
Transcription Factor Driven Gene Regulation in COVID-19 Patients


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Preparation
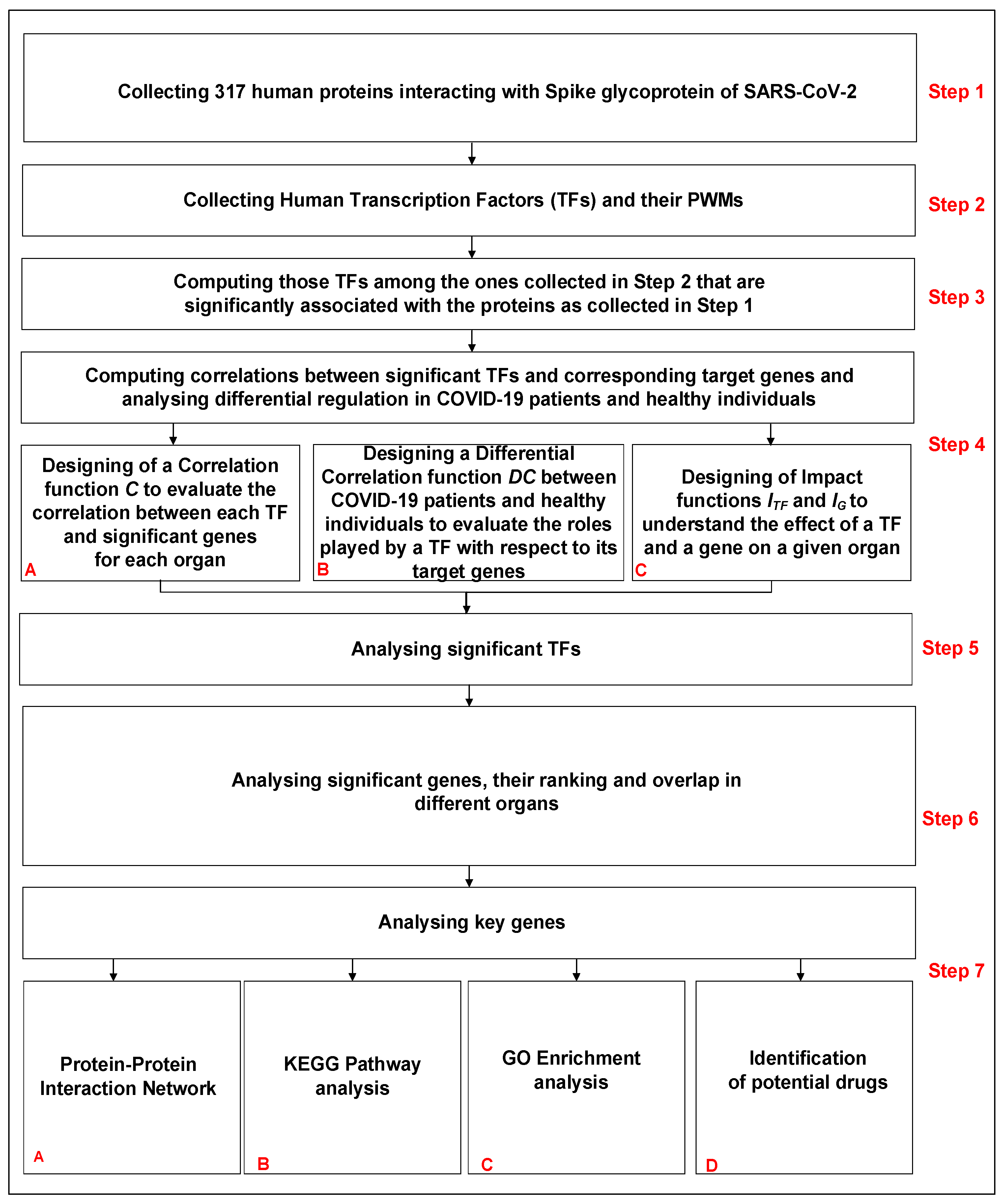
2.2. Pipeline of the Work
- Step 1: Collecting human proteins interacting with Spike glycoprotein of SARS-CoV-2.
- Step 2: Collecting human Transcription Factors and their PWMs.
- Step 3: Computing those TFs among the ones collected in Step 2 that are significantly associated with the proteins collected in Step 1.
- Step 4: Computing correlations between significant TFs (Step 3) and corresponding target genes and analysing differential regulation in COVID-19 patients and healthy individuals.
- Step 5: Analysing significant TFs identified in Step 3.
- Step 6: Analysing significant genes, their ranking and overlap in different organs.
- Step 7: Analysing key genes.
2.2.1. Identification of Significant TFs
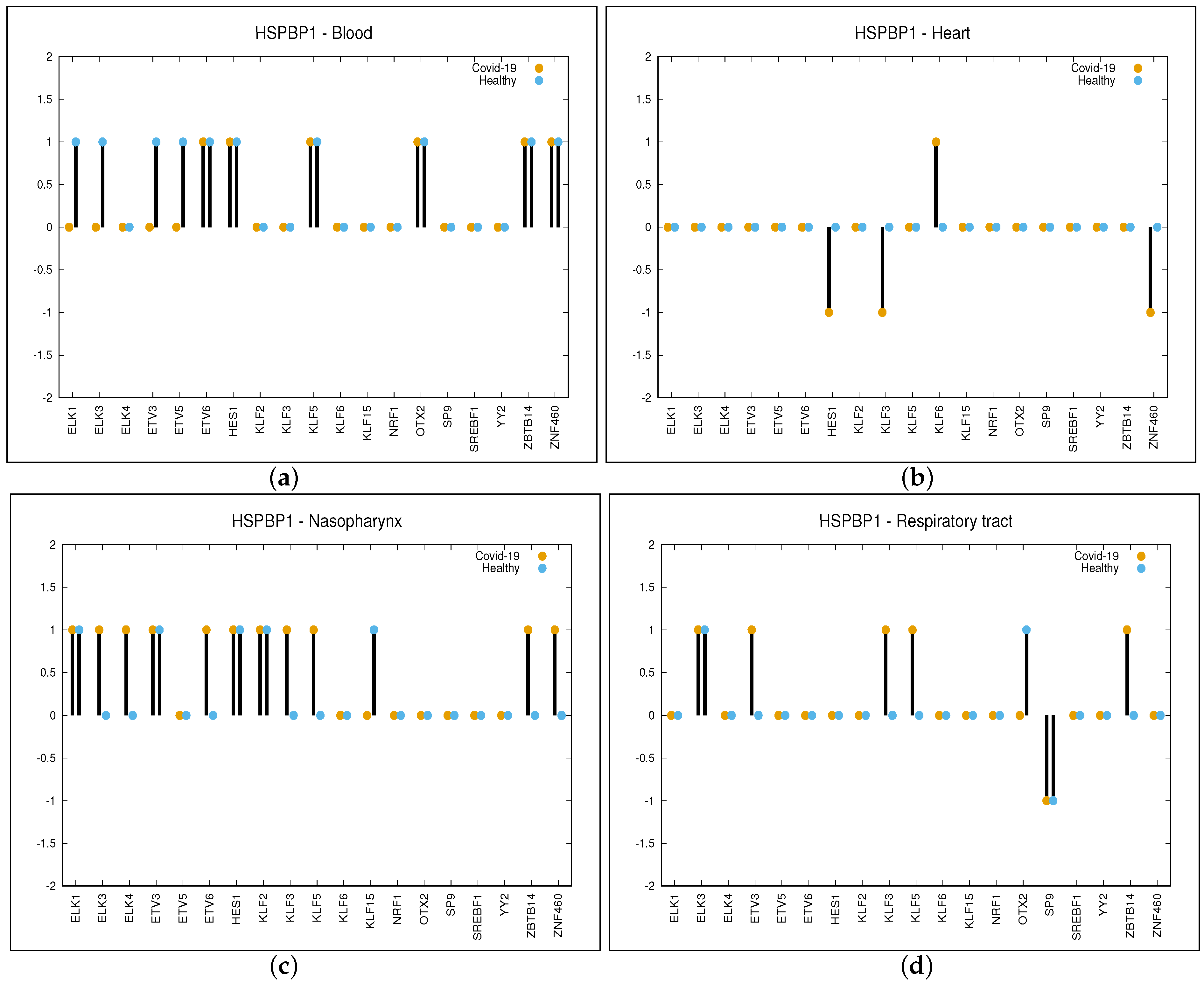
2.2.2. Gene—TF Correlations between COVID-19 Patients and Healthy Individuals
- C(tf,g,org,p) = −1, if g is a target gene of (at least one predicted TFBS associated with occurs in the promoter region of g), the Pearson correlation value between expression data of and g for the organ for condition p (COVID-19 or healthy) is negative and the associated p-value is smaller than .
- C(tf,g,org,p) = +1, if g is a target gene of (at least one predicted TFBS associated with occurs in the promoter region of g), the Pearson correlation value between expression data of and g for the organ for condition p (COVID-19 or healthy) is positive and the associated p-value is smaller than .
- C(tf,g,org,p) = 0, otherwise
- A strictly positive value of for the couple means that the given TF pushes up expression of gene G in COVID-19 patients while it does not do so in healthy individuals, or that the given TF pushes down expression of G in healthy individuals while it does not do so in COVID-19 patients.
- On the contrary, a strictly negative value of for the couple means that the given TF pushes up expression of gene G in healthy individuals while it does not do so in COVID-19 patients, or that the given TF pushes down expression of G in COVID-19 patients while the inverse is not true.
3. Results
3.1. Transcription Factors Significantly Associated with COVID-19 Infection
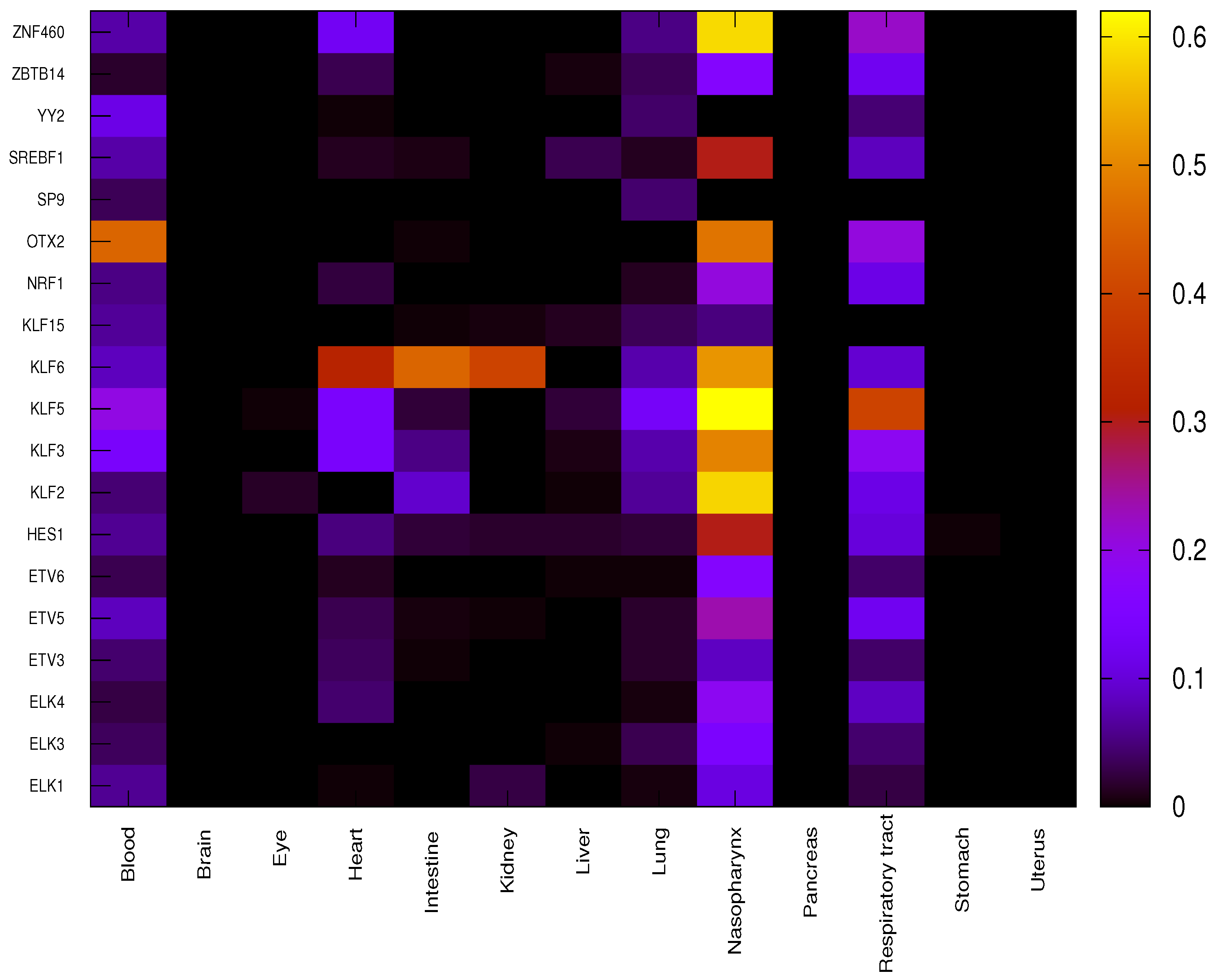
3.2. Transcription Factors Having a Major Impact on Specific Organs
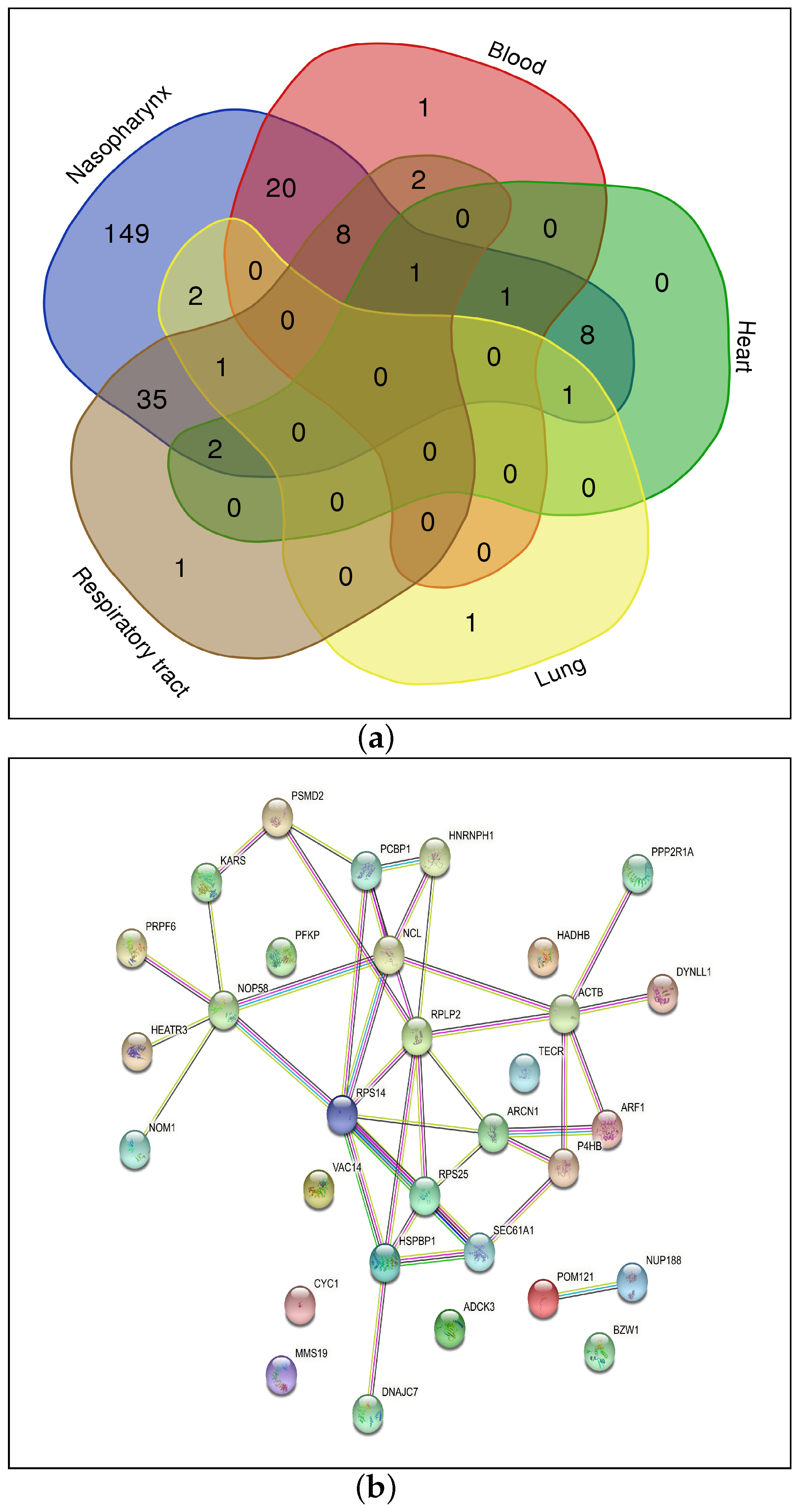
3.3. Differentially Regulated Genes in Specific Organs
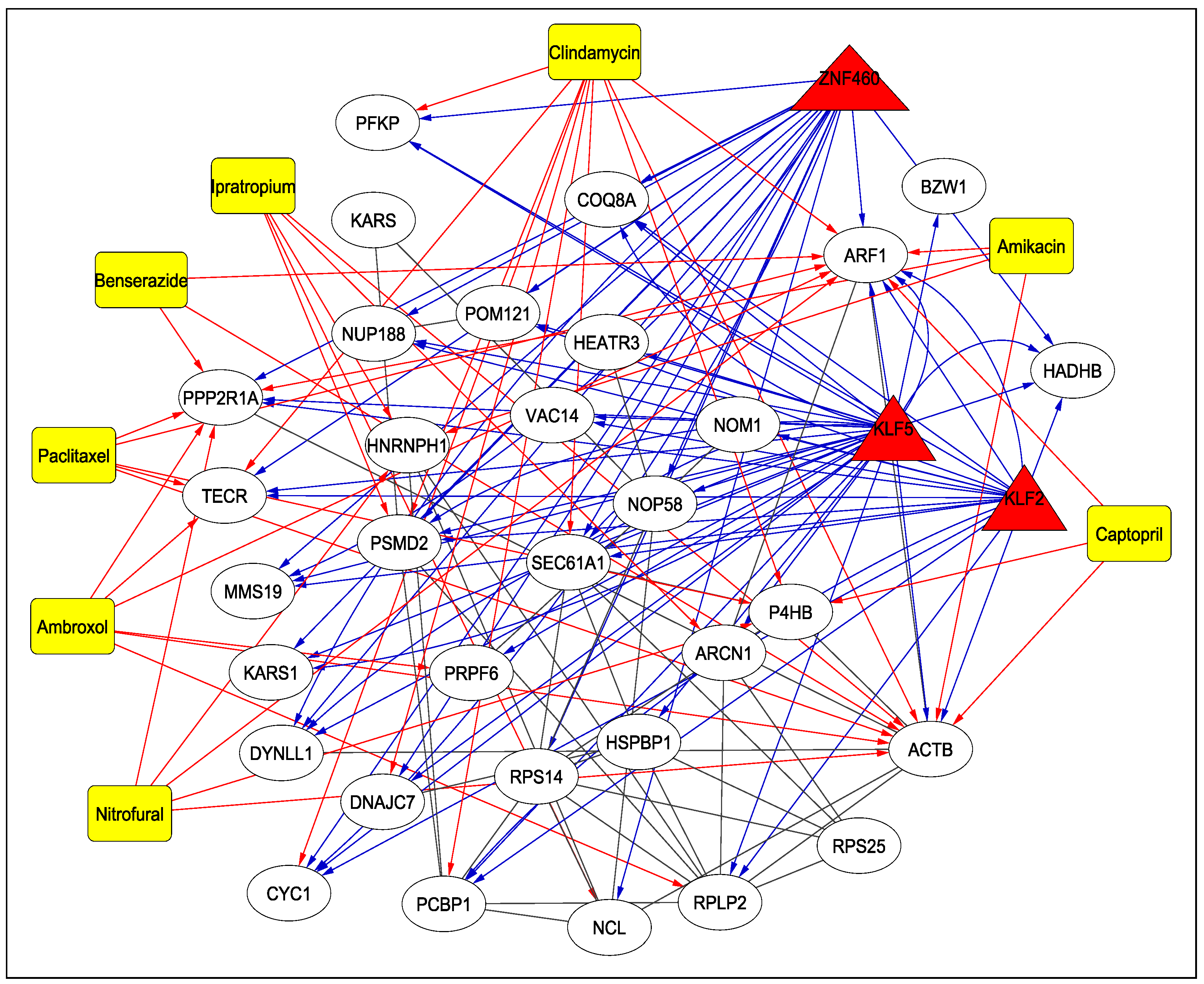
3.4. Drugs
4. Discussion
4.1. Protein–Protein Interaction Network and KEGG Pathway Analysis
4.2. Gene Ontology (GO) Enrichment Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TF | Transcription Factors |
| TFBS | Transcription Factor Binding Site |
| C | Correlation |
| DC | Differential Correlation |
| PWM | Position Weight Matrix |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| GO | Gene Ontology |
References
- Enjuanes, L.; Almazan, F.; Sola, I.; Zuniga, S. Biochemical Aspects of Coronavirus Replication and Virus-Host Interaction. Annu. Rev. Microbiol. 2006, 60, 211–230. [Google Scholar] [CrossRef] [PubMed]
- van Hemert, M.J.; van den Worm, S.H.E.; Knoops, K.; Mommaas, A.M.; Gorbalenya, A.E.; Snijder, E.J. SARS-Coronavirus Replication/Transcription Complexes Are Membrane-Protected and Need a Host Factor for Activity In Vitro. PLoS Pathog. 2008, 4, e1000054. [Google Scholar] [CrossRef] [PubMed]
- Vkovski, P.; Gerber, M.; Kelly, J.; Pfaender, S.; Ebert, N.; Braga, L.S.; Simillion, C.; Portmann, J.; Stalder, H.; Gaschen, V. Determination of host proteins composing the microenvironment of coronavirus replicase complexes by proximity-labeling. eLife 2019, 8, e42037. [Google Scholar] [CrossRef]
- di Bari, I.; Franzin, R.; Picerno, A.; Stasi, A.; Cimmarusti, M.T.; Di Chiano, M.; Curci, C.; Pontrelli, P.; Chironna, M.; Castellano, G.; et al. Severe acute respiratory syndrome coronavirus 2 may exploit human transcription factors involved in retinoic acid and interferon-mediated response: A hypothesis supported by an in silico analysis. New Microbes New Infect. 2021, 41, 100853. [Google Scholar] [CrossRef] [PubMed]
- Mosharaf, M.P.; Reza, M.S.; Kibria, M.K.; Ahmed, F.F.; Kabir, M.H.; Hasan, S.; Mollah, M.N.H. Computational identification of host genomic biomarkers highlighting their functions, pathways and regulators that influence SARS-CoV-2 infections and drug repurposing. Sci. Rep. 2022, 12, 4279. [Google Scholar] [CrossRef] [PubMed]
- Sardar, R.; Satish, D.; Gupta, D. Identification of Novel SARS-CoV-2 Drug Targets by Host MicroRNAs and Transcription Factors Co-regulatory Interaction Network Analysis. Front. Genet. 2020, 11, 1105. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Marishta, A.; Ellison, C.; Verzi, M.P. Identification of Transcription Factors Regulating SARS-CoV-2 Entry Genes in the Intestine. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 181–184. [Google Scholar] [CrossRef]
- Xu, S.; Liu, Y.; Ding, Y.; Luo, S.; Zheng, X.; Wu, X.; Liu, Z.; Ilyas, I.; Chen, S.; Han, S.; et al. The zinc finger transcription factor, KLF2, protects against COVID-19 associated endothelial dysfunction. Signal Transduct. Target. Ther. 2021, 6, 266. [Google Scholar] [CrossRef]
- Chapola, H.; de Bastiani, M.A.; Duarte, M.M.; Freitas, M.B.; Schuster, J.S.; de Vargas, D.M.; Klamt, F. A comparative study of COVID-19 transcriptional signatures between clinical samples and preclinical cell models in the search for disease master regulators and drug repositioning candidates. Virus Res. 2023, 326, 199053. [Google Scholar] [CrossRef]
- Ghosh, N.; Saha, I.; Sharma, N. Interactome of human and SARS-CoV-2 proteins to identify human hub proteins associated with comorbidities. Comput. Biol. Med. 2021, 138, 104889. [Google Scholar] [CrossRef]
- Chen, E.; Tan, C.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.; Jones, M.; Rouillard, A.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Derelitto, C.; Santoni, D. TRANSPARENT: A Python tool for designing transcription factor regulatory networks. Soft Comput. 2023, 27, 1–6. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, Y.; Min, Z.; Mo, J.; Ju, Z.; Guan, W.; Zeng, B.; Liu, Y.; Chen, J.; Zhang, Q.; et al. COVID19db: A comprehensive database platform to discover potential drugs and targets of COVID-19 at whole transcriptomic scale. Nucleic Acids Res. 2021, 50, D747–D757. [Google Scholar] [CrossRef] [PubMed]
- Cumbo, F.; Vergni, D.; Santoni, D. Investigating transcription factor synergism in humans. DNA Res. 2017, 25, 103–112. [Google Scholar] [CrossRef]
- Fornes, O.; Castro-Mondragon, J.A.; Khan, A.; Van der Lee, R.; Zhang, X.; Richmond, P.A.; Modi, B.P.; Correard, S.; Gheorghe, M.; Baranašić, D.; et al. JASPAR 2020: Update of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2019, 48, D87–D92. [Google Scholar] [CrossRef]
- Gao, L.; Li, G.S.; Li, J.D.; He, J.; Zhang, Y.; Zhou, H.F.; Kong, J.L.; Chen, G. Identification of the susceptibility genes for COVID-19 in lung adenocarcinoma with global data and biological computation methods. Comput. Struct. Biotechnol. J. 2021, 19, 6229–6239. [Google Scholar] [CrossRef]
- Melms, J.C.; Biermann, J.; Huang, H.; Wang, Y.; Nair, A.; Tagore, S.; Katsyv, I.; Rendeiro, A.F.; Amin, A.D.; Schapiro, D.; et al. A molecular single-cell lung atlas of lethal COVID-19. Nature 2021, 595, 114–119. [Google Scholar] [CrossRef]
- Chrysanthopoulou, A.; Antoniadou, C.; Natsi, A.M.; Gavriilidis, E.; Papadopoulos, V.; Xingi, E.; Didaskalou, S.; Mikroulis, D.; Tsironidou, V.; Kambas, K.; et al. Down-regulation of KLF2 in lung fibroblasts is linked with COVID-19 immunofibrosis and restored by combined inhibition of NETs, JAK-1/2 and IL-6 signaling. Clin. Immunol. 2023, 247, 109240. [Google Scholar] [CrossRef]
- Wen, W.; Su, W.; Tang, H.; Le, W.; Zhang, X.; Zheng, Y.; Liu, X.; Xie, L.; Li, J.; Ye, J.; et al. Immune cell profiling of COVID-19 patients in the recovery stage by single-cell sequencing. Cell Discov. 2020, 6, 31. [Google Scholar] [CrossRef]
- Hillary, V.E.; Ceasar, S.A. An update on COVID-19: SARS-CoV-2 variants, antiviral drugs, and vaccines. Heliyon 2023, 9, e13952. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.Z.; Zia, Q.; Haque, A.; Alqahtani, A.S.; Almarfadi, O.M.; Banawas, S.; Alqahtani, M.S.; Ameta, K.L.; Haque, S. Aminoglycosides as potential inhibitors of SARS-CoV-2 main protease: An in silico drug repurposing study on FDA-approved antiviral and anti-infection agents. J. Infect. Public Health 2021, 14, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, G.; Tang, Y.; Peng, Z.; Pan, H. The coronavirus diseases 2019 (COVID-19) pneumonia with spontaneous pneumothorax: A case report. BMC Infect. Dis. 2020, 20, 662. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Dong, H.Y.; Xie, H.Z.; Li, Y.M.; Jia, L. Why do we lack a specific magic anti-COVID-19 drug? Analyses and solutions. Drug Discov. Today 2021, 26, 631–636. [Google Scholar] [CrossRef]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Li, J.; Li, Q.; Xie, C.; Zhou, H.; Wang, Y.; Zhang, N.; Shao, H.; Chan, S.C.; Peng, X.; Lin, S.; et al. Beta-actin is required for mitochondria clustering and ROS generation in TNF-induced, caspase-independent cell death. J. Cell Sci. 2004, 117, 4673–4680. [Google Scholar] [CrossRef]
- Zhang, B.; Yu, Q.; Huo, D.; Li, J.; Liang, C.; Li, H.; Yi, X.; Xiao, C.; Zhang, D.; Li, M. Arf1 regulates the ER-mitochondria encounter structure (ERMES) in a reactive oxygen species-dependent manner. J. Cell Sci. 2018, 285, 2004–2018. [Google Scholar] [CrossRef]
- Brokatzky, D.; Hacker, G. Mitochondria: Intracellular sentinels of infections. Med. Microbiol. Immunol. 2022, 211, 161–172. [Google Scholar] [CrossRef]
- Lee, J.; Song, C. Effect of Reactive Oxygen Species on the Endoplasmic Reticulum and Mitochondria during Intracellular Pathogen Infection of Mammalian Cells. Antioxidants 2021, 10, 872. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GEOID | Organ | Number of Genes | COVID-19 | Healthy |
|---|---|---|---|---|
| GSE163151 | Blood | 21,952 | 145 | 113 |
| GSE164332 | Brain | 57,996 | 9 | 7 |
| GSE164073 | Eye | 25,222 | 9 | 9 |
| GSE162736 | Heart | 23,194 | 24 | 24 |
| GSE159201 | Intestine | 33,550 | 12 | 12 |
| GSE173707 | Kidney | 24,975 | 9 | 9 |
| GSE151803 | Liver | 22,316 | 12 | 9 |
| GSE147507 | Lung | 20,748 | 21 | 29 |
| GSE152075 | Nasopharynx | 19,744 | 430 | 54 |
| GSE165890 | Pancreas | 22,057 | 6 | 6 |
| GSE156063 | Respiratory tract | 15,811 | 93 | 141 |
| GSE153684 | Stomach | 26,501 | 9 | 9 |
| GSE171995 | Uterus | 19,715 | 5 | 3 |
| TF | Matrix ID–PWM | p-Value | Adjusted p-Value |
|---|---|---|---|
| KLF2 | MA1515.1 | ||
| KLF3 | MA1516.1 | ||
| KLF15 | MA1513.1 | ||
| KLF6 | MA1517.1 | ||
| NRF1 | MA0506.1 | 0.00016 | |
| SP9 | MA1564.1 | 0.00027 | |
| ZNF460 | MA1596.1 | 0.00061 | |
| ELK3 | MA0759.1 | 0.00088 | |
| HES1 | MA1099.2 | 0.00381 | |
| ZBTB14 | MA1650.1 | 0.00418 | |
| YY2 | MA0748.2 | 0.00845 | |
| ETV3 | MA0763.1 | 0.01290 | |
| ELK1 | MA0028.2 | 0.01493 | |
| OTX2 | MA0712.2 | 0.01594 | |
| ETV5 | MA0765.2 | 0.03179 | |
| ETV6 | MA0645.1 | 0.03611 | |
| KLF5 | MA0599.1 | 0.03843 | |
| SREBF1 | MA0829.2 | 0.03895 | |
| ELK4 | MA0076.2 | 0.04098 |
| Designation | Organs | Genes |
|---|---|---|
| 4* | Blood, Heart, Nasopharynx, Respiratory tract | HSPBP1 |
| 3* | Blood, Nasopharynx, Respiratory tract | P4HB |
| Blood, Nasopharynx, Respiratory tract | RPS14 | |
| Heart, Lung, Nasopharynx | ARCN1 | |
| Heart, Nasopharynx, Respiratory tract | PRPF6 | |
| Lung, Nasopharynx, Respiratory tract | BZW1 | |
| Blood, Nasopharynx, Respiratory tract | RPS25 | |
| Blood, Nasopharynx, Respiratory tract | NCL | |
| Blood, Nasopharynx, Respiratory tract | HEATR3 | |
| Blood, Heart, Nasopharynx | HNRNPH1 | |
| Blood, Nasopharynx, Respiratory tract | NOP58 | |
| 2* | Nasopharynx, Respiratory tract | ARF1 |
| Heart, Nasopharynx | POM121 | |
| Nasopharynx, Respiratory tract | COQ8A | |
| Nasopharynx, Respiratory tract | PCBP1 | |
| Nasopharynx, Respiratory tract | DYNLL1 | |
| Blood, Nasopharynx | TECR | |
| Nasopharynx, Respiratory tract | ACTB | |
| Nasopharynx, Respiratory tract | MMS19 | |
| Heart, Nasopharynx | PFKP | |
| Blood, Nasopharynx | HADHB | |
| 1* | Nasopharynx | PPP2R1A |
| Nasopharynx | RPLP2 | |
| Nasopharynx | NUP188 | |
| Nasopharynx | VAC14 | |
| Nasopharynx | NOM1 | |
| Nasopharynx | CYC1 | |
| Nasopharynx | SEC61A1 | |
| Nasopharynx | PSMD2 | |
| Nasopharynx | DNAJC7 | |
| Nasopharynx | KARS1 |
| Human Genes | Drugs | Adjusted p-Value | Drug Bank ID | Treatment |
|---|---|---|---|---|
| ARF1, PPP2R1A, HNRNPH1, P4HB, ACTB | Nitrofural | 0.0052 | DB00336 | Topical antibacterial for the prevention and treatment of bacterial infections of the skin |
| SEC61A1, ARF1, DNAJC7, PCBP1, PSMD2, TECR, CYC1, P4HB, ACTB, PFKP | Clindamycin | 0.0077 | DB01190 | Antibiotic used to treat serious infections caused by susceptible anaerobic, streptococcal, staphylococcal and pneumococcal bacteria |
| HNRNPH1, PSMD2, NCL, ACTB, ARCN1 | Ipratropium Bromide | 0.0101 | DB00332 | Used in the control of symptoms related to bronchospasm in chronic obstructive pulmonary disease (COPD) |
| ARF1, PRPF6, PPP2R1A, TECR, RPLP2, ACTB | Ambroxol | 0.0130 | DB06742 | Airway secretion clearance therapy |
| ARF1, PPP2R1A, TECR, P4HB, ACTB | Paclitaxel | 0.0133 | DB01229 | Treatment of advanced carcinoma of the ovary, and other various cancers including breast and lung cancer |
| ARF1, PPP2R1A, ACTB | Benserazide | 0.0236 | DB12783 | Treat Parkinson’s disease, Parkinsonism, and restless leg syndrome |
| ARF1, PPP2R1A, HNRNPH1, ACTB | Amikacin | 0.0351 | DB00479 | An aminoglycoside used to treat infections caused by more resistant strains of Gram-negative bacteria and some Gram-positive bacteria |
| ARF1, P4HB, ACTB | Captopril | 0.0420 | DB01197 | An ACE inhibitor used for the management of essential or renovascular hypertension, congestive heart failure, left ventricular dysfunction following myocardial infarction and nephropathy |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santoni, D.; Ghosh, N.; Derelitto, C.; Saha, I. Transcription Factor Driven Gene Regulation in COVID-19 Patients. Viruses 2023, 15, 1188. https://doi.org/10.3390/v15051188
Santoni D, Ghosh N, Derelitto C, Saha I. Transcription Factor Driven Gene Regulation in COVID-19 Patients. Viruses. 2023; 15(5):1188. https://doi.org/10.3390/v15051188
Chicago/Turabian StyleSantoni, Daniele, Nimisha Ghosh, Carlo Derelitto, and Indrajit Saha. 2023. "Transcription Factor Driven Gene Regulation in COVID-19 Patients" Viruses 15, no. 5: 1188. https://doi.org/10.3390/v15051188
APA StyleSantoni, D., Ghosh, N., Derelitto, C., & Saha, I. (2023). Transcription Factor Driven Gene Regulation in COVID-19 Patients. Viruses, 15(5), 1188. https://doi.org/10.3390/v15051188