

Molecular Analysis of the Global Population of Potato Virus S Redefines Its Phylogeny, and Has Crop Biosecurity Implications



Abstract
:1. Introduction
2. Materials and Methods
2.1. Surveys
2.2. RT-PCR and Sequencing
2.3. Recombination and Phylogenetic Analysis
2.4. Population Structure
3. Results
3.1. PVS Occurrence and Sequences
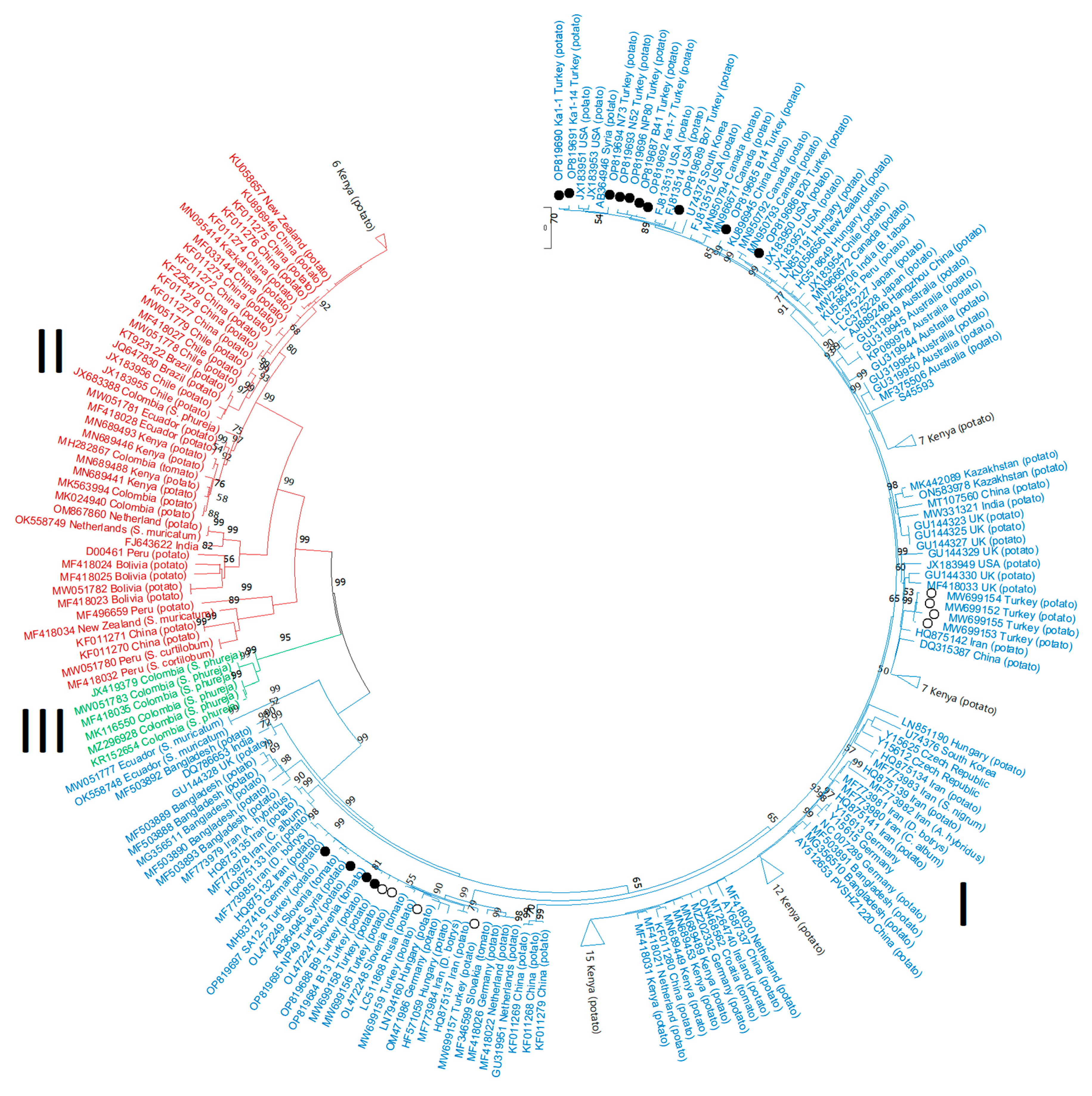
3.2. Recombination and Phylogenetic Analyses
3.3. Phylogenetic Analysis
3.4. Genetic Diversity and Selection Pressure Analyses
3.5. Neutrality Tests
3.6. Gene Flow and Genetic Separation amongst Populations
4. Discussion
4.1. Research Highlights
4.2. Incursion History
4.3. Recombination Findings
4.4. Phylogenetics
4.5. Population Genetics
4.6. Biosecurity Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wetter, C. AAB/CMI Descriptions of Plant Viruses, No. 60: Potato virus S; Association of Applied Biologists: Warwick, UK, 1971; p. 3. [Google Scholar]
- McKee, R.K. Virus infection in South American potatoes. Potato Res. 1964, 7, 145–151. [Google Scholar] [CrossRef]
- Stevenson, W.R.; Loria, R.; Franc, G.D.; Weingartner, D.P. Compendium of Potato Diseases, 2nd ed.; APS Press: St. Paul, MN, USA, 2001; p. 106. [Google Scholar]
- Jones, R.A.C. Virus disease problems facing potato industries worldwide: Viruses found, climate change implications, rationalizing virus strain nomenclature and addressing the Potato virus Y issue. In The Potato: Botany, Production and Uses; Navarre, R., Pavek, M.J., Eds.; CABI: Wallingford, UK, 2014; pp. 202–224. [Google Scholar]
- Duan, G.; Zhan, F.; Du, Z.; Ho, S.Y.; Gao, F. Europe was a hub for the global spread of potato virus S in the 19th century. Virology 2018, 525, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Santillan, F.W.; Fribourg, C.E.; Adams, I.P.; Gibbs, A.J.; Boonham, N.; Kehoe, M.A.; Maina, S.; Jones, R.A.C. The biology and phylogenetics of Potato virus S isolates from the Andean region of South America. Plant Dis. 2018, 102, 869–885. [Google Scholar] [CrossRef] [Green Version]
- Kreuze, J.F.; Souza-Dias, J.A.C.; Jeevalatha, A.; Figueira, A.R.; Valkonen, J.P.T.; Jones, R.A.C. Viral Diseases in Potato. In The Potato Crop; Campos, H., Ortiz, O., Eds.; Springer Cham: New York, NY, USA, 2020; pp. 389–430. [Google Scholar] [CrossRef] [Green Version]
- De Bokx, J.A.; van der Want, J.P.H. Viruses of Potatoes and Seed-Potato Production, 2nd ed.; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1987; p. 256. [Google Scholar]
- Jeffries, C. FAO/IPGRI Technical Guidelines for the Safe Movement of Germplasm; No. 19. Potato; Plant Genetic Resources Institute, Food and Agriculture Organization of the United Nations Rome/International: Rome, Italy, 1998; p. 177. [Google Scholar]
- Loebenstein, G.; Berger, P.H.; Brunt, A.A.; Lawson, R.H. (Eds.) Virus and Virus-Like Diseases of Potatoes and Production of Seed-Potatoes; Springer: Dordercht, The Netherlands, 2001; p. 460. [Google Scholar]
- Nyalugwe, E.P.; Wilson, C.R.; Coutts, B.A.; Jones, R.A.C. Biological properties of Potato virus X in potato: Effects of mixed infection with Potato virus S and resistance phenotypes in cultivars from three continents. Plant Dis. 2012, 96, 43–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martelli, G.P.; Adams, M.J.; Kreuze, J.F.; Dolja, V.V. Family Flexiviridae: A case study in virion and genome plasticity. Annu. Rev. Phytopathol. 2007, 45, 73–100. [Google Scholar] [CrossRef] [Green Version]
- Hinostroza-Orihuela, A.M. Some properties of potato virus S isolated from Peruvian potato varieties. Potato Res. 1973, 16, 244–250. [Google Scholar] [CrossRef]
- Jones, R.A.C. The ecology of viruses infecting wild and cultivated potatoes in the Andean Region of South America. In Pests, Pathogens and Vegetation; Thresh, J.M., Ed.; Pitman: London, UK, 1981; pp. 89–107. [Google Scholar]
- Foster, G.D.; Mills, P.R. The 3′-nucleotide sequence of an ordinary strain of potato virus S. Virus Genes 1992, 6, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Slack, S.A. Identification of an isolate of the Andean strain of potato virus S in North America. Plant Dis. 1983, 67, 786–789. [Google Scholar] [CrossRef]
- Matoušek, J.; Schubert, J.; Ptáček, J.; Kozlová, P.; Dědič, P. Complete nucleotide sequence and molecular probing of potato virus S genome. Acta Virol. 2005, 49, 195–205. [Google Scholar]
- Cox, B.A.; Jones, R.A.C. Genetic variability of the coat protein gene of Potato virus S and distinguishing its biologically distinct strains. Arch. Virol. 2010, 155, 1163–1169. [Google Scholar] [CrossRef]
- Gil, J.F.; Cotes, J.M.; Marín, M. Detection and molecular characterization of Potato virus S (PVS, Carlavirus) from Colombia. Rev. Biol. Trop. 2013, 61, 565–575. (In Spanish) [Google Scholar] [CrossRef]
- Gutiérrez, P.A.; Alzate, J.F.; Montoya, M.M. Pirosecuenciación del genoma de una cepa andina de Potato virus S (PVS, Carlavirus) infectando Solanum phureja (Solanaceae) en Colombia. Rev. Fac. Cienc Agrar 2012, 8, 84–93. (In Spanish) [Google Scholar] [CrossRef]
- Gutiérrez, P.A.; Alzate, J.F.; Marín-Montoya, M.A. Complete genome sequence of a novel potato virus S strain infecting Solanum phureja in Colombia. Arch. Virol. 2013, 158, 2205–2208. [Google Scholar] [CrossRef]
- Jones, R.A.C.; Kehoe, M.A. A proposal to rationalize within-species plant virus nomenclature: Benefits and implications of inaction/in silico characterization of coat protein of PVS-bitlis isolate and docking analysis with host protein. Arch. Virol. 2016, 161, 2051–2057. [Google Scholar] [CrossRef] [Green Version]
- Hanssen, I.M.; Thomma, B.P.H.J. Pepino mosaic virus: A successful pathogen that rapidly evolved from emerging to endemic in tomato crops. Mol. Plant Pathol. 2010, 11, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Moya, A.; Holmes, E.C.; González-Candelas, F. The population genetics and evolutionary epidemiology of RNA viruses. Nat. Rev. Microbiol. 2004, 2, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Coşkan, S.; Morca, A.F.; Akbaş, B.; Çelik, A.; Santosa, A.I. Comprehensive surveillance and population study on plum pox virus in Ankara Province of Turkey. J. Plant Dis. Prot. 2022, 129, 981–991. [Google Scholar] [CrossRef]
- Çelik, A.; Coşkan, S.; Morca, A.F.; Santosa, A.I.; Koolivand, D. Insight into population structure and evolutionary analysis of the emerging tomato brown rugose fruit virus. Plants 2022, 11, 3279. [Google Scholar] [CrossRef]
- TÜİK-Turkish Statistical Institute: Crop Production Statistic. 2021. Available online: https://www.tuik.gov.tr/ (accessed on 19 November 2022).
- Bostan, H.; Haliloglu, K. Distribution of PLRV, PVS, PVX and PVY (PVYN, PVYo and PVYc) in the seed potato tubers in Turkey. Pak. J. Biol. Sci. 2004, 7, 1140–1143. [Google Scholar] [CrossRef] [Green Version]
- Güner, Ü.; Yorganci, Ü. Niğde ve Nevşehir İlleri patates ekiliş alanlarında saptanan viral etmenler/Viral species detected in potato cultivation areas of Niğde and Nevşehir Provinces. Bitki Koruma Bülteni 2006, 46, 35–49. (In Turkish) [Google Scholar]
- Güner, Ü. Afyon, Bolu, Nevşehir ve Niğde Illerinde Patateslerdeki virüs Hastalıklarının Tanılanması, Hastalık Oranları ve en Yaygın Üretilen Çeşitlerin bazı Virüslere Karşı Reaksiyonlarının Belirlenmesi/Identification of Virus Diseases of Potato in Afyon, Bolu, Nevşehir, and Niğde Provinces, Disease Rates and Detection of the Most Common Grown Potato Varieties at the Variety Reactions through Some Plant Viruses. Ph.D. Thesis, Department of Plant Protection, Graduate School of Natural and Applied Sciences, Ege University, Izmir, Turkey, 2007. (In Turkish). [Google Scholar]
- Yardımcı, N.; Kılıç, H.C.; Demir, Y. Detection of PVY, PVX, PVS, PVA, and PLRV on different potato varieties in Turkey using DAS-ELISA. J. Agric. Sci. Technol. 2015, 17, 757–764. [Google Scholar]
- Bostan, H.; Guclu, C.; Ozturk, E.; Ozdemir, I.; Ilbagi, H. Influence of aphids on the epidemiology of potato virus diseases (PVY, PVS and PLRV) in the high-altitude areas of Turkey. Pak. J. Biol. Sci. 2006, 9, 759–765. [Google Scholar] [CrossRef] [Green Version]
- Kökten, M. Ödemiş bölgesinde üretimi yapılan patates yumrularında PVY, PVX, PVS ve PLRV’lerinin RT-PCR yöntemiyle saptanması/Determination of PVY, PVX, PVS and PLRV at potato tubers which were produced in Ödemiş region with RT-PCR method. Master’s Thesis, Graduate School of Natural and Applied Sciences, Adnan Menderes University, Aydın, Turkey, 2007. (In Turkish). [Google Scholar]
- Engür, A.M.; Topkaya, S. Prevalence and molecular characterization of important potato viruses in the Tokat province of Turkey. Mol. Biol. Rep. 2023, 50, 2171–2181. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, G.; Mustafa, U.; Demirel, S. Potato virus S (PVS)-Bitlis izolatının kılıf proteinin in silico karakterizasyonu ve konak proteini ile Docking analizi/In Silico characterization of coat protein of PVS-Bitlis isolate and docking analysis with host protein. J. Agric. 2022, 5, 58–69. (In Turkish) [Google Scholar] [CrossRef]
- Astruc, N.; Marcos, J.F.; Macquaire, G.; Candresse, T.; Pallás, V. Studies on the diagnosis of hop stunt viroid in fruit trees: Identification of new hosts and application of a nucleic acid extraction procedure based on non-organic solvents. Eur. J. Plant Pathol. 1996, 102, 837–846. [Google Scholar] [CrossRef]
- Salari, K.; Massumi, H.; Heydarnejad, J.; Pour, A.H.; Varsani, A. Analysis of Iranian Potato virus S isolates. Virus Genes 2011, 43, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Martin, D.P.; Varsani, A.; Roumagnac, P.; Botha, G.; Maslamoney, S.; Schwab, T.; Kelz, Z.; Kumar, V.; Murrell, B. RDP5: A computer program for analyzing recombination in, and removing signals of recombination from, nucleotide sequence datasets. Virus Evol. 2020, 7, veaa087. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Fu, Y.X.; Li, W.H. Statistical tests of neutrality of mutations. Genetics 1993, 133, 693–709. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Hudson, R.R.; Boos, D.D.; Kaplan, N.L. A statistical test for detecting population subdivision. Mol. Biol. Evol. 1992, 9, 138–151. [Google Scholar] [PubMed] [Green Version]
- Hudson, R.R. A new statistic for detecting genetic differentiation. Genetics 2000, 155, 2011–2014. [Google Scholar] [CrossRef] [PubMed]
- Çelik, A.; Santosa, A.I.; Gibbs, A.J.; Ertunç, F. Prunus necrotic ringspot virus in Turkey: An immigrant population. Arch. Virol. 2022, 167, 553–562. [Google Scholar] [CrossRef]
- Santosa, A.I.; Randa-Zelyüt, F.; Karanfil, A.; Korkmaz, S.; Hartono, S.; Ertunç, F. Phylogenetic and diversity analyses revealed that leek yellow stripe virus population consists of three types: S, L, and N. Virus Genes 2023, 59, 121–131. [Google Scholar] [CrossRef]
- Lin, Y.H.; Abad, J.A.; Maroon-Lango, C.J.; Perry, K.L.; Pappu, H.R. Molecular characterization of domestic and exotic potato virus S isolates and a global analysis of genomic sequences. Arch. Virol. 2014, 159, 2115–2122. [Google Scholar] [CrossRef]
- Bradshaw, J.E. Potato Breeding: Theory and Practice; Springer: Cham, Switzerland, 2021; p. 563. [Google Scholar] [CrossRef]
- Martin, R.R.; Constable, F.; Tzanetakis, I.E. Quarantine regulations and the impact of modern detection methods. Annu. Rev. Phytopathol. 2016, 54, 189–205. [Google Scholar] [CrossRef] [Green Version]
- Historical Data of Export and Import of Potatoes Seed, Fresh or Chilled. Available online: https://oec.world/en/profile/hs/potatoes-seed-fresh-or-chilled (accessed on 20 December 2022).
- Ge, B.; He, Z.; Zhang, Z.; Wang, H.; Li, S. Genetic variation in potato virus M isolates infecting pepino (Solanum muricatum) in China. Arch. Virol. 2014, 159, 3197–3210. [Google Scholar] [CrossRef]
- Santosa, A.I.; Ertunc, F. Phylogenetic and diversity analyses of Garlic common latent virus based on the TGB and CP gene sequence. Plant Prot. Sci. 2021, 57, 179–187. [Google Scholar] [CrossRef]
- Dolby, C.A.; Jones, R.A.C. Occurrence of the Andean strain of Potato virus S in imported potato material and its effects on potato cultivars. Plant Pathol. 1987, 36, 381–388. [Google Scholar] [CrossRef]
- Rose, D.G. Some properties of an unusual isolate of potato virus S. Potato Res. 1983, 26, 49–62. [Google Scholar] [CrossRef]
- Wardrop, E.A.; Gray, A.B.; Singh, R.P.; Peterson, J.F. Aphid transmission of potato virus S. Amer. Potato J. 1989, 66, 449–459. [Google Scholar] [CrossRef]
- Birch, P.R.; Bryan, G.; Fenton, B.; Gilroy, E.M.; Hein, I.; Jones, J.T.; Prashar, A.; Taylor, M.A.; Torrance, L.; Toth, I.K. Crops that feed the world 8: Potato: Are the trends of increased global production sustainable? Food Secur. 2012, 4, 477–508. [Google Scholar] [CrossRef]
- Scott, G.J. A review of root, tuber and banana crops in developing countries: Past, present and future. Int. J. Food Sci. Technol. 2021, 56, 1093–1114. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Province | No. of Samples Collected | No. of Samples with PVS | No. of PVS Isolates Sequenced | Isolate Name (CP Accession no.) and (PVSI Subclade) | Isolate Origins |
|---|---|---|---|---|---|
| Afyon | 85 | 16 | 6 | B13 (OP819684) (1) B9 (OP819688) (1) SA12-5 (OP819697) (1) B14 (OP819685) (3) B20 (OP819686) (5) B41 (OP819687) (4) | This study |
| Bolu | 71 | 8 | 1 | Bo7 (OP819689) (4) | This study |
| Kayseri | 35 | 3 | 3 | Ka1-1 (OP819690) (4) Ka1-7 (OP819692) (4) Ka1-14 (OP819691) (4) | This study |
| Nevşehir | 73 | 8 | 4 | N52 (OP819693) (4) N73 (OP819694) (4) NP80 (OP819696) (4) NP49 [OP819695] (1) | This study |
| Total | 264 | 35 | 14 | - | - |
| Tokat * | 418 | 70 | 8 | PA3-2 (MW699152) (2) PB5-4 (MW699153) (2) PB6-2 (MW699154) (2) PA7-1 (MW699155) (2) PN3-6 (MW699156) (1) PA3-3 (MW699157) (1) PN5-2 (MW699158) (1) PN14-3 (MW699159) (1) | [34] |
| No. | Recombinant | Parents: Major/Minor | Breakpoints 1 (Start/End) | RDP Implemented Method 2 (p Value) |
|---|---|---|---|---|
| 1. | AJ863510 (Czech Republic) | MZ202332 (PVSI, Germany)/MW0517582 (PVSII, Bolivia) | 18/6055 | R (3.287 × 10−58) G (1.037 × 10−53) B (1.387 × 10−57) M (8.144 × 10−33) C (1.961 × 10−31) S (9.952 × 10−96) 3S (7.898 × 10−167) |
| 2. | KC430335 (China) | MN689457 (PVSI, Kenya)/FJ813513 (PVSI, USA) | 4386/4814 | R (5.887 × 10−05) G (8.432 × 10−06) B (1.295 × 10−07) M (1.488 × 10−02) S (4.501 × 10−05) |
| 3. | LC375227 (Japan) | MK096268 (recombinant, China)/PVSI, LC375228 (Japan) | 6857/7184 | R (4.778 × 10−16) G (1.780 × 10−15) B (5.198 × 10−19) M (1.856 × 10−08) C (1.054 × 10−06) S (1.521 × 10−14) 3S (1.146 × 10−07) |
| 4. | LN851189 (Ukraine) | MN689477 (PVSI, Kenya)/OL472247 (PVSI, Slovenia) | 2626/8306 | R (1.571 × 10−24) G (3.017 × 10−19) B (4.610 × 10−25) M (1.825 × 10−21) C (1.712 × 10−02) S (5.127 × 10−37) 3S (1.298 × 10−25) |
| 5. | LN851192 (Ukraine) | OL472247 (PVSI, Slovenia)/MF418030 (PVSI, Netherlands) | 64/2751 | R (6.627 × 10−32) G (6.602 × 10−24) M (2.352 × 10−25) C (1.881 × 10−26) S (8.927 × 10−25) 3S (1.114 × 10−72) |
| 6. | LN851193 (Ukraine) | OL472247 (PVSI, Slovenia)/MF418030 (PVSI, Netherlands) | 64/2751 | R (6.627 × 10−32) G (6.602 × 10−24) M (2.352 × 10−25) C (1.881 × 10−26) S (8.927 × 10−25) 3S (1.114 × 10−72) |
| 7. | LN851194 (Poland) | MF418021 (PVSI, Netherlands)/LN851192 (recombinant, Ukraine) | 5910/7132 | R (6.083 × 10−26) G (1.320 × 10−26) B (6.687 × 10−29) M (1.101 × 10−12) C (2.457 × 10−12) S (4.668 × 10−15) 3S (8.473 × 10−32) |
| 8. | MN689463 (Kenya) | MN689494 (PVSI, Kenya)/MN689448 (PVSII, Kenya) | 21/311 | G (5.316 × 10−22) B (1.761 × 10−28) M (2.354 × 10−04) C (2.273 × 10−04) S (3.638 × 10−02) 3S (4.644 × 10−08) |
| 9. | MK096268 (China) | FJ813513 (PVSI, USA)/MK442089 (PVSI, Kazakhstan) | 6241/6926 | R (8.935 × 10−06) G (3.161 × 10−07) B (1.268 × 10−08) M (9.138 × 10−10) C (7.334 × 10−04) S (7.621 × 10−23) 3S (8.319 × 10−04) |
| Phylogroups | N | h | Hd | S | η | k | π | dS | dN | Ω |
|---|---|---|---|---|---|---|---|---|---|---|
| Full ORFs | 130 | 128 | 0.999 | 4133 | 6308 | 1063.591 | 0.1278 | 0.3607 | 0.0593 | 0.1644 |
| PVSI | 91 | 89 | 1.000 | 3489 | 4439 | 474.948 | 0.0571 | 0.1668 | 0.0247 | 0.1481 |
| PVSIII | 6 | 6 | 1.000 | 520 | 531 | 244.000 | 0.0293 | 0.0917 | 0.0109 | 0.1189 |
| PVSII | 33 | 33 | 1.000 | 2970 | 3724 | 784.453 | 0.0943 | 0.2775 | 0.0404 | 0.1456 |
| RdRP | 130 | 128 | 0.999 | 3045 | 4749 | 807.984 | 0.1363 | 0.4316 | 0.0502 | 0.1163 |
| PVSI | 91 | 89 | 1.000 | 2600 | 3334 | 359.116 | 0.0606 | 0.2028 | 0.0191 | 0.0942 |
| PVSIII | 6 | 6 | 1.000 | 411 | 420 | 190.533 | 0.0321 | 0.1111 | 0.0091 | 0.0819 |
| PVSII | 33 | 33 | 1.000 | 2245 | 2840 | 599.011 | 0.1011 | 0.3363 | 0.0324 | 0.0963 |
| TGB | 130 | 123 | 0.999 | 541 | 750 | 118.844 | 0.0991 | 0.2647 | 0.0483 | 0.1825 |
| PVSI | 91 | 84 | 0.998 | 453 | 559 | 59.845 | 0.0499 | 0.1302 | 0.0252 | 0.1935 |
| PVSIII | 6 | 6 | 1.000 | 45 | 48 | 23.133 | 0.0192 | 0.0513 | 0.0087 | 0.1696 |
| PVSII | 33 | 33 | 1.000 | 350 | 418 | 86.358 | 0.0719 | 0.1886 | 0.0364 | 0.1931 |
| CP | 224 | 210 | 0.999 | 435 | 687 | 93.011 | 0.1052 | 0.3759 | 0.0227 | 0.0603 |
| PVSI | 170 | 157 | 0.999 | 389 | 521 | 42.951 | 0.0486 | 0.1701 | 0.0115 | 0.0676 |
| PVSIII | 6 | 6 | 1.000 | 53 | 53 | 25.533 | 0.0288 | 0.1081 | 0.0046 | 0.0426 |
| PVSII | 49 | 47 | 0.998 | 304 | 408 | 72.423 | 0.0818 | 0.2914 | 0.0179 | 0.0614 |
| Phylogroups | Fu and Li’s D* | Fu and Li’s F* | Tajima’s D |
|---|---|---|---|
| Full ORFs | 0.58524 ns | 0.21631 ns | −0.27604 ns |
| PVSI | −0.80344 ns | −1.36717 ns | −1.57175 ns |
| PVSIII | 0.22861 ns | 0.27325 ns | 0.32038 ns |
| PVSII | 0.16750 ns | −0.10101 ns | −0.56069 ns |
| RdRp | 0.70023 ns | 0.30507 ns | −0.24821 ns |
| PVSI | −0.74118 ns | −1.31813 ns | −1.55871 ns |
| PVSIII | 0.14202 ns | 0.17776 ns | 0.23318 ns |
| PVSII | 0.21777 ns | −0.06014 ns | −0.55630 ns |
| TGB | −0.00180 ns | −0.25618 ns | −0.45833 ns |
| PVSI | −1.00886 ns | −1.49813 ns | −1.56128 ns |
| PVSIII | 0.63604 ns | 0.69883 ns | 0.64308 ns |
| PVSII | −0.05321 ns | −0.29591 ns | −0.62058 ns |
| CP | −0.45046 ns | −0.62638 ns | −0.60418 ns |
| PVSI | −2.07196 ns | −2.26514 ns | −1.71837 ns |
| PVSIII | 0.55376 ns | 0.62780 ns | 0.64148 ns |
| PVSII | −0.18097 ns | −0.47960 ns | −0.76064 ns |
| Comparison | αKS * | αKST * | p Value | αZ * | p Value | Snn | p Value | βFST |
|---|---|---|---|---|---|---|---|---|
| Complete ORFs | ||||||||
| PVSI (n = 91)/PVSIII (n = 6) | 5.9673 | 0.0329 | 0.0000 *** | 7.2663 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.7975 |
| PVSI (n = 91)/PVSII (n = 33) | 6.0726 | 0.0794 | 0.0000 *** | 7.4262 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.6394 |
| PVSIII (n = 6)/PVSII (n = 33) | 6.1672 | 0.0622 | 0.0000 *** | 5.2196 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.7019 |
| RdRp | ||||||||
| PVSI (n = 91)/PVSIII (n = 6) | 5.6939 | 0.0342 | 0.0000 *** | 7.2675 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.7966 |
| PVSI (n = 91)/PVSII (n = 33) | 5.7986 | 0.0828 | 0.0000 *** | 7.4271 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.6392 |
| PVSIII (n = 6)/PVSII (n = 33) | 5.9015 | 0.0641 | 0.0000 *** | 5.2236 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.6994 |
| TGB | ||||||||
| PVSI (n = 91)/PVSIII (n = 6) | 3.8905 | 0.0498 | 0.0000 *** | 7.2612 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.7981 |
| PVSI (n = 91)/PVSII (n = 33) | 3.9838 | 0.1061 | 0.0000 *** | 7.4409 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.6054 |
| PVSIII (n = 6)/PVSII (n = 33) | 3.9818 | 0.0957 | 0.0000 *** | 5.1959 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.7177 |
| CP | ||||||||
| PVSI (n = 170)/PVSIII (n = 6) | 3.6536 | 0.0291 | 0.0000 *** | 8.5398 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.7892 |
| PVSI (n = 170)/PVSII (n = 49) | 3.7281 | 0.1127 | 0.0000 *** | 8.6346 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.6669 |
| PVSIII (n = 6)/PVSII (n = 49) | 3.8668 | 0.0707 | 0.0000 *** | 6.0309 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.7005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Topkaya, Ş.; Çelik, A.; Santosa, A.I.; Jones, R.A.C. Molecular Analysis of the Global Population of Potato Virus S Redefines Its Phylogeny, and Has Crop Biosecurity Implications. Viruses 2023, 15, 1104. https://doi.org/10.3390/v15051104
Topkaya Ş, Çelik A, Santosa AI, Jones RAC. Molecular Analysis of the Global Population of Potato Virus S Redefines Its Phylogeny, and Has Crop Biosecurity Implications. Viruses. 2023; 15(5):1104. https://doi.org/10.3390/v15051104
Chicago/Turabian StyleTopkaya, Şerife, Ali Çelik, Adyatma Irawan Santosa, and Roger A. C. Jones. 2023. "Molecular Analysis of the Global Population of Potato Virus S Redefines Its Phylogeny, and Has Crop Biosecurity Implications" Viruses 15, no. 5: 1104. https://doi.org/10.3390/v15051104
APA StyleTopkaya, Ş., Çelik, A., Santosa, A. I., & Jones, R. A. C. (2023). Molecular Analysis of the Global Population of Potato Virus S Redefines Its Phylogeny, and Has Crop Biosecurity Implications. Viruses, 15(5), 1104. https://doi.org/10.3390/v15051104