
Effect of Vaccination on Platelet Mitochondrial Bioenergy Function of Patients with Post-Acute COVID-19
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Patients
2.1.1. Control Group (C)
2.1.2. Vaccinated Patients with Post-Acute COVID-19 (V + PAC19) 2 Weeks after Infection
2.1.3. Unvaccinated Patients with Post-Acute COVID-19 (PAC19) 4–7 Weeks after Infection
2.2. Methods
2.2.1. Platelet Preparation
2.2.2. High-Resolution Respirometry Method
2.2.3. High-Performance Liquid Chromatography (HPLC)—Determination of CoQ10-TOTAL, α-tocopherol, γ-tocopherol, β-carotene
2.2.4. TBARS
2.2.5. Data Analysis
3. Results
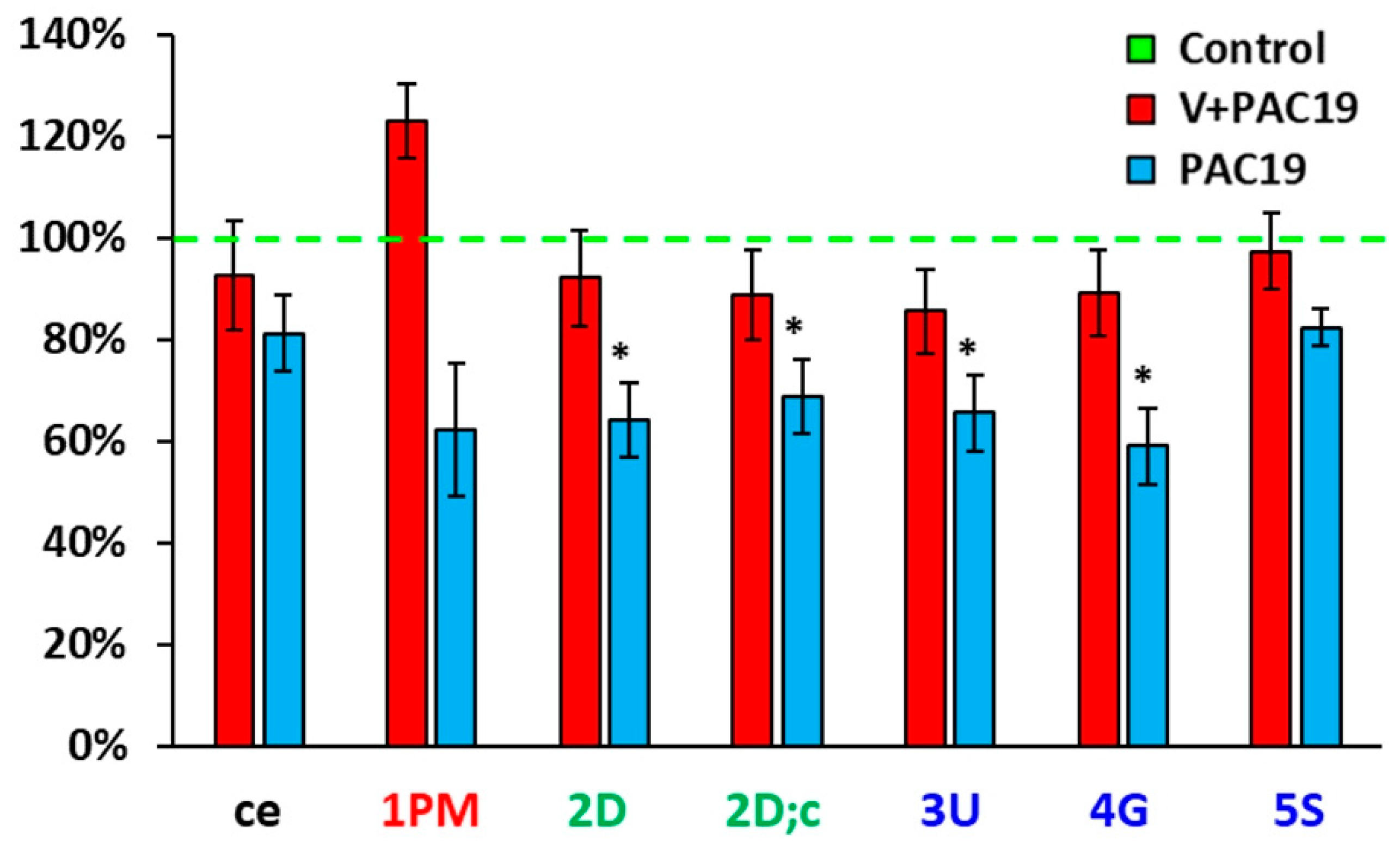
3.1. The Effect of Vaccination on the Platelet Mitochondrial Bioenergy Function in Patients with Post-Acute COVID-19 (V + PAC19)
3.2. The Effect of SARS-CoV-2 Virus on the Platelet Mitochondrial Bioenergy Function in Unvaccinated Patients with Post-Acute COVID-19 (PAC19)
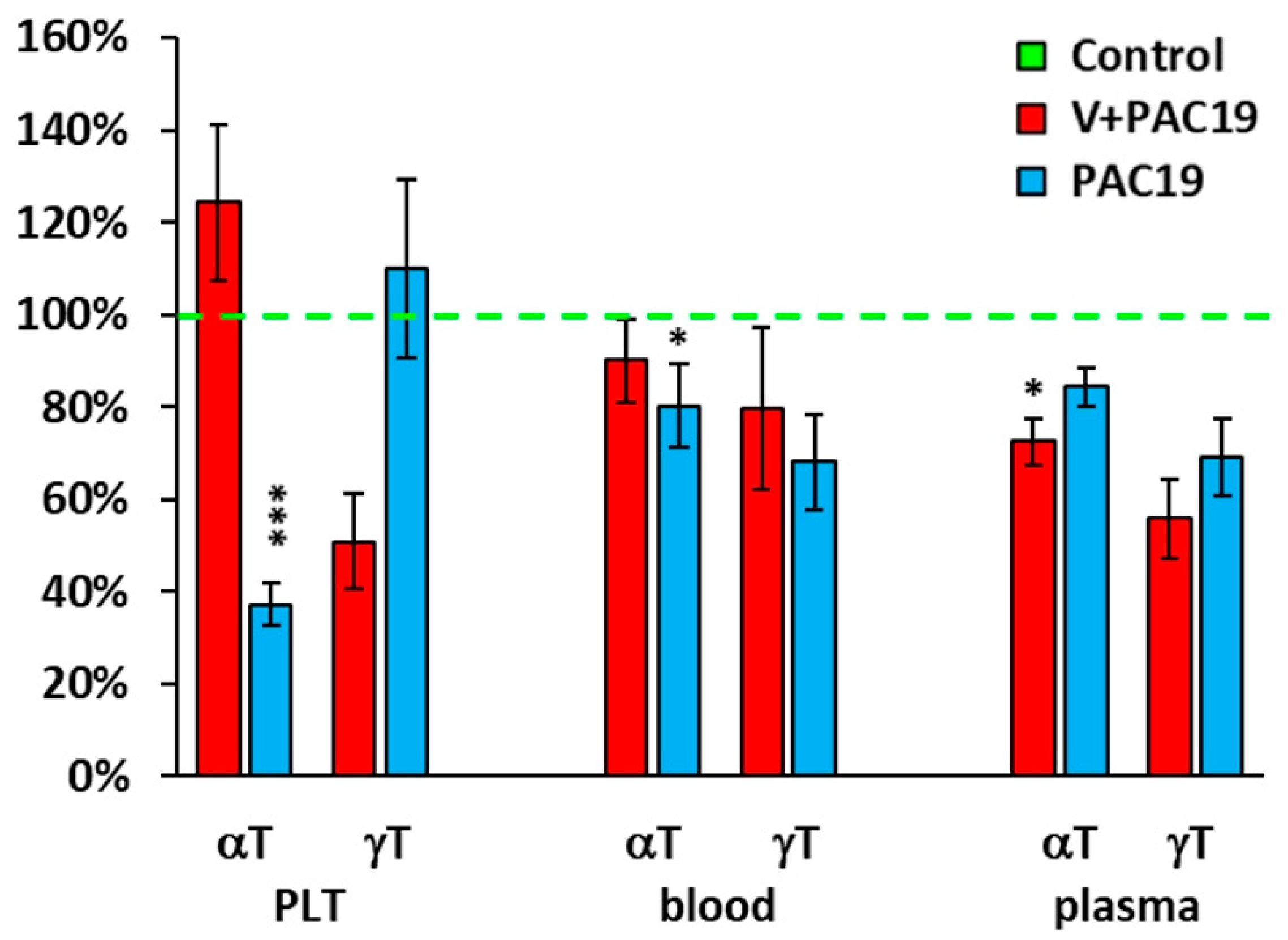
3.3. The Effect of SARS-CoV-2 Virus on the Concentration of CoQ10-TOTAL in Platelets, Blood, Plasma and TBARS in Vaccinated and Unvaccinated Patients with Post-Acute COVID-19 (V + PAC19, PAC19)
3.4. The Effect of the SARS-CoV-2 Virus on the Antioxidants in Platelets, Blood, and Plasma in Vaccinated and Unvaccinated Patients with Post-Acute COVID-19 (V + PAC19, PAC19)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cucinotta, D.; Vanelli, M. WHO declares COVID-19 a pandemic. Acta Biomed. 2020, 91, 157–160. [Google Scholar] [CrossRef]
- WHO (World Health Organization). Coronvirus Diseas 2019 (COVID-19) Situation Report-121. 2022. Available online: https://apps.who.int/iris/handle/10665/332156 (accessed on 5 December 2022).
- Nalbandian, A.; Sehgal, K.; Gupta, A.; Madhavan, M.V.; McGroder, C.; Stevens, J.S.; Cook, J.R.; Nordvig, A.S.; Shalev, D.; Sehrawat, T.S.; et al. Post-acute COVID-19 syndrome. Nat. Med. 2021, 27, 601–615. [Google Scholar] [CrossRef]
- Fugazzaro, S.; Contri, S.; Esserough, O.; Kaleci, S.; Croci, S.; Massari, M.; Facciolongo, N.C.; Besuti, G.; Iori, M.; Salvarani, C.; et al. COVID-19 Working Group Rehabilitation interventionas for post-acute COVID-19 syndrome: A systematic review. Int. J. Environ. Res. Public Health 2022, 19, 5185. [Google Scholar] [CrossRef] [PubMed]
- Taboada, M.; Carinena, A.; Moreno, E.; Rodriguez, N.; Dominguez, M.J.; Casal, A.; Riveiro, V.; Diaz-Vieito, M.; Valdes, L.; Alvarez, J.; et al. Post-COVID-19 functional status six-months after hospitalization. J. Inf. Secur. 2021, 82, e31–e33. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lluch, G. Mitochondrial activity and dynamics changes regarding metabolism in ageing and obesity. Mech. Ageing Dev. 2017, 162, 108–121. [Google Scholar] [CrossRef]
- Fernandez-Ayala, D.J.M.; Navas, P.; Lopez-Lluch, G. Age-related mitochondrial dysfunction as a key factor in COVID-19 disease. Exp. Gerontol. 2020, 142, 111147. [Google Scholar] [CrossRef]
- Nunn, A.V.W.; Guy, G.W.; Brysch, W.; Botchway, S.W.; Frasch, W.; Calabreses, E.J.; Bell, J.D. SARS-CoV-2 and mitochondrial health: Implications of lifestyle and ageing. Immun. Ageing 2020, 17, 33. [Google Scholar] [CrossRef]
- Ganji, R.; Reddy, P.H. Impact of COVID-19 on mitochondrial based immunity in ageing and age-related diseases. Front. Ageing Neurosci. 2021, 12, 614650. [Google Scholar] [CrossRef] [PubMed]
- Ohta, A.; Nishiyama, Y. Mitochondria and viruses. Mitochondrion 2011, 11, 1–12. [Google Scholar] [CrossRef]
- Gvozdjakova, A.; Klaučo, F.; Kucharska, J.; Sumbalova, Z. Is mitochondrial bioenergetics and coenzyme Q10 target of a virus causing COVID-19? Bratisl. Med. J. 2020, 121, 775–778. [Google Scholar] [CrossRef]
- Gvozdjáková, A.; Sumbalová, Z.; Kucharská, J.; Rausová, Z.; Kovalčíková, E.; Takácsová, T.; Navas, P.; López-Lluch, G.; Mojto, V.; Palacka, P. Mountain spa rehabilitation improved health of patients with post-COVID-19 syndrome: Pilot study. Environ. Sci. Pollut. Res. Int. 2023, 30, 14200–14211. [Google Scholar] [CrossRef] [PubMed]
- Sumbalová, Z.; Kucharská, J.; Rausová, Z.; Palacka, P.; Kovalcíková, E.; Takácsová, T.; Mojto, V.; Navas, P.; López-Lluch, G.; Gvozdjáková, A. Reduced platelet mitochondrial respiration and oxidative phosphorylation in patients with post COVID-19 syndrome are regenerated after spa rehabilitation and targeted ubiquinol therapy. Front. Mol. Biosci. Cell Biochem. 2022, 9, 1016352. [Google Scholar] [CrossRef] [PubMed]
- Aleksova, A.; Gagno, G.; Sinagra, G.; Beltrami, A.P.; Janjusevic, M.; Ippolito, G.; Zumla, A.; Fluca, A.L.; Ferro, F. Effects of SARS-CoV-2 on cardiovascular system: The dual role of angiotensin-converting enzyme 2 (ACE2) as the virus receptor and homeostasis regulator-review. Int. J. Mol. Sci. 2021, 22, 4526. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Zhang, J.; Fang, Y.; Lu, S.; Wu, J.; Zheng, X.; Deng, F. SARS-CoV-2 interacts with platelets and megakaryocytes via ACE2-independent mechanism. J. Hematol. Oncol. 2021, 14, 72. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yang, Y.; Li, Y.; Wang, Z.; Ma, F.; Luo, R.; Xu, X.; Zhou, G.; Wang, J.; Niu, J.; et al. Platelets mediate inflammatory monocytes activation by SARS-CoV-2 spike protein. J. Clin. Investig. 2022, 132, e150501. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.T.; Yang, F.Y.; Liu, C.; Cao, X.; Lu, J.; Zhang, X.L.; Yuan, M.X.; Chen, C.; Yang, J.K. Angiotensin-converting enzyme 2 regulates mitochondrial function in pancreatic beta-cells. Biochem. Biophys. Res. Commun. 2018, 495, 860–866. [Google Scholar] [CrossRef]
- Wong, L.R.; Perlman, S. Immune dysregulation and immunopathology induced by SARS-CoV-2 and related coronaviruses—Are we our own worst enemy? Nat. Rev. Immunol. 2022, 22, 47–56. [Google Scholar] [CrossRef]
- Li, X.; Hou, P.; Ma, W.; Wang, X.; Wang, H.; Yu, Z.; Chang, H.; Wang, T.; Jin, S.; Wang, X.; et al. SARS-CoV-2 ORF10 suppresses the antiviral innate immune response by degrading MAVS through mitophagy. Cell. Mol. Immunol. 2022, 19, 67–78. [Google Scholar] [CrossRef]
- Holtz, E.D.; Azevedo-Quintaniha, I.G.; Palhinha, L.; Teixeira, L.; Barreto, E.A.; Pao, C.R.R.; Righy, C.; Franco, S.; Souza, T.M.L.; Kurtz, P.; et al. Platelet activation and platelet-monocyte aggregate formation trigger tissue factor expression in patients with severe COVID-19. Blood 2020, 136, 1330–1341. [Google Scholar]
- Singh, K.K.; Chaubey, G.; Chen, J.Y.; Suravajhala, P. Decoding SARS-CoV-2 hijacking of host mitochondria in COVID-19 pathogenesis. Am. J. Phys. Cell Phys. 2020, 319, C258–C267. [Google Scholar] [CrossRef]
- Gibellini, L.; De Biasi, S.; Paolini, A.; Borella, R.; Boraldi, F.; Mattioli, M.; Lo Tartaro, D.; Fidanza, L.; Caro-Maldonado, A.; Meschiari, M.; et al. Altered bioenergetics and mitochondrial dysfunction of monocytes in patients with COVID-19 pneumonia. EMBO Mol. Med. 2020, 12, e13001. [Google Scholar] [CrossRef] [PubMed]
- Ajaz, S.; McPhail, J.; Singh, K.K.; Mujib, S.; Trovato, F.M.; Napoli, S.; Agarwal, K. Mitochondrial metabolic manipulation by SARS-CoV-2 in peripheral blood mononuclear cells of patients with COVID-19. Am. J. Phys. Cell Phys. 2021, 320, C57–C65. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Amar, S.; Gehlot, P.; Patra, S.K.; Kanwar, N.; Kanwal, A. Mitochondrial modulations, autophagy pathways shifts in viral infections: Consequences of COVID-19. Int. J. Mol. Sci. 2021, 22, 8180. [Google Scholar] [CrossRef] [PubMed]
- Mantle, D.; Heaton, R.A.; Hargreaves, I.P. Coenzyme Q10 and immune functions: An overview. Antioxidants 2021, 10, 759. [Google Scholar] [CrossRef]
- Kawamukai, M. Biosynthesis of coenzyme Q in eukaryocytes. Biosci. Biotechnol. Biochem. 2016, 80, 23–33. [Google Scholar] [CrossRef]
- Hidalgo-Gutierrez, A.; Gonzalez-Garcia, P.; Diaz-Casado, M.E.; Bariocanal-Casado, E.; Lopez-Herrador, S.; Quinzii, C.M.; Lopez, L.C. Metabolic targets of coenzyme Q10 in mitochondria. Antioxidants 2021, 10, 520. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, M. Newly emerged antiviral strategies for SARS-CoV-2: From deciphering viral protein structural function to the development of vaccines, antibodies, and small molecules. Int. J. Mol. Sci. 2022, 23, 6083. [Google Scholar] [CrossRef]
- Zhou, X.; Jiang, X.; Qu, M.; Aninwene, G.E.; Jacaus, V.; Moon, J.J.; Gu, Z.; Sun, W.; Khademhosseini, A. Engineering antiviral vaccines. ACS Nano 2020, 14, 12370–12389. [Google Scholar] [CrossRef]
- Yagnik, D. Coenzyme Q10 and vitamin D interventions could ameliorate COVID-19 related cellular bioenergetic dysfunction and cytokine storms. J. Immunol. Sci. 2021, 5, 1–6. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Y. Potential interventions for novel coronavirus in China: A systematic review. J. Med. Virol. 2020, 92, 479–490. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Z.Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; You, Z.; Wang, Q.; Zhou, Z.J.; Qiu, Y.; Luo, R.; Ge, X.Y. The epidemic of 2019-novel-coronavirus (2019-nCoV) pneumonia and insight for emerging infectious diseases in the future. Microbes Infect. 2020, 22, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Sumbalova, Z.; Kucharska, J.; Palacka, P.; Rausova, Z.; Langsjoen, P.H.; Langsjoen, A.M.; Gvozdjakova, A. Platelet mitochondrial function and endogenous coenzyme Q10 levels are reduced in patients after COVID-19. Bratisl. Med. J. 2022, 123, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Sumbalova, Z.; Droescher, S.; Hiller, E.; Chang, S.C.; Garcia-Souza, L.F.; Calabria, E.; Volani, C.; Krumschnabel, G.; Gnaiger, E. O2k-Protocols: Isolation of peripheral blood mononuclear cells and platelets from human blood for HRFR. Mitochondr. Physiol. Netw. 2018, 3, 1–16. [Google Scholar]
- Palacka, P.; Gvozdjakova, A.; Rausova, Z.; Kucharska, J.; Slopovsky, J.; Furka, D.; Singh, K.K.; Sumbalova, Z. Platelet mitochondrial bioenergetics reprogramming in patients with urothelial carcinoma. Int. J. Mol. Sci. 2022, 23, 388. [Google Scholar] [CrossRef]
- Gvozdjakova, A.; Sumbalova, Z.; Kucharska, J.; Chladekova, A.; Rausova, Z.; Vančova, O.; Komlosi, M.; Ulična, O.; Mojto, V. Platelet mitochondrial bioenergetics analysis in patients with nephropathies and non-communicable diseases: A new method. Bratisl. Med. J. 2019, 12, 630–635. [Google Scholar] [CrossRef]
- Pesta, D.; Gnaiger, E. High-resolution respirometry: OXPHOS protocols for human cells and permeabilized fibers from small biopsies of human muscle. Methods Mol. Biol. 2012, 810, 25–58. [Google Scholar]
- Sjovall, F.; Ehinger, J.K.; Marelsson, S.E.; Morota, S.; Frostner, E.A.; Uchino, H.; Lundgren, J.; Arnbjörnsson, E.; Hansson, M.; Fellman, V.; et al. Mitochondrial respiration in human viable platelets—Methodology and influence gender, age and storage. Mitochondrion 2013, 13, 7–14. [Google Scholar] [CrossRef]
- Doerrier, C.; Sumbalova, Z.; Krumschnabel, G.; Hiller, E.; Gnaiger, E. SUIT reference protocol for OXPHOS analysis by high-resolution respirometry. Mitochondr. Physiol. Netw. 2016, 21, 1–12. [Google Scholar] [CrossRef]
- Lang, J.K.; Gohil, K.; Packer, L. Simultaneous determination of tocopherols, ubiquinols, and ubiquinones in blood, plasma, tissue homogenates, and subcellular fractions. Anal. Biochem. 1986, 157, 106–116. [Google Scholar] [CrossRef]
- Kucharska, J.; Gvozdjakova, A.; Mizera, S.; Braunova, Z.; Schreinerova, Z.; Schramekova, E.; Pechan, I.; Fabian, J. Participation of coenzyme Q10 in the rejection development of the transplanted heart. Physiol. Res. 1998, 47, 399–404. [Google Scholar] [PubMed]
- Mosca, F.; Fattorini, D.; Bompadre, S.; Littarru, G.P. Assay of coenzyme Q10 in plasma by a single dilution step. Anal. Biochem. 2002, 305, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Niklowitz, P.; Menke, T.; Andler, W.M.; Okun, J.G. Simultaneous analysis of coenzyme Q10 in plasma, erythrocytes and platelets: Comparison of the antioxidant level in blood cells and their enviroment in healthy children and after oral supplementation in adult. Clin. Chim. Acta 2004, 342, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Janero, D.R.; Bughardt, B. Thiobarbituric acid-reactive malondialdehyd formation during superoxide-dependent, iron-catalyzed lipid peroxidation: Influence of peroxidation conditions. Lipids 1989, 24, 125–131. [Google Scholar] [CrossRef] [PubMed]
- COVID-19 Excess Mortality Collaborators. Estimating excess mortality due to the COVID-19 pandemic: A systemic analysis of COVID-19-related mortality, 2020–2021. Meta-analysis. Lancet 2022, 399, 1513–1536. [Google Scholar] [CrossRef] [PubMed]
- Jimeno-Almazan, A.; Pallares, J.G.; Buendia-Romero, A.; Martinez-Cava, A.; Franco-Lopez, F.; Sanchez-Alcaraz, M.B.J.; Bernal-Morel, E.; Courel-Ibanez, J. Post-COVID-19 syndrome and the potential benefits of exercise. Int. J. Environ. Res. Public Health 2021, 18, 5329. [Google Scholar] [CrossRef]
- Burtscher, J.; Burtscher, M.; Millet, G.P. The central role of mitochondrial fitness on antiviral defences: An advocacy for the physical activity during the COVID-19 pandemic. Redox Biol. 2021, 43, 101976. [Google Scholar] [CrossRef]
- Jiang, Q. Natural form of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 46–90. [Google Scholar] [CrossRef]
- Fiorino, S.; Gallo, C.; Zippi, M.; Sabbatani, S.; Manfredi, R.; Moretti, R.; Fogacci, E.; Maggioli, K.; Loffredo, F.; Giampieri, E.; et al. Cytokine storm in aged people with CoV-2: Possible role of vitamins as therapy or preventive strategy. Aging Clin. Exp. Res. 2020, 32, 2115–2131. [Google Scholar] [CrossRef]
- Herman, B.; Wong, M.C.; Viwattanakulvanid, P. Vaccination status, favipiravir, and micronutrient supplementation roles in post-COVID symptoms: A longitudinal study. PLoS ONE 2022, 17, e0271385. [Google Scholar] [CrossRef]
- Jovic, T.H.; Ali, S.R.; Ibrahim, N.; Jessop, Z.M.; Tarassoli, S.P.; Dobbs, T.D.; Holford, P.; Thornton, C.A.; Whitaker, I.S. Could vitamins help in the fight against COVID-19? Nutrients 2020, 12, 2550. [Google Scholar] [CrossRef] [PubMed]
- Machhi, J.; Herskovitz, J.; Senan, A.M.; Dutta, D.; Nath, B.; Oleynikov, M.D.; Blombeg, W.R.; Meigs, D.D.; Hassa, M.; Patel, M.; et al. The natural history, pathobiology, and clinical manifestations of SARS-CoV-2 infections. J. Neuroimmune Pharmacol. 2020, 15, 359–386. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Gao, Y.; Liu, X.; Nie, Z.; Sun, H.; Lin, K.; Peng, H.; Wang, S. Identification and functional analysis of the SARS-CoV-2 nucleocapsid protein. BMC Microbiol. 2021, 21, 58. [Google Scholar] [CrossRef]
- Silvas, J.A.; Vasquez, D.M.; Park, J.G.; Chiem, K.; Allue-Guardia, A.; Garcia-Vilanova, A.; Platt, R.N.; Miorin, L.; Kehrer, T.; Cupic, A.; et al. Contribution of SARS-CoV-2 accessory proteins to viral pthogenicity in K18 human ACE2 transgenic mice. J. Virol. 2021, 95, e0040221. [Google Scholar] [CrossRef] [PubMed]
- Di Gennaro, F.; Pizzol, D.; Marotta, C.; Antunes, M.; Racalbuto, V.; Veronese, N.; Smith, L. Coronavirus diseases (COVID-19) current status and future perspectives: A narrative review. Int. J. Environ. Res. Public Health 2020, 17, 2690. [Google Scholar] [CrossRef] [PubMed]
- Archer, S.L.; Dasgupta, A.; Chen, K.H.; Wu, D.; Baid, K.; Mamtis, J.E.; Gonzales, V.; Read, A.; Bentley, R.E.T.; Martin, A.Y.; et al. SARS-CoV-2 mitochondriopathy in COVID-19 pneumonia exacerbates hypoxemia. Redox Biol. 2022, 58, 102508. [Google Scholar] [CrossRef]
- Lapuente-Brun, E.; Moreno-Loshuertos, R.; Acin-Perez, R.; Latorre-Pellicer, A.; Colas, C.; Balsa, E.; Perales-Clemente, E.; Quiros, P.M.; Calvo, E.; Rodriguez-Hernandez, M.A.; et al. Supercomplex assembly determines electron flux in the mitochondrial electron transport chain. Science 2013, 340, 1567–1570. [Google Scholar] [CrossRef]
- Scialo, F.; Fernandez-Ayala, D.J.; Sanz, A. Role of mitochondrial reverse electron transport in ROS signaling: Potential roles in health and disease. Front. Physiol. 2017, 8, 428. [Google Scholar] [CrossRef]
- Guzzi, P.H.; Mercatelli, D.; Ceraolo, C.; Giorgi, F.M. Master regulator analysis of the SARS-CoV-2/human interactome. J. Clin. Med. 2020, 9, 982. [Google Scholar] [CrossRef]
- Kucharska, J.; Sumbalova, Z.; Rausova, Z.; Palacka, P.; Navas, P.; Lopez-Lluch, G.; Kovalcikova, E.; Takacsova, T.; Gvozdjakova, A. Benefit of mountain spa rehabilitation and ubiquinol treatment in patients with post-COVID-19 syndrome. Bratisl. Med. J. 2023, 12, 89–96. [Google Scholar] [CrossRef]
- Caruso, F.; Rossi, M.; Pedersen, J.Z.; Incerpi, S. Computational studies reveal mechanism by which quinone derivatives can inhibit SARS-CoV-2. Study of embelin and two therapeutic compounds of interest, methyl prednisolone and dexamethasone. J. Infect. Public Health 2020, 13, 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chen, Y.; Tian, Z.; Zhu, M.; Zhang, B.; Du, S.; Li, Y.; Liu, Z.; Hou, S.; Yang, Y. Coenzyme Q10 attentuates human platelet aggregation induced by SARS-CoV-2 spike protien via reducing oxidative stress in vitro. Int. J. Mol. Sci. 2022, 23, 12345. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, M.; Norouzi, P.; Aazami, H.; Moosavi-Movahedi, A.A. Review on oxidative stress relation on COVID-19: Biomolecular and bioanalytical approach. Int. J. Biol. Macromol. 2021, 189, 802–818. [Google Scholar] [CrossRef]
- Çakırca, G.; Çakırca, T.D.; Üstünel, M.; Torun, A.; Koyuncu, I. Thiol level and total oxidant/antioxidant status in patients with COVID-19 infection. Ir. J. Med. Sci. 2021, 191, 1925–1930. [Google Scholar] [CrossRef]
- Žarkovic, N.; Orehovec, B.; Milkovic, L.; Baršic, B.; Tatzber, F.; Wonisch, W.; Tarle, M.; Kmet, M.; Mataic, A.; Jakovcevic, A.; et al. Preliminary findings on the association of the lipid peroxidation product 4-hydroxynonenal with the lethal outcome of aggressive COVID-19. Antioxidants 2021, 10, 1341. [Google Scholar] [CrossRef]
- Atanasovska, E.; Petrusevska, M.; Zendelovska, D.; Spasovska, K.; Stevanovikj, M.; Kasapinova, K.; Gjorgjievska, K.; Labachevski, N. Vitamin D levels and oxidative stress markers in patients hospitalized with COVID-19. Redox Rep. 2021, 26, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Tsermpini, E.E.; Glamočlij, U.; Ulucan-Karmak, F.; Trampuž, S.R.; Dolžan, V. Molecular mechanisms related to responses to oxidative stress and antioxidative therapies in COVID-19: A systematic review. Antioxidants 2022, 11, 1609. [Google Scholar] [CrossRef] [PubMed]
- Abdulla, K.A.; Um, C.Y.; Gross, M.D.; Bostick, R.M. Circulating γ-tocopherol concentrations are inversely associated with antioxidant exposures and directly associated with systemic oxidative stress and inflammation in adults. J. Nutr. 2018, 148, 1453–1461. [Google Scholar] [CrossRef]
- Stephensen, C.B.; Lietz, G. Vitamin A in resistance to and recovery from infection: Relevance to SARS-CoV2. Br. J. Nutr. 2021, 126, 1663–1672. [Google Scholar] [CrossRef]
- Lage, S.L.; Amaral, E.P.; Hilligan, K.L.; Laidlaw, E.; Rupert, A.; Namasivayan, S.; Rocco, J.; Galindo, F.; Kellogg, A.; Kumar, P.; et al. Persistent oxidative stress and inflammasome activation in CD14 high CD16 monocytes from COVID-19 Patients. Front. Immunol. 2022, 12, 799558. [Google Scholar] [CrossRef]
- Martín-Fernández, M.; Aller, R.; Heredia-Rodríguez, M.; Gómez-Sánchez, E.; Martínez-Paz, P.; Gonzalo-Benito, H.; Sánchez-de Prada, L.; Gorgojo, Ó.; Carnicero-Frutos, I.; Tamayo, E.; et al. Lipid peroxidation as a hallmark of severity in COVID-19 patients. Redox Biol. 2021, 48, 102181. [Google Scholar] [CrossRef] [PubMed]
- Cekerevac, I.; Turnic, T.N.; Draginic, N.; Andjic, M.; Zivkovic, V.; Simovic, S.; Susa, R.; Novkovic, L.; Mijailovic, Z.; Andjelkovic, M.; et al. Predicting severity and intrahospital mortality in COVID-19: The place and role of oxidative stress. Oxidative Med. Cell. Longev. 2021, 2021, 6615787. [Google Scholar] [CrossRef] [PubMed]
- Gvozdjáková, A.; Sumbalová, Z.; Kucharská, J.; Chládeková, A.; Rausová, Z.; Vančová, O.; Kubalová, M.; Kuzmiaková, Z.; Nemec, M.; Uličná, O.; et al. Platelets mitochondrial function depends on coenzyme Q10 concentration in human young, not in elderly subjects. J. Nutr. Ther. 2018, 7, 67–76. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| O2 Flow [pmol/s/106 Cells] | Control n = 16 | V + PAC19 n = 10 | p vs. Control | PAC19 n = 10 | p vs. Control |
|---|---|---|---|---|---|
| ce | 0.099 ± 0.008 | 0.092 ± 0.011 | 0.6 | 0.080 ± 0.007 | 0.13 |
| Dig | 0.002 ± 0.001 | 0.000 ± 0.000 | 0.13 | 0.000 ± 0.000 | 0.13 |
| 1PM | 0.021 ± 0.003 | 0.026 ± 0.002 | 0.18 | 0.013 ± 0.003 | 0.073 |
| 2D | 0.118 ± 0.015 | 0.108 ± 0.011 | 0.6 | 0.075 ± 0.008 | 0.034 |
| 2D;c | 0.129 ± 0.016 | 0.115 ± 0.012 | 0.5 | 0.089 ± 0.009 | 0.026 |
| 3U | 0.140 ± 0.017 | 0.119 ± 0.011 | 0.3 | 0.092 ± 0.010 | 0.039 |
| 4G | 0.161 ± 0.018 | 0.143 ± 0.013 | 0.5 | 0.095 ± 0.011 | 0.020 |
| 5S | 0.350 ± 0.027 | 0.327 ± 0.025 | 0.6 | 0.276 ± 0.012 | 0.071 |
| Control n = 16 | V + PAC19 n = 10 | p vs. Control | PAC19 n = 10 | p vs. Control | |
|---|---|---|---|---|---|
| Platelets (pmol/109 PLT) | |||||
| CoQ10 | 82.4 ± 5.4 | 65.6 ± 5.3 | 0.046 | 58.9 ± 3.6 | 0.004 |
| α-tocopherol | 3666.6 ± 382.1 | 4556.6 ± 620.4 | 0.2 | 1361.1 ± 161.2 | 0.0001 |
| γ-tocopherol | 173.1 ± 33.2 | 87.9 ± 17.7 | 0.051 | 190.5 ± 31.6 | 0.9 |
| Blood (µmol/L) | |||||
| CoQ10 | 0.306 ± 0.023 | 0.238 ± 0.031 | 0.085 | 0.217 ± 0.030 | 0.031 |
| α-tocopherol | 23.3 ± 1.55 | 21.0 ± 2.08 | 0.4 | 18.6 ± 1.99 | 0.085 |
| γ-tocopherol | 1.34 ± 0.24 | 1.07 ± 0.24 | 0.4 | 0.91 ± 0.13 | 0.2 |
| β-carotene | 0.273 ± 0.047 | 0.330 ± 0.053 | 0.4 | 0.302 ± 0.068 | 0.7 |
| Plasma (µmol/L) | |||||
| CoQ10 | 0.475 ± 0.038 | 0.318 ± 0.038 | 0.010 | 0.394 ± 0.043 | 0.2 |
| α-tocopherol | 34.9 ± 2.84 | 25.3 ± 1.79 | 0.021 | 29.4 ± 1.40 | 0.2 |
| γ-tocopherol | 2.17 ± 0.37 | 1.21 ± 0.19 | 0.067 | 1.50 ± 0.17 | 0.2 |
| β-carotene | 0.327 ± 0.054 | 0.364 ± 0.055 | 0.7 | 0.410 ± 0.077 | 0.4 |
| TBARS (µmol/L) | 5.04 ± 0.17 | 5.42 ± 0.24 | 0.2 | 4.62 ± 0.22 | 0.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gvozdjáková, A.; Kucharská, J.; Rausová, Z.; Lopéz-Lluch, G.; Navas, P.; Palacka, P.; Bartolčičová, B.; Sumbalová, Z. Effect of Vaccination on Platelet Mitochondrial Bioenergy Function of Patients with Post-Acute COVID-19. Viruses 2023, 15, 1085. https://doi.org/10.3390/v15051085
Gvozdjáková A, Kucharská J, Rausová Z, Lopéz-Lluch G, Navas P, Palacka P, Bartolčičová B, Sumbalová Z. Effect of Vaccination on Platelet Mitochondrial Bioenergy Function of Patients with Post-Acute COVID-19. Viruses. 2023; 15(5):1085. https://doi.org/10.3390/v15051085
Chicago/Turabian StyleGvozdjáková, Anna, Jarmila Kucharská, Zuzana Rausová, Guillermo Lopéz-Lluch, Plácido Navas, Patrik Palacka, Barbora Bartolčičová, and Zuzana Sumbalová. 2023. "Effect of Vaccination on Platelet Mitochondrial Bioenergy Function of Patients with Post-Acute COVID-19" Viruses 15, no. 5: 1085. https://doi.org/10.3390/v15051085
APA StyleGvozdjáková, A., Kucharská, J., Rausová, Z., Lopéz-Lluch, G., Navas, P., Palacka, P., Bartolčičová, B., & Sumbalová, Z. (2023). Effect of Vaccination on Platelet Mitochondrial Bioenergy Function of Patients with Post-Acute COVID-19. Viruses, 15(5), 1085. https://doi.org/10.3390/v15051085