

Noncoding RNA of Zika Virus Affects Interplay between Wnt-Signaling and Pro-Apoptotic Pathways in the Developing Brain Tissue

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
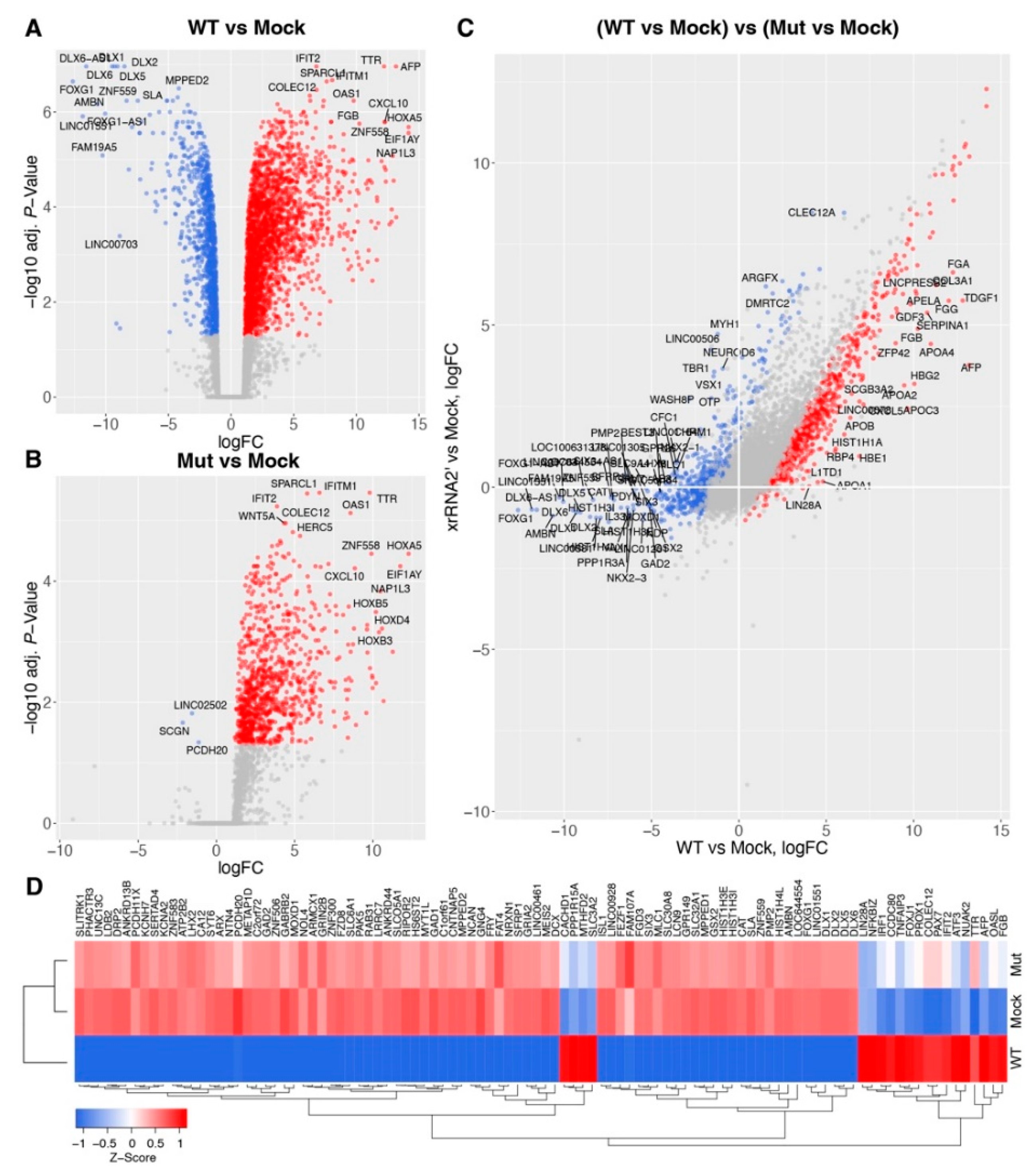
3.1. Production of sfRNA Alters Transcriptome of ZIKV-Infected Human Brain Organoids
3.2. Production of sfRNA in ZIKV-Infected Human Brain Organoids Affects Activity of Multiple Pathways Related to Brain Development
3.3. ZIKV sfRNA Affects Brain Development via a Link between Pro-Apoptotic Signaling and Wnt Pathway
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pierson, T.C.; Diamond, M.S. The Emergence of Zika Virus and Its New Clinical Syndromes. Nature 2018, 560, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Baronti, C.; Lieutaud, P.; Bardsley, M.; de Lamballerie, X.; Resman Rus, K.; Korva, M.; Petrovec, M.; Avsic-Zupanc, T.; Matusali, G.; Meschi, S.; et al. The Importance of Biobanking for Response to Pandemics Caused by Emerging Viruses: The European Virus Archive As an Observatory of the Global Response to the Zika Virus and COVID-19 Crisis. Biopreservation Biobanking 2020, 18, 561–569. [Google Scholar] [CrossRef]
- Valentine, M.J.; Murdock, C.C.; Kelly, P.J. Sylvatic Cycles of Arboviruses in Non-Human Primates. Parasites Vectors 2019, 12, 463. [Google Scholar] [CrossRef] [PubMed]
- Falcao, M.B.; Cimerman, S.; Luz, K.G.; Chebabo, A.; Brigido, H.A.; Lobo, I.M.; Timerman, A.; Angerami, R.N.; Cunha, C.A.; Bacha, H.A.; et al. Management of Infection by the Zika Virus. Ann. Clin. Microbiol. Antimicrob. 2016, 15, 57. [Google Scholar] [CrossRef]
- Mlakar, J.; Korva, M.; Tul, N.; Popović, M.; Poljšak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Rus, K.R.; Vipotnik, T.V.; Vodušek, V.F.; et al. Zika Virus Associated with Microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Calvet, G.; Aguiar, R.S.; Melo, A.S.O.; Sampaio, S.A.; de Filippis, I.; Fabri, A.; Araujo, E.S.M.; de Sequeira, P.C.; de Mendonça, M.C.L.; de Oliveira, L.; et al. Detection and Sequencing of Zika Virus from Amniotic Fluid of Fetuses with Microcephaly in Brazil: A Case Study. Lancet Infect. Dis. 2016, 16, 653–660. [Google Scholar] [CrossRef]
- Rice, M.E.; Galang, R.R.; Roth, N.M.; Ellington, S.R.; Moore, C.A.; Valencia-Prado, M.; Ellis, E.M.; Tufa, A.J.; Taulung, L.A.; Alfred, J.M.; et al. Vital Signs: Zika-Associated Birth Defects and Neurodevelopmental Abnormalities Possibly Associated with Congenital Zika Virus Infection—U.S. Territories and Freely Associated States, 2018. Morb. Mortal. Wkly. Rep. 2018, 67, 858–867. [Google Scholar] [CrossRef]
- Pijlman, G.P.; Funk, A.; Kondratieva, N.; Leung, J.; Torres, S.; van der Aa, L.; Liu, W.J.; Palmenberg, A.C.; Shi, P.-Y.Y.; Hall, R.A.; et al. A Highly Structured, Nuclease-Resistant, Noncoding RNA Produced by Flaviviruses Is Required for Pathogenicity. Cell Host Microbe 2008, 4, 579–591. [Google Scholar] [CrossRef]
- Funk, A.; Truong, K.; Nagasaki, T.; Torres, S.; Floden, N.; Balmori Melian, E.; Edmonds, J.; Dong, H.; Shi, P.-Y.; Khromykh, A. a RNA Structures Required for Production of Subgenomic Flavivirus RNA. J. Virol. 2010, 84, 11407–11417. [Google Scholar] [CrossRef]
- Villordo, S.M.; Carballeda, J.M.; Filomatori, C.V.; Gamarnik, A.V. RNA Structure Duplications and Flavivirus Host Adaptation. Trends Microbiol. 2016, 24, 270–283. [Google Scholar] [CrossRef]
- Slonchak, A.; Parry, R.; Pullinger, B.; Sng, J.D.J.; Wang, X.; Buck, T.F.; Torres, F.J.; Harrison, J.J.; Colmant, A.M.G.; Hobson-Peters, J.; et al. Structural Analysis of 3’UTRs in Insect Flaviviruses Reveals Novel Determinants of SfRNA Biogenesis and Provides New Insights into Flavivirus Evolution. Nat. Commun. 2022, 13, 1279. [Google Scholar] [CrossRef]
- Slonchak, A.; Khromykh, A.A. Subgenomic Flaviviral RNAs: What Do We Know after the First Decade of Research. Antivir. Res. 2018, 159, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Setoh, Y.X.; Amarilla, A.A.; Peng, N.Y.; Slonchak, A.; Periasamy, P.; Figueiredo, L.T.M.; Aquino, V.H.; Khromykh, A.A. Full Genome Sequence of Rocio Virus Reveal Substantial Variations from the Prototype Rocio Virus SPH 34675 Sequence. Arch. Virol. 2018, 163, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Clarke, B.D.; Roby, J.A.; Slonchak, A.; Khromykh, A.A. Functional Non-Coding RNAs Derived from the Flavivirus 3’ Untranslated Region. Virus Res. 2015, 206, 53–61. [Google Scholar] [CrossRef]
- Akiyama, B.M.; Laurence, H.M.; Massey, A.R.; Costantino, D.A.; Xie, X.; Yang, Y.; Shi, P.-Y.Y.; Nix, J.C.; Beckham, J.D.; Kieft, J.S. Zika Virus Produces Noncoding RNAs Using a Multi-Pseudoknot Structure That Confounds a Cellular Exonuclease. Science 2016, 354, 1148–1152. [Google Scholar] [CrossRef] [PubMed]
- Slonchak, A.; Hugo, L.E.; Freney, M.E.; Hall-Mendelin, S.; Amarilla, A.A.; Torres, F.J.; Setoh, Y.X.; Peng, N.Y.G.; Sng, J.D.J.; Hall, R.A.; et al. Zika Virus Noncoding RNA Suppresses Apoptosis and Is Required for Virus Transmission by Mosquitoes. Nat. Commun. 2020, 11, 2205. [Google Scholar] [CrossRef]
- Slonchak, A.; Wang, X.; Aguado, J.; Sng, J.D.J.; Chaggar, H.; Freney, M.E.; Yan, K.; Torres, F.J.; Amarilla, A.A.; Balea, R.; et al. Zika Virus Noncoding RNA Cooperates with the Viral Protein NS5 to Inhibit STAT1 Phosphorylation and Facilitate Viral Pathogenesis. Sci. Adv. 2022, 8, eadd8095. [Google Scholar] [CrossRef]
- Pallarés, H.M.; Soledad, G.; Navarro, C.; Villordo, S.M.; Merwaiss, F.; De Borba, L.; Gonzalez, M.M.; Ledesma, L.; Ojeda, D.S.; Henrion-Lacritick, A.; et al. Zika Virus Subgenomic Flavivirus RNA Generation Requires Cooperativity between Duplicated RNA Structures That Are Essential for Productive Infection in Human Cells. J. Virol. 2020, 94, 343–363. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Renner, M.; Martin, C.A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral Organoids Model Human Brain Development and Microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef]
- Qian, X.; Nguyen, H.N.; Jacob, F.; Song, H.; Ming, G. Using Brain Organoids to Understand Zika Virus-Induced Microcephaly. Development 2017, 144, 952–957. [Google Scholar] [CrossRef]
- Setoh, Y.X.; Amarilla, A.A.; Peng, N.Y.G.; Griffiths, R.E.; Carrera, J.; Freney, M.E.; Nakayama, E.; Ogawa, S.; Watterson, D.; Modhiran, N.; et al. Determinants of Zika Virus Host Tropism Uncovered by Deep Mutational Scanning. Nat. Microbiol. 2019, 4, 876–887. [Google Scholar] [CrossRef] [PubMed]
- Slonchak, A.; Clarke, B.; Mackenzie, J.; Amarilla, A.A.; Setoh, Y.X.; Khromykh, A.A. West Nile Virus Infection and Interferon Alpha Treatment Alter the Spectrum and the Levels of Coding and Noncoding Host RNAs Secreted in Extracellular Vesicles. BMC Genom. 2019, 20, 474. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Nusse, R. Wnt/β-Catenin Signaling and Disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef]
- Honein, M.A.; Dawson, A.L.; Petersen, E.E.; Jones, A.M.; Lee, E.H.; Yazdy, M.M.; Ahmad, N.; Macdonald, J.; Evert, N.; Bingham, A.; et al. Birth Defects among Fetuses and Infants of US Women with Evidence of Possible Zika Virus Infection during Pregnancy. JAMA 2017, 317, 59–68. [Google Scholar] [CrossRef]
- Sparks, H.; Monogue, B.; Akiyama, B.; Kieft, J.; Beckham, J.D. Disruption of Zika Virus XrRNA1-Dependent SfRNA1 Production Results in Tissue-Specific Attenuated Viral Replication. Viruses 2020, 12, 1177. [Google Scholar] [CrossRef]
- Tiwari, S.K.; Dang, J.; Qin, Y.; Lichinchi, G.; Bansal, V.; Rana, T.M. Zika Virus Infection Reprograms Global Transcription of Host Cells to Allow Sustained Infection. Emerge Microbes Infect. 2017, 5, 50. [Google Scholar] [CrossRef]
- Zhang, F.; Hammack, C.; Ogden, S.C.; Cheng, Y.; Lee, E.M.; Wen, Z.; Qian, X.; Nguyen, H.N.; Li, Y.; Yao, B.; et al. Molecular Signatures Associated with ZIKV Exposure in Human Cortical Neural Progenitors. Nucleic Acids Res. 2016, 44, 8610–8620. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Hua, S.; Chen, H.-R.R.; Ouyang, Z.; Einkauf, K.; Tse, S.; Ard, K.; Ciaranello, A.; Yawetz, S.; Sax, P.; et al. Transcriptional Changes during Naturally Acquired Zika Virus Infection Render Dendritic Cells Highly Conducive to Viral Replication. Cell Rep. 2017, 21, 3471–3482. [Google Scholar] [CrossRef]
- Kumamoto, T.; Hanashima, C. Evolutionary Conservation and Conversion of Foxg1 Function in Brain Development. Dev. Growth Differ. 2017, 59, 258–269. [Google Scholar] [CrossRef]
- Hou, P.-S.; Chuang, C.-Y.; Kao, C.-F.; Chou, S.-J.; Stone, L.; Ho, H.-N.; Chien, C.-L.; Kuo, H.-C. LHX2 Regulates the Neural Differentiation of Human Embryonic Stem Cells via Transcriptional Modulation of PAX6 and CER1. Nucleic Acids Res. 2013, 41, 7753–7770. [Google Scholar] [CrossRef]
- Caires-Júnior, L.C.; Goulart, E.; Melo, U.S.; Araujo, B.S.H.; Alvizi, L.; Soares-Schanoski, A.; De Oliveira, D.F.; Kobayashi, G.S.; Griesi-Oliveira, K.; Musso, C.M.; et al. Discordant Congenital Zika Syndrome Twins Show Differential In Vitro Viral Susceptibility of Neural Progenitor Cells. Nat. Commun. 2018, 9, 475. [Google Scholar] [CrossRef]
- Jiang, X.; Dong, X.; Li, S.H.; Zhou, Y.P.; Rayner, S.; Xia, H.M.; Gao, G.F.; Yuan, H.; Tang, Y.P.; Luo, M.H. Proteomic Analysis of Zika Virus Infected Primary Human Fetal Neural Progenitors Suggests a Role for Doublecortin in the Pathological Consequences of Infection in the Cortex. Front. Microbiol. 2018, 9, 1067. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Salic, A.; Krüger, R.; Heinrich, R.; Kirschner, M.W. The Roles of APC and Axin Derived from Experimental and Theoretical Analysis of the Wnt Pathway. PLoS Biol. 2003, 1, e10. [Google Scholar] [CrossRef]
- Lin, J.-J.; Chin, T.-Y.; Chen, C.-P.; Chan, H.-L.; Wu, T.-Y. Zika Virus: An Emerging Challenge for Obstetrics and Gynecology. Taiwan J. Obs. Gynecol. 2017, 56, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, Z.F.; Kulikauskas, R.M.; Bomsztyk, K.; Moon, R.T.; Chien, A.J. Activation of Wnt/β-Catenin Signaling Increases Apoptosis in Melanoma Cells Treated with Trail. PLoS ONE 2013, 8, e69593. [Google Scholar] [CrossRef]
- Pai, S.G.; Carneiro, B.A.; Mota, J.M.; Costa, R.; Leite, C.A.; Barroso-Sousa, R.; Kaplan, J.B.; Chae, Y.K.; Giles, F.J. Wnt/Beta-Catenin Pathway: Modulating Anticancer Immune Response. J. Hematol. Oncol. 2017, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Ai, P.; Xu, X.; Xu, S.; Wei, Z.; Tan, S.; Li, J. Overexpression of Wnt7a Enhances Radiosensitivity of Non-Small-Cell Lung Cancer via the Wnt/JNK Pathway. Biol. Open 2020, 9, bio050575. [Google Scholar] [CrossRef]
- Carta, L.; Sassoon, D. Wnt7a Is a Suppressor of Cell Death in the Female Reproductive Tract and Is Required for Postnatal and Estrogen-Mediated Growth1. Biol Reprod 2004, 71, 444–454. [Google Scholar] [CrossRef]
- Peng, S.; Zhang, J.; Chen, J.; Wang, H. Effects of Wnt5a Protein on Proliferation and Apoptosis in JAR Choriocarcinoma Cells. Mol. Med. Rep. 2011, 4, 99–104. [Google Scholar] [CrossRef]
- Magni, M.; Buscemi, G.; Maita, L.; Peng, L.; Chan, S.Y.; Montecucco, A.; Delia, D.; Zannini, L. TSPYL2 Is a Novel Regulator of SIRT1 and P300 Activity in Response to DNA Damage. Cell Death Differ. 2019, 26, 918–931. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, N.; Qin, C.F.; Liu, X.; Shi, L.; Xu, Z. Zika Virus Disrupts Neural Progenitor Development and Leads to Microcephaly in Mice. Cell Stem Cell 2016, 19, 120–126. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slonchak, A.; Chaggar, H.; Aguado, J.; Wolvetang, E.; Khromykh, A.A. Noncoding RNA of Zika Virus Affects Interplay between Wnt-Signaling and Pro-Apoptotic Pathways in the Developing Brain Tissue. Viruses 2023, 15, 1062. https://doi.org/10.3390/v15051062
Slonchak A, Chaggar H, Aguado J, Wolvetang E, Khromykh AA. Noncoding RNA of Zika Virus Affects Interplay between Wnt-Signaling and Pro-Apoptotic Pathways in the Developing Brain Tissue. Viruses. 2023; 15(5):1062. https://doi.org/10.3390/v15051062
Chicago/Turabian StyleSlonchak, Andrii, Harman Chaggar, Julio Aguado, Ernst Wolvetang, and Alexander A. Khromykh. 2023. "Noncoding RNA of Zika Virus Affects Interplay between Wnt-Signaling and Pro-Apoptotic Pathways in the Developing Brain Tissue" Viruses 15, no. 5: 1062. https://doi.org/10.3390/v15051062
APA StyleSlonchak, A., Chaggar, H., Aguado, J., Wolvetang, E., & Khromykh, A. A. (2023). Noncoding RNA of Zika Virus Affects Interplay between Wnt-Signaling and Pro-Apoptotic Pathways in the Developing Brain Tissue. Viruses, 15(5), 1062. https://doi.org/10.3390/v15051062