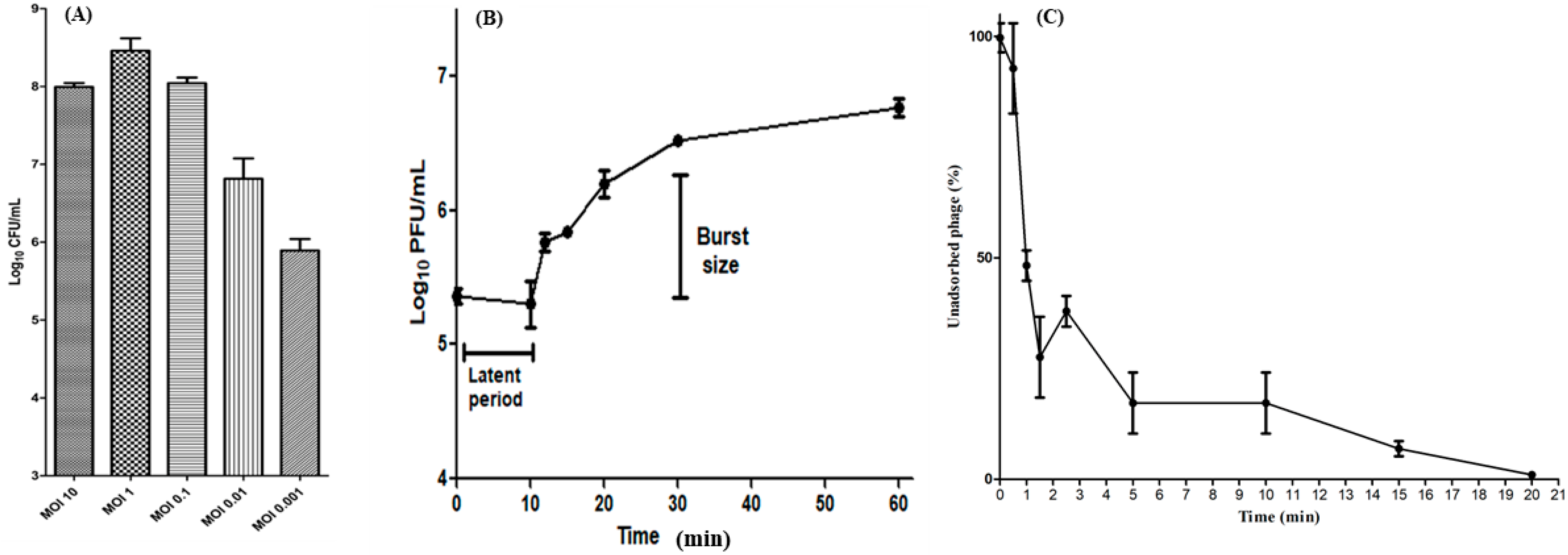
2.6. Determination of Optimal MOI
2.7. One-Step Growth Curve
2.8. Adsorption Rate Assay
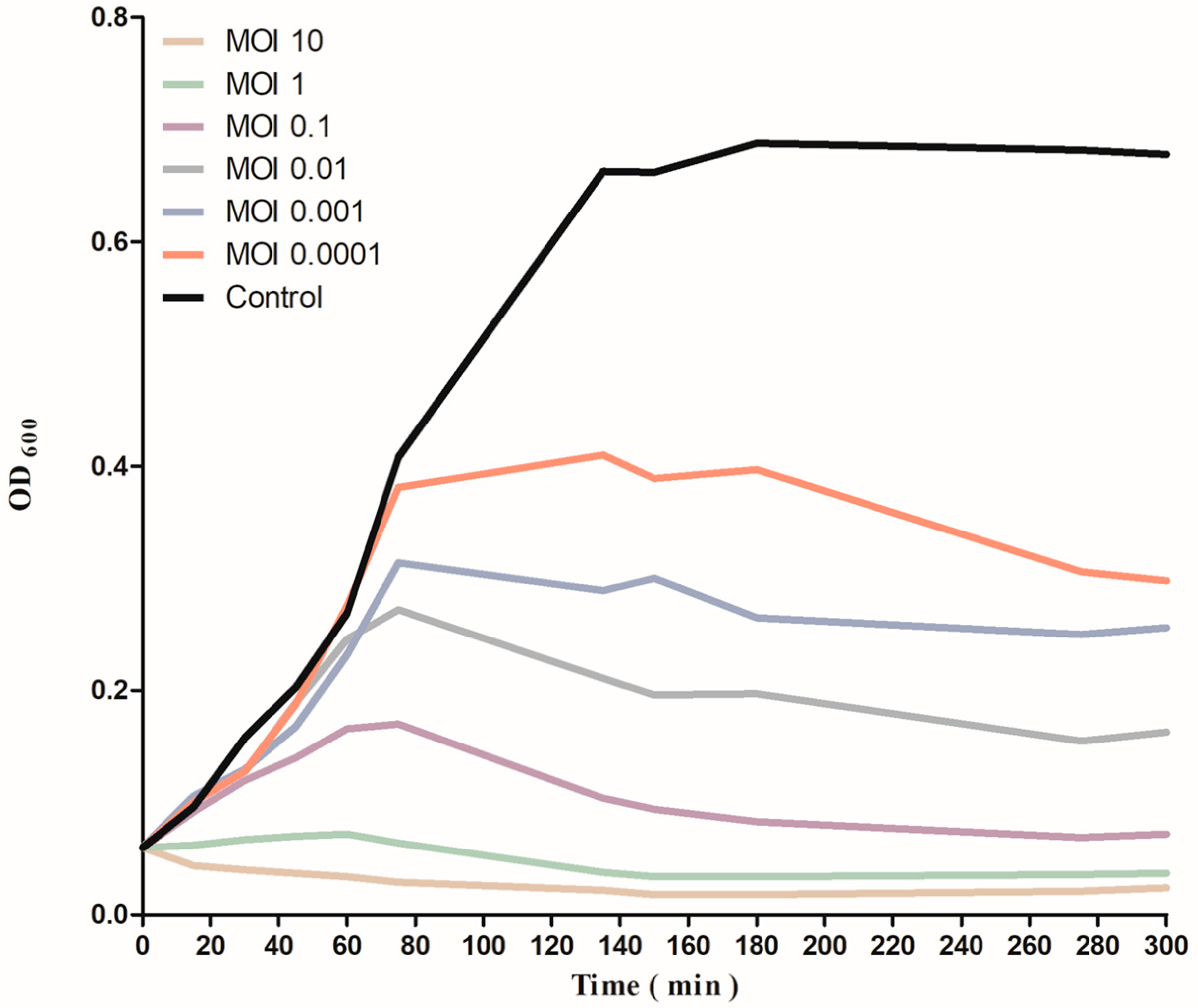
2.9. Time Killing Curve
2.10. Phage DNA Extraction and Genome Sequencing
2.11. Genome Analysis
2.11.1. Assembly and Annotation
2.11.2. Phylogenetic Analysis
2.12. Phage-Antibiotic Synergy
3. Results and Discussion
3.1. Host Range
3.2. Morphological Characteristics of ZCSE9
3.3. Physicochemical Stability of Phage
3.4. Optimal MOI, One-Step Growth Curve, and Adsorption Rate Assay
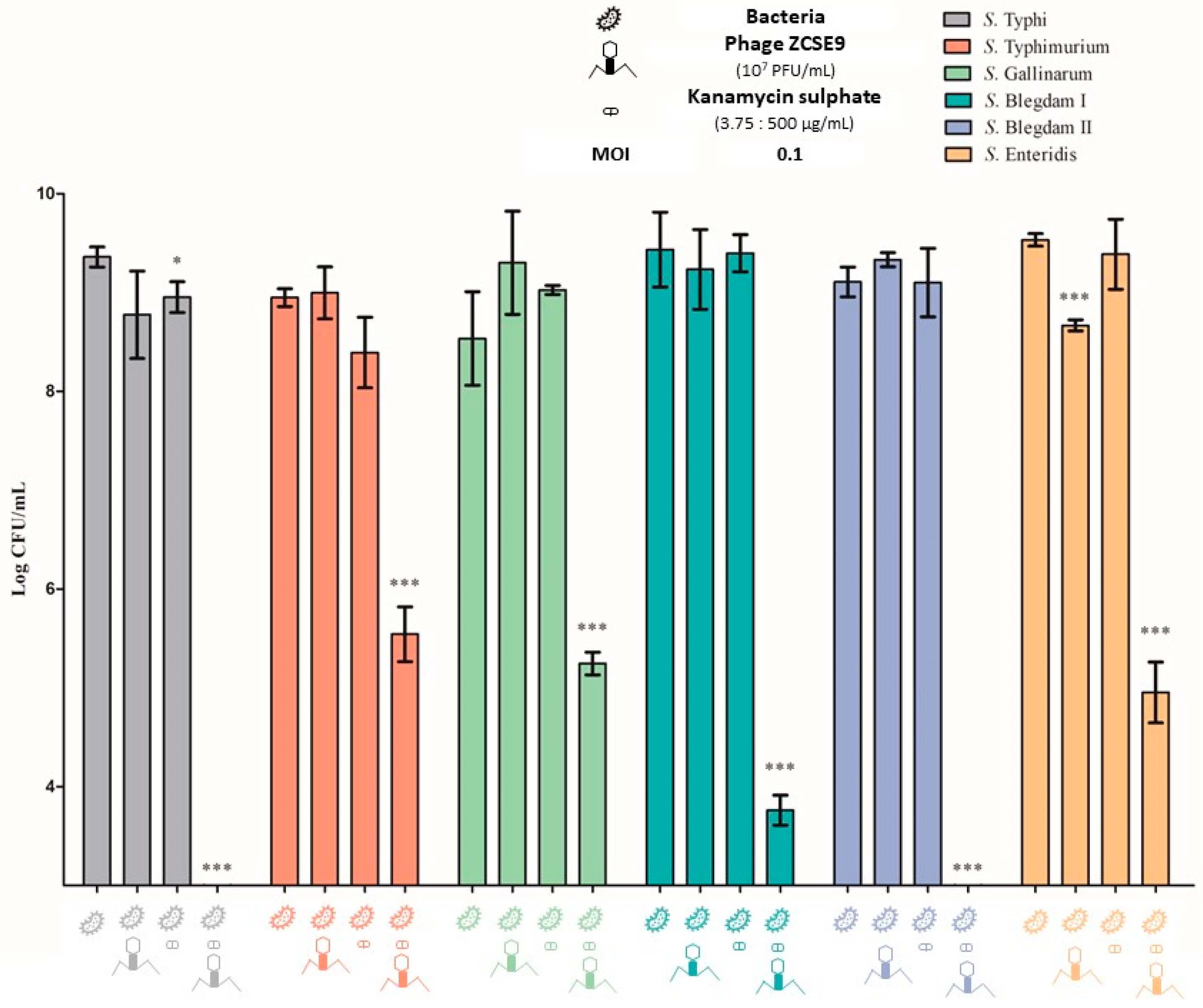
3.5. Lytic Activity
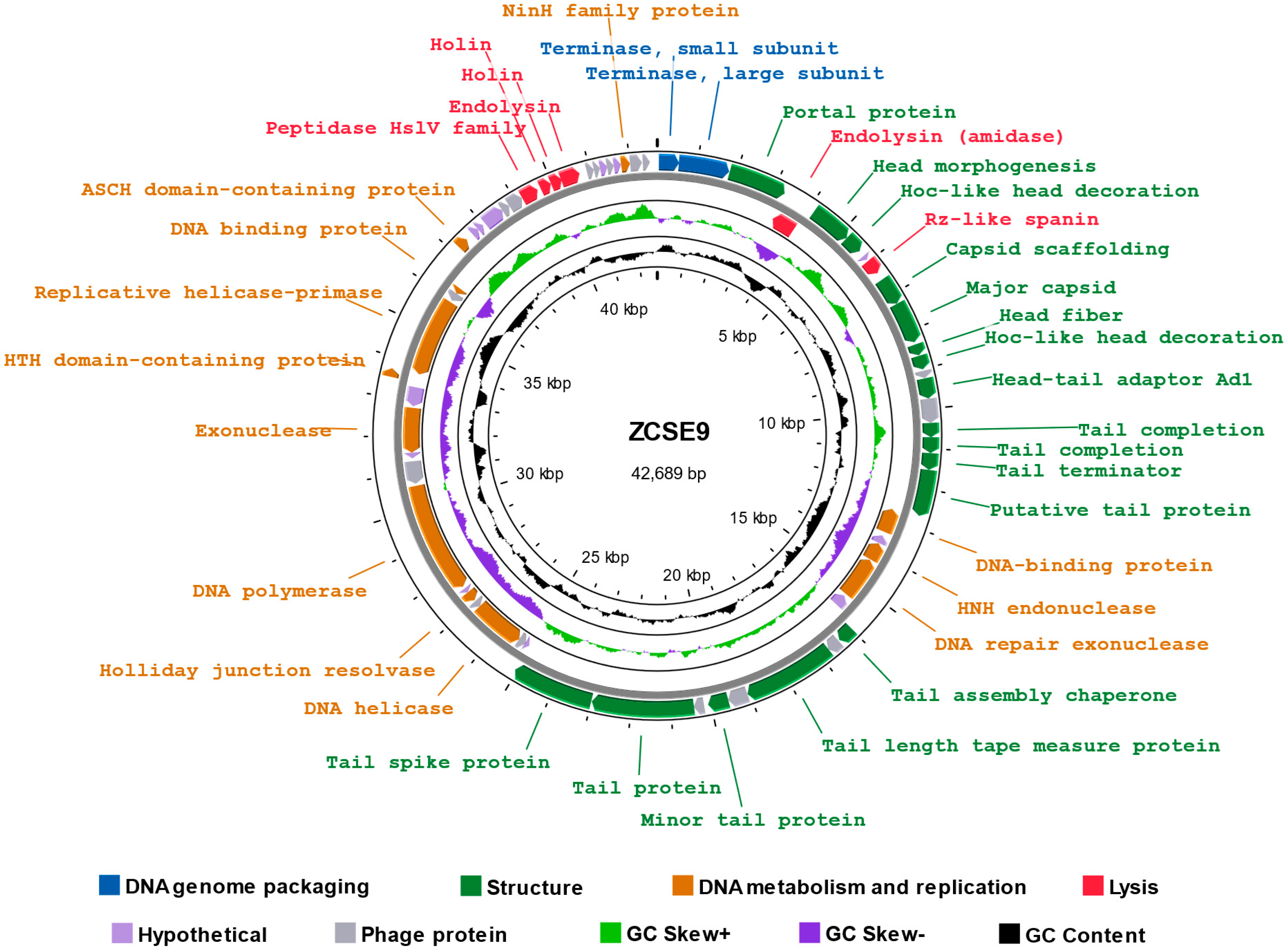
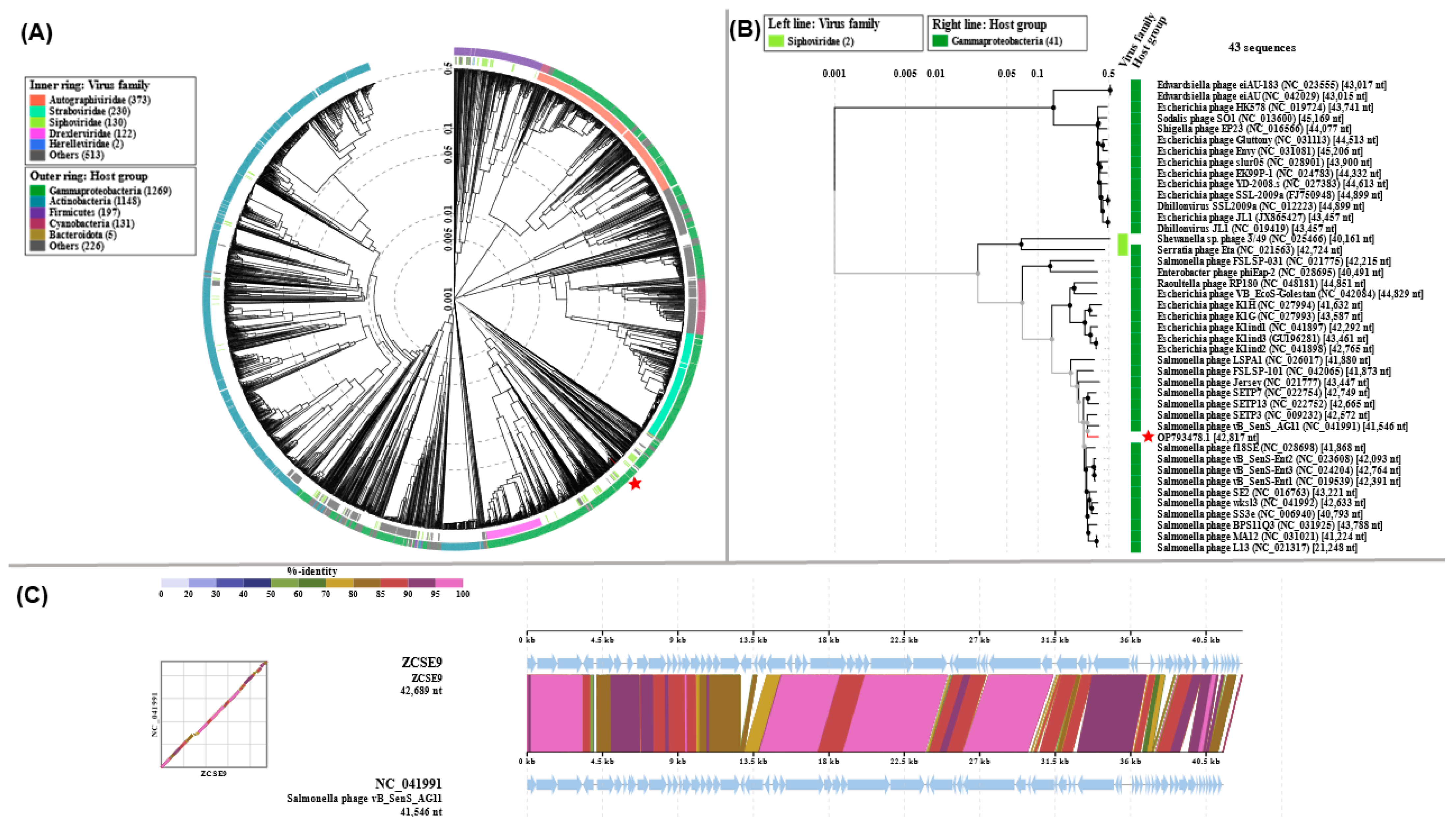
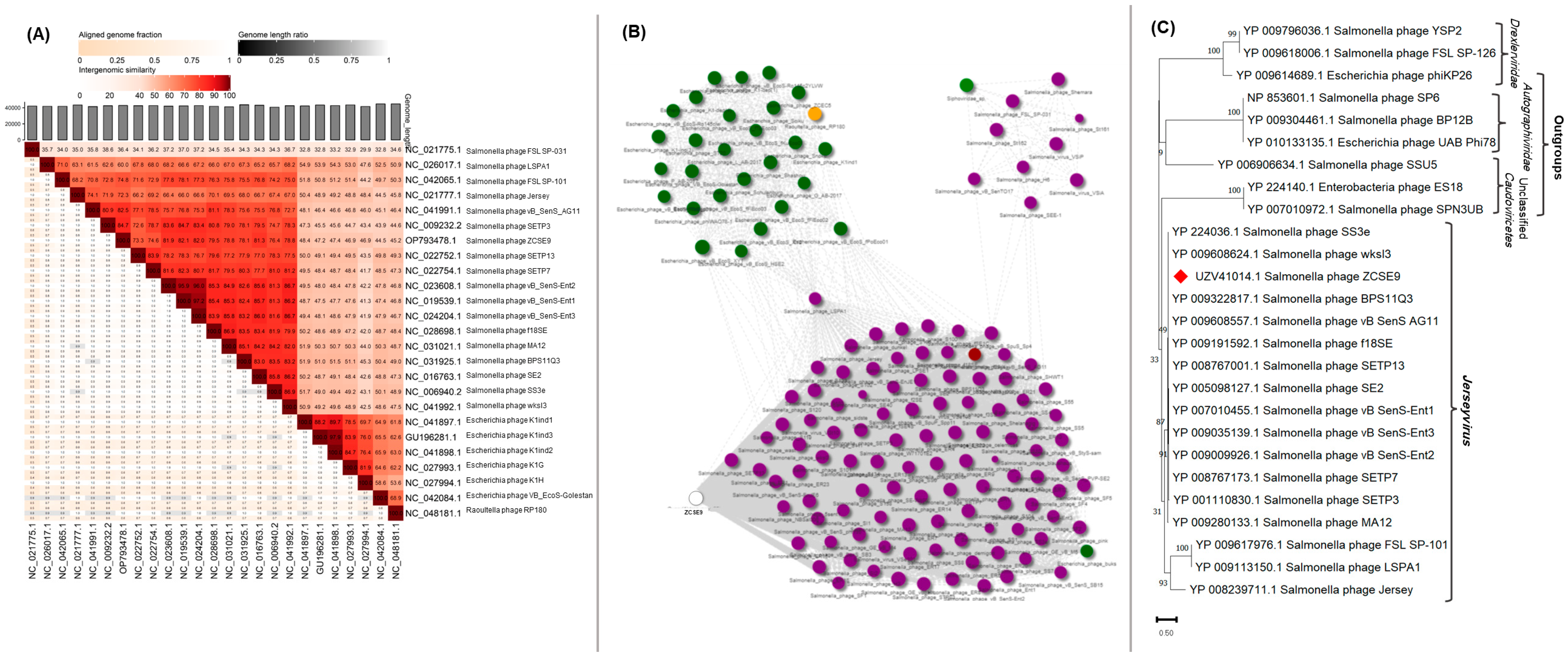
3.6. Genomic Characterization of Phage ZCSE9
3.6.1. Analysis and Annotation of the Genome
3.6.2. Phylogenetic Analysis
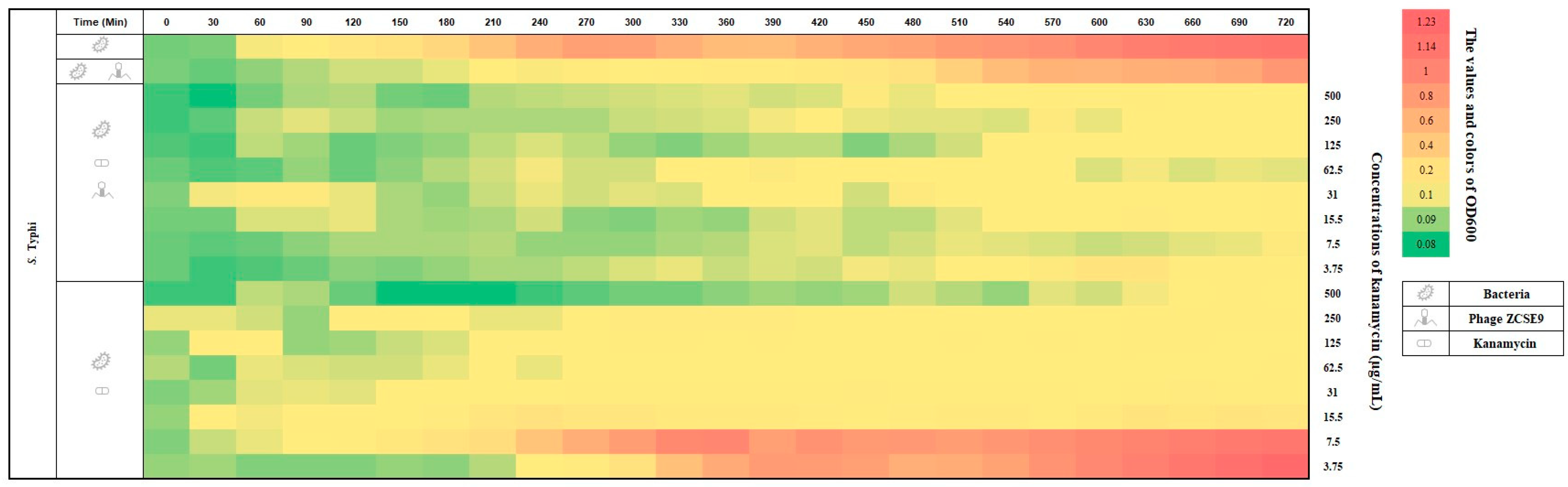
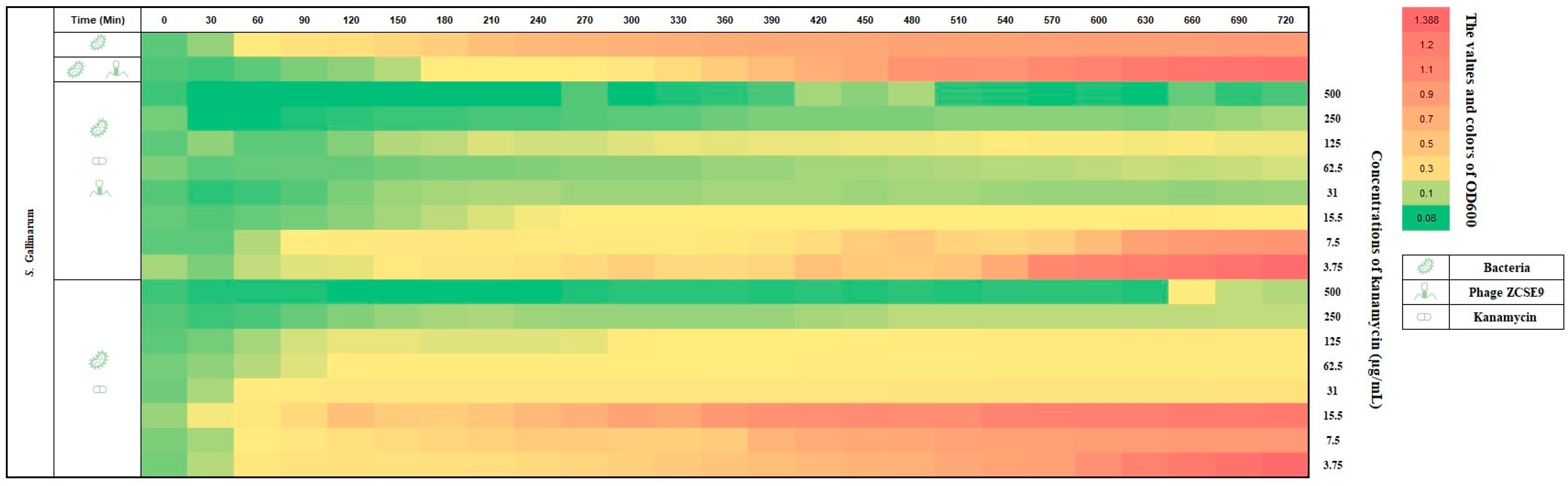
3.7. Synergistic Activity of Phage ZCSE9 and Kanamycin
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kurtz, J.R.; Goggins, J.A.; McLachlan, J.B. Salmonella Infection: Interplay between the Bacteria and Host Immune System. Immunol. Lett. 2017, 190, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Andino, A.; Hanning, I. Salmonella enterica: Survival, Colonization, and Virulence Differences among Serovars. Sci. World J. 2015, 2015, 520179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jajere, S.M. A Review of Salmonella enterica with Particular Focus on the Pathogenicity and Virulence Factors, Host Specificity and Antimicrobial Resistance Including Multidrug Resistance. Vet. World 2019, 12, 504–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Ge, C.; Li, S.; Tang, S.; Wu, X.; Luo, H.; Deng, X.; Zhang, G.; Stevenson, A.; Baker, R.C. Evaluation of Nanopore Sequencing Technology to Identify Salmonella enterica Choleraesuis Var. Kunzendorf and Orion Var. 15+, 34+. Int. J. Food Microbiol. 2021, 346, 109167. [Google Scholar] [CrossRef]
- John, J.; Van Aart, C.J.C.; Grassly, N.C. The Burden of Typhoid and Paratyphoid in India: Systematic Review and Meta-Analysis. PLoS Negl. Trop. Dis. 2016, 10, e0004616. [Google Scholar] [CrossRef] [PubMed]
- Eng, S.K.; Pusparajah, P.; Ab Mutalib, N.S.; Ser, H.L.; Chan, K.G.; Lee, L.H. Salmonella: A Review on Pathogenesis, Epidemiology and Antibiotic Resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Pui, C.F.; Wong, W.C.; Chai, L.C.; Tunung, R.; Jeyaletchumi, P.; Hidayah, N.; Ubong, A.; Farinazleen, M.G.; Cheah, Y.K.; Son, R. Salmonella: A Foodborne Pathogen. Int. Food Res. 2011, 18, 465–473. [Google Scholar]
- Mølbak, K. Human Health Consequences of Antimicrobial Drug-Resistant Salmonella and Other Foodborne Pathogens. Clin. Infect. Dis. 2005, 41, 1613–1620. [Google Scholar] [CrossRef]
- Roth, N.; Käsbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The Application of Antibiotics in Broiler Production and the Resulting Antibiotic Resistance in Escherichia coli: A Global Overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef]
- Grundin, J.; Blanco Penedo, I.; Fall, N.; Sternberg Lewerin, S. “The Swedish Experience”—A Summary on the Swedish Efforts towards a Low and Prudent Use of Antibiotics in Animal Production; SLU Framtidens djur, natur och hälsa: Uppsala, Sweden, 2020. [Google Scholar]
- Aarestrup, F.M.; Seyfarth, A.M.; Emborg, H.D.; Pedersen, K.; Hendriksen, R.S.; Bager, F. Effect of Abolishment of the Use of Antimicrobial Agents for Growth Promotion on Occurrence of Antimicrobial Resistance in Fecal Enterococci from Food Animals in Denmark. Antimicrob. Agents Chemother. 2001, 45, 2054–2059. [Google Scholar] [CrossRef] [Green Version]
- Casewell, M.; Friis, C.; Marco, E.; McMullin, P.; Phillips, I. The European Ban on Growth-Promoting Antibiotics and Emerging Consequences for Human and Animal Health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, M.J.; Legido-Quigley, H.; Hsu, L.Y. Antimicrobial Resistance in One Health. Adv. Sci. Technol. Secur. Appl. 2020, 209–229. [Google Scholar] [CrossRef]
- Wallinga, D.; Smit, L.A.M.; Davis, M.F.; Casey, J.A.; Nachman, K.E. A Review of the Effectiveness of Current US Policies on Antimicrobial Use in Meat and Poultry Production. Curr. Environ. Health Rep. 2022, 9, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Pechere, J.C. Patients’ Interviews and Misuse of Antibiotics. Clin. Infect. Dis. 2001, 33, S170–S173. [Google Scholar] [CrossRef] [Green Version]
- Agyare, C.; Boamah, V.E.; Zumbi, C.N.; Osei, F.B. Antibiotic Use in Poultry Production and Its Effects on Bacterial Resistance. In Antimicrobial Resistance—A Global Threat; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Abdelsattar, A.S.; Makky, S.; Nofal, R.; Hebishy, M.; Agwa, M.M.; Aly, R.G.; Abo El-Naga, M.Y.; Heikal, Y.A.; Fayez, M.S.; Rezk, N.; et al. Enhancement of Wound Healing via Topical Application of Natural Products: In Vitro and in Vivo Evaluations. Arab. J. Chem. 2022, 15, 103869. [Google Scholar] [CrossRef]
- El-Shibiny, A.; El-Sahhar, S. Bacteriophages: The Possible Solution to Treat Infections Caused by Pathogenic Bacteria. Can. J. Microbiol. 2017, 63, 865–879. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahman, F.; Easwaran, M.; Daramola, O.I.; Ragab, S.; Lynch, S.; Oduselu, T.J.; Khan, F.M.; Ayobami, A.; Adnan, F.; Torrents, E.; et al. Phage-Encoded Endolysins. Antibiotics 2021, 10, 124. [Google Scholar] [CrossRef] [PubMed]
- Abdelsattar, A.S.; Gouda, S.M.; Hassan, Y.Y.; Farouk, W.M.; Makky, S.; Nas, A.; Hakim, T.A.; El-Shibiny, A. In Vitro Bacteriophage-Mediated Synthesis of Silver Nanoparticles for Antibacterial Applications and Heavy Metal Detection. Mater. Lett. 2022, 318, 132184. [Google Scholar] [CrossRef]
- Theuretzbacher, U.; Bush, K.; Harbarth, S.; Paul, M.; Rex, J.H.; Tacconelli, E.; Thwaites, G.E. Critical Analysis of Antibacterial Agents in Clinical Development. Nat. Rev. Microbiol. 2020, 18, 286–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, A.; Lisboa, T.; Blot, S.; Martin-Loeches, I.; Sole-Violan, J.; De Mendoza, D.; Rello, J. Mortality in ICU Patients with Bacterial Community-Acquired Pneumonia: When Antibiotics Are Not Enough. Intensive Care Med. 2009, 35, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Keen, E.C. Phage Therapy: Concept to Cure. Front. Microbiol. 2012, 3, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasman, L.M.; Porter, L.D. Bacteriophages. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2022; pp. 280–283. [Google Scholar] [CrossRef]
- Mahony, J.; McAuliffe, O.; Ross, R.P.; van Sinderen, D. Bacteriophages as Biocontrol Agents of Food Pathogens. Curr. Opin. Biotechnol. 2011, 22, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Atterbury, R.J.; Gigante, A.M.; Rubio Lozano, M.D.L.S.; Méndez Medina, R.D.; Robinson, G.; Alloush, H.; Barrow, P.A.; Allen, V.M. Reduction of Salmonella Contamination on the Surface of Chicken Skin Using Bacteriophage. Virol. J. 2020, 17, 98. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Virk, S.M.; Shi, J.; Zhou, Y.; Willias, S.P.; Morsy, M.K.; Abdelnabby, H.E.; Liu, J.; Wang, X.; Li, J. Isolation, Characterization, and Application of Bacteriophage LPSE1 against Salmonella enterica in Ready to Eat (RTE) Foods. Front. Microbiol. 2018, 9, 1046. [Google Scholar] [CrossRef] [PubMed]
- Hungaro, H.M.; Mendonça, R.C.S.; Gouvêa, D.M.; Vanetti, M.C.D.; de Oliveira Pinto, C.L. Use of Bacteriophages to Reduce Salmonella in Chicken Skin in Comparison with Chemical Agents. Food Res. Int. 2013, 52, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Patel, J.R.; Conway, W.S.; Ferguson, S.; Sulakvelidze, A. Effectiveness of Bacteriophages in Reducing Escherichia coli O157:H7 on Fresh-Cut Cantaloupes and Lettuce. J. Food Prot. 2009, 72, 1481–1485. [Google Scholar] [CrossRef] [Green Version]
- Örmälä, A.-M.; Jalasvuori, M. Phage Therapy. Bacteriophage 2013, 3, e24219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oechslin, F. Resistance Development to Bacteriophages Occurring during Bacteriophage Therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef] [Green Version]
- Hesse, S.; Rajaure, M.; Wall, E.; Johnson, J.; Bliskovsky, V.; Gottesman, S.; Adhya, S. Phage Resistance in Multidrug-Resistant Klebsiella Pneumoniae St258 Evolves via Diverse Mutations That Culminate in Impaired Adsorption. MBio 2020, 11, e02530-19. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Loh, B.; Gordillo Altamirano, F.; Yu, Y.; Hua, X.; Leptihn, S. Colistin-Phage Combinations Decrease Antibiotic Resistance in Acinetobacter baumannii via Changes in Envelope Architecture. Emerg. Microbes Infect. 2021, 10, 2205–2219. [Google Scholar] [CrossRef]
- Chang, R.Y.K.; Das, T.; Manos, J.; Kutter, E.; Morales, S.; Chan, H.K. Bacteriophage PEV20 and Ciprofloxacin Combination Treatment Enhances Removal of Pseudomonas Aeruginosa Biofilm Isolated from Cystic Fibrosis and Wound Patients. AAPS J. 2019, 21, 49. [Google Scholar] [CrossRef]
- Letrado, P.; Corsini, B.; DÍez-Martínez, R.; Bustamante, N.; Yuste, J.E.; García, P. Bactericidal Synergism between Antibiotics and Phage Endolysin Cpl-711 to Kill Multidrug-Resistant Pneumococcus. Future Microbiol. 2018, 13, 1215–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Chang, R.Y.K.; Britton, W.J.; Morales, S.; Kutter, E.; Chan, H.K. Synergy of Nebulized Phage PEV20 and Ciprofloxacin Combination against Pseudomonas aeruginosa. Int. J. Pharm. 2018, 551, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Abdelsattar, A.S.; Safwat, A.; Nofal, R.; Elsayed, A.; Makky, S.; El-Shibiny, A. Isolation and Characterization of Bacteriophage ZCSE6 against Salmonella Spp.: Phage Application in Milk. Biologics 2021, 1, 164–176. [Google Scholar] [CrossRef]
- Taha, O.A.; Connerton, P.L.; Connerton, I.F.; El-Shibiny, A. Bacteriophage ZCKP1: A Potential Treatment for Klebsiella Pneumoniae Isolated from Diabetic Foot Patients. Front. Microbiol. 2018, 9, 2127. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Wang, Y.; Hong, B.; Li, Y.; Ma, Y.; Wang, J. Isolation and Characterization of a Lytic Vibrio Parahaemolyticus Phage VB_VpaP_GHSM17 from Sewage Samples. Viruses 2022, 14, 1601. [Google Scholar] [CrossRef]
- Bagińska, N.; Harhala, M.A.; Cieślik, M.; Orwat, F.; Weber-Dąbrowska, B.; Dąbrowska, K.; Górski, A.; Jończyk-Matysiak, E. Biological Properties of 12 Newly Isolated Acinetobacter Baumannii-Specific Bacteriophages. Viruses 2023, 15, 231. [Google Scholar] [CrossRef]
- Pickard, D.J.J. Preparation of Bacteriophage Lysates and Pure DNA. In Bacteriophages; Springer: Berlin/Heidelberg, Germany, 2009; pp. 3–9. [Google Scholar]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes de Novo Assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef]
- Besemer, J. GeneMarkS: A Self-Training Method for Prediction of Gene Starts in Microbial Genomes. Implications for Finding Sequence Motifs in Regulatory Regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A Better Web Interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARAGORN, a Program to Detect TRNA Genes and TmRNA Genes in Nucleotide Sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Hockenberry, A.J.; Wilke, C.O. BACPHLIP: Predicting Bacteriophage Lifestyle from Conserved Protein Domains. PeerJ 2021, 9, e11396. [Google Scholar] [CrossRef] [PubMed]
- Yukgehnaish, K.; Rajandas, H.; Parimannan, S.; Manickam, R.; Marimuthu, K.; Petersen, B.; Clokie, M.R.J.; Millard, A.; Sicheritz-Pontén, T. PhageLeads: Rapid Assessment of Phage Therapeutic Suitability Using an Ensemble Machine Learning Approach. Viruses 2022, 14, 342. [Google Scholar] [CrossRef]
- Zaki, B.M.; Fahmy, N.A.; Aziz, R.K.; Samir, R.; El-Shibiny, A. Characterization and Comprehensive Genome Analysis of Novel Bacteriophage, VB_Kpn_ZCKp20p, with Lytic and Anti-Biofilm Potential against Clinical Multidrug-Resistant Klebsiella Pneumoniae. Front. Cell. Infect. Microbiol. 2023, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Stothard, P.; Wishart, D.S. Circular Genome Visualization and Exploration Using CGView. Bioinformatics 2005, 21, 537–539. [Google Scholar] [CrossRef] [Green Version]
- Duarte, J.A.G. PhageDPO: Phage Depolymerase Finder. Doctoral Dissertation, Universidade do Minho, Braga, Portugal, 2021. [Google Scholar]
- Hallgren, J.; Tsirigos, K.D.; Pedersen, M.D.; Almagro Armenteros, J.J.; Marcatili, P.; Nielsen, H.; Krogh, A.; Winther, O. DeepTMHMM Predicts Alpha and Beta Transmembrane Proteins Using Deep Neural Networks. BioRxiv 2022, 2004–2022. [Google Scholar]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. ViPTree: The Viral Proteomic Tree Server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef]
- Moraru, C.; Varsani, A.; Kropinski, A.M. VIRIDIC-A Novel Tool to Calculate the Intergenomic Similarities of Prokaryote-Infecting Viruses. Viruses 2020, 12, 1268. [Google Scholar] [CrossRef]
- Rangel-Pineros, G.; Millard, A.; Michniewski, S.; Scanlan, D.; Sirén, K.; Reyes, A.; Petersen, B.; Clokie, M.R.J.; Sicheritz-Pontén, T. From Trees to Clouds: PhageClouds for Fast Comparison of 640,000 Phage Genomic Sequences and Host-Centric Visualization Using Genomic Network Graphs. Phage 2021, 2, 194–203. [Google Scholar] [CrossRef]
- Davis, P.; Seto, D.; Mahadevan, P. CoreGenes5.0: An Updated User-Friendly Webserver for the Determination of Core Genes from Sets of Viral and Bacterial Genomes. Viruses 2022, 14, 2534. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M. Phylogeny. Fr: Robust Phylogenetic Analysis for the Non-Specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Parvekar, P.; Palaskar, J.; Metgud, S.; Maria, R.; Dutta, S. The Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) of Silver Nanoparticles against Staphylococcus aureus. Biomater. Investig. Dent. 2020, 7, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.; Hu, Y.; Mizan, M.; Rahaman, F.; Yan, T.; Nime, I.; Zhou, Y.; Li, J. Characterization of Salmonella Phage LPST153 That Effectively Targets Most Prevalent Salmonella serovars. Microorganisms 2020, 8, 1089. [Google Scholar] [CrossRef] [PubMed]
- Alic, S.; Naglic, T.; Tusek-Znidari, M.; Ravnikar, M.; Racki, N.; Peterka, M.; Dreo, T. Newly Isolated Bacteriophages from the Podoviridae, Siphoviridae, and Myoviridae Families Have Variable Effects on Putative Novel Dickeya Spp. Front. Microbiol. 2017, 8, 1870. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Tu, J.; Liu, J.; Molineux, I.J. Structural Dynamics of Bacteriophage P22 Infection Initiation Revealed by Cryo-Electron Tomography. Nat. Microbiol. 2019, 4, 1049–1056. [Google Scholar] [CrossRef]
- Lee, C.-N.; Tseng, T.-T.; Lin, J.-W.; Fu, Y.-C.; Weng, S.-F.; Tseng, Y.-H. Lytic Myophage Abp53 Encodes Several Proteins Similar to Those Encoded by Host Acinetobacter Baumannii and Phage PhiKO2. Appl. Environ. Microbiol. 2011, 77, 6755–6762. [Google Scholar] [CrossRef] [Green Version]
- Dyson, Z.A.; Tucci, J.; Seviour, R.J.; Petrovski, S. Lysis to Kill: Evaluation of the Lytic Abilities, and Genomics of Nine Bacteriophages Infective for Gordonia Spp. and Their Potential Use in Activated Sludge Foam Biocontrol. PLoS ONE 2015, 10, e0134512. [Google Scholar] [CrossRef] [Green Version]
- Seed, K.D. Battling Phages: How Bacteria Defend against Viral Attack. PLoS Pathog. 2015, 11, e1004847. [Google Scholar] [CrossRef] [Green Version]
- Duc, H.M.; Son, H.M.; Yi, H.P.S.; Sato, J.; Ngan, P.H.; Masuda, Y.; Honjoh, K.-I.; Miyamoto, T. Isolation, Characterization and Application of a Polyvalent Phage Capable of Controlling Salmonella and Escherichia coli O157:H7 in Different Food Matrices. Food Res. Int. 2020, 131, 108977. [Google Scholar] [CrossRef]
- Hazem, A. Effects of Temperatures, PH-Values, Ultra-Violet Light, Ethanol and Chloroform on the Growth of Isolated Thermophilic Bacillus Phages. New Microbiol. 2002, 25, 469–476. [Google Scholar]
- McCormick, B.A.; Colgan, S.P.; Delp-Archer, C.; Miller, S.I.; Madara, J.L. Salmonella Typhimurium Attachment to Human Intestinal Epithelial Monolayers: Transcellular Signalling to Subepithelial Neutrophils. J. Cell Biol. 1993, 123, 895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, E.A.; Naliboff, B.D.; Chang, L. Evolving Pathophysiological Model of Functional Gastrointestinal Disorders: Implications for Treatment. Eur. J. Surg. Suppl. 2002, 168, 3–9. [Google Scholar]
- Abdelsattar, A.S.; Abdelrahman, F.; Dawoud, A.; Connerton, I.F.; El-Shibiny, A. Encapsulation of E. coli Phage ZCEC5 in Chitosan–Alginate Beads as a Delivery System in Phage Therapy. AMB Express 2019, 9, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwak, K.M.; Choi, I.Y.; Lee, J.; Oh, J.-H.; Park, M.-K. Isolation and Characterization of a Lytic and Highly Specific Phage against Yersinia Enterocolitica as a Novel Biocontrol Agent. J. Microbiol. Biotechnol. 2018, 28, 1946–1954. [Google Scholar] [CrossRef] [Green Version]
- Atamer, Z.; Dietrich, J.; Müller-Merbach, M.; Neve, H.; Heller, K.J.; Hinrichs, J. Screening for and Characterization of Lactococcus lactis Bacteriophages with High Thermal Resistance. Int. Dairy J. 2009, 19, 228–235. [Google Scholar] [CrossRef]
- Wang, H.; Shen, Y.; Li, P.; Xiao, Y.; Li, Y.; Hu, X.; Wang, Z.; Cheng, Z.; Wang, Z.; Liu, Q. Characterization and Genomic Analysis of a Demerecviridae Phage SP76 with Lytic Multiple-Serotypes of Salmonella. Arch. Microbiol. 2022, 204, 175. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Ding, Y.; Huang, C.; Zhang, Y.; Wang, J.; Wang, X. Characterization of a Novel Siphoviridae Salmonella Bacteriophage T156 and Its Microencapsulation Application in Food Matrix. Food Res. Int. 2021, 140, 110004. [Google Scholar] [CrossRef]
- Lin, J.T.; Kirst, S.; Cuci, S.; Klem, A.; She, Y.-M.; Kropinski, A.M.; Anany, H. Isolation, Characterization, and Genome Analysis of a Novel Bacteriophage, Escherichia Phage VB_EcoM-4HA13, Representing a New Phage Genus in the Novel Phage Family Chaseviridae. Viruses 2022, 14, 2356. [Google Scholar] [CrossRef]
- Balcao, V.M.; Moreli, F.C.; Silva, E.C.; Belline, B.G.; Martins, L.F.; Rossi, F.P.N.; Pereira, C.; Vila, M.M.D.C.; da Silva, A.M. Isolation and Molecular Characterization of a Novel Lytic Bacteriophage That Inactivates MDR Klebsiella pneumoniae Strains. Pharmaceutics 2022, 14, 1421. [Google Scholar] [CrossRef]
- Szermer-Olearnik, B.; Sochocka, M.; Zwolinska, K.; Ciekot, J.; Czarny, A.; Szydzik, J.; Kowalski, K.; Boratynski, J. Comparison of Microbiological and Physicochemical Methods for Enumeration of Microorganisms. Postęp. Hig. Med. Dośw. 2014, 68, 1392–1396. [Google Scholar] [CrossRef]
- Costa, P.; Gomes, A.T.P.C.; Braz, M.; Pereira, C.; Almeida, A. Application of the Resazurin Cell Viability Assay to Monitor Escherichia coli and Salmonella typhimurium Inactivation Mediated by Phages. Antibiotics 2021, 10, 974. [Google Scholar] [CrossRef] [PubMed]
- Hoyle, N.; Kutter, E. Phage Therapy: Bacteriophages as Natural, Self-Replicating Antimicrobials. In Practical Handbook of Microbiology; CRC Press: Boca Raton, FL, USA, 2021; pp. 801–824. ISBN 1003099270. [Google Scholar]
- Wang, C.; Chen, Q.; Zhang, C.; Yang, J.; Lu, Z.; Lu, F.; Bie, X. Characterization of a Broad Host-Spectrum Virulent Salmonella Bacteriophage Fmb-P1 and Its Application on Duck Meat. Virus Res. 2017, 236, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Van Belleghem, J.D.; de Vries, C.R.; Burgener, E.; Chen, Q.; Manasherob, R.; Aronson, J.R.; Amanatullah, D.F.; Tamma, P.D.; Suh, G.A. The Safety and Toxicity of Phage Therapy: A Review of Animal and Clinical Studies. Viruses 2021, 13, 1268. [Google Scholar] [CrossRef]
- Garneau, J.R.; Depardieu, F.; Fortier, L.-C.; Bikard, D.; Monot, M. PhageTerm: A Tool for Fast and Accurate Determination of Phage Termini and Packaging Mechanism Using next-Generation Sequencing Data. Sci. Rep. 2017, 7, 8292. [Google Scholar] [CrossRef] [PubMed]
- Baxa, U.; Weintraub, A.; Seckler, R. Self-Competitive Inhibition of the Bacteriophage P22 Tailspike Endorhamnosidase by O-Antigen Oligosaccharides. Biochemistry 2020, 59, 4845–4855. [Google Scholar] [CrossRef]
- Subramanian, S.; Dover, J.A.; Parent, K.N.; Doore, S.M. Host Range Expansion of Shigella Phage Sf6 Evolves through Point Mutations in the Tailspike. J. Virol. 2022, 96, e00929-22. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Yan, Y.; Ji, W.; Du, B.; Meng, X.; Wang, H.; Sun, J. Characterization and Determination of Holin Protein of Streptococcus suis Bacteriophage SMP in Heterologous Host. Virol. J. 2012, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Comeau, A.M.; Tétart, F.; Trojet, S.N.; Prère, M.F.; Krisch, H.M. La «Synergie Phages-Antibiotiques»—Un Enjeu Pour La Phagothérapie. Médecine/Science 2008, 24, 449–451. [Google Scholar] [CrossRef]
- Cooper, C.J.; Mirzaei, M.K.; Nilsson, A.S. Adapting Drug Approval Pathways for Bacteriophage-Based Therapeutics. Front. Microbiol. 2016, 7, 1209. [Google Scholar] [CrossRef] [Green Version]
- Llor, C.; Bjerrum, L. Antimicrobial Resistance: Risk Associated with Antibiotic Overuse and Initiatives to Reduce the Problem. Ther. Adv. Drug Saf. 2014, 5, 229. [Google Scholar] [CrossRef] [Green Version]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage Selection Restores Antibiotic Sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engeman, E.; Freyberger, H.R.; Corey, B.W.; Ward, A.M.; He, Y.; Nikolich, M.P.; Filippov, A.A.; Tyner, S.D.; Jacobs, A.C. Synergistic Killing and Re-Sensitization of Pseudomonas Aeruginosa to Antibiotics by Phage-Antibiotic Combination Treatment. Pharmaceuticals 2021, 14, 184. [Google Scholar] [CrossRef] [PubMed]
- Petsong, K.; Uddin, M.J.; Vongkamjan, K.; Ahn, J. Combined Effect of Bacteriophage and Antibiotic on the Inhibition of the Development of Antibiotic Resistance in Salmonella typhimurium. Food Sci. Biotechnol. 2018, 27, 1239–1244. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Salmonella Bacteria | The Bacteriolytic Activity of ZCSE9 |
|---|---|---|
| CMPZC52 int | Salmonella enterica Blegdam II | + |
| CMPZC49G.B | Salmonella enterica Gallinarum | + |
| CMPZC78 Sple | Salmonella enterica Enteritidis | + |
| CMPZC18 | Salmonella enterica spp. | + |
| CMPZC77 liv | Salmonella enterica Kentucky | + |
| CMPZC53 sple | Salmonella enterica Blegdam I | + |
| CMPZC85 liv | Salmonella enterica Virchom | − |
| CMPZC29 int | Salmonella enterica Enteritidis II | + |
| CMPZC49 liv | Salmonella enterica Blegdam III | + |
| CMPZC52 liv (15) | Salmonella enterica spp. | + |
| CMPZC2 | Salmonella enterica Kentucky II | − |
| CMPZC3 | Salmonella enterica spp. | − |
| CMPZC7 L | Salmonella enterica Typhimurium II | − |
| CMPZC77 int | Salmonella enterica Kentucky III | − |
| CMPZC100 Sple | Salmonella enterica Nigeria | − |
| CMPZC84 G.B | Salmonella enterica spp. | − |
| CMPZC52 Sple (7) | Salmonella enterica Blegdam IV | − |
| CMPZC 77 liv | Salmonella enterica Kentucky IV | + |
| CMPZC20 | Salmonella enterica Typhi II | − |
| CMPZC33 | Salmonella enterica Typhimurium III | − |
| CMPZC34 | Salmonella enterica spp. | + |
| CMPZC37 | Salmonella enterica spp. | − |
| NCTC 13348 | Salmonella enterica Typhimurium | + |
| NCTC 160 | Salmonella enterica Typhi | + |
| Bacteria | ID | Kanamycin Concentration (µg/mL) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 500 | 250 | 125 | 62 | 30 | 15 | 7.5 | 3.75 | |||
| Kanamycin | S. Typhi | NCTC 160 | √ | |||||||
| S. Typhimurium | NCTC 13348 | √ | ||||||||
| S. Gallinarum | CMPZC49G.B | √ | ||||||||
| S. Blegdam I | CMPZC53 sple | √ | ||||||||
| S. Blegdam II | CMPZC52 int | √ | ||||||||
| S. Enteritidis | CMPZC78 Sple | √ | ||||||||
| Phage MOI 0.1 + Kanamycin | S. Typhi | NCTC 160 | √ | |||||||
| S. Typhimurium | NCTC 13348 | √ | ||||||||
| S. Gallinarum | CMPZC49G.B | √ | ||||||||
| S. Blegdam I | CMPZC53 sple | √ | ||||||||
| S. Blegdam II | CMPZC52 int | √ | ||||||||
| S. Typhi | CMPZC78 Sple | √ | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelsattar, A.S.; Eita, M.A.; Hammouda, Z.K.; Gouda, S.M.; Hakim, T.A.; Yakoup, A.Y.; Safwat, A.; El-Shibiny, A. The Lytic Activity of Bacteriophage ZCSE9 against Salmonella enterica and Its Synergistic Effects with Kanamycin. Viruses 2023, 15, 912. https://doi.org/10.3390/v15040912
Abdelsattar AS, Eita MA, Hammouda ZK, Gouda SM, Hakim TA, Yakoup AY, Safwat A, El-Shibiny A. The Lytic Activity of Bacteriophage ZCSE9 against Salmonella enterica and Its Synergistic Effects with Kanamycin. Viruses. 2023; 15(4):912. https://doi.org/10.3390/v15040912
Chicago/Turabian StyleAbdelsattar, Abdallah S., Mohamed Atef Eita, Zainab K. Hammouda, Shrouk Mohamed Gouda, Toka A. Hakim, Aghapy Yermans Yakoup, Anan Safwat, and Ayman El-Shibiny. 2023. "The Lytic Activity of Bacteriophage ZCSE9 against Salmonella enterica and Its Synergistic Effects with Kanamycin" Viruses 15, no. 4: 912. https://doi.org/10.3390/v15040912
APA StyleAbdelsattar, A. S., Eita, M. A., Hammouda, Z. K., Gouda, S. M., Hakim, T. A., Yakoup, A. Y., Safwat, A., & El-Shibiny, A. (2023). The Lytic Activity of Bacteriophage ZCSE9 against Salmonella enterica and Its Synergistic Effects with Kanamycin. Viruses, 15(4), 912. https://doi.org/10.3390/v15040912