
β-Glucan Induces Training Immunity to Promote Antiviral Activity by Activating TBK1

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Treatment of PMs with Different Stimuli
2.3. Mice
2.4. RNA Extraction, cDNA Synthesis, and Quantitative RT-PCR
2.5. ELISA
2.6. Western Blotting
2.7. Histopathology
2.8. Statistical Analysis
3. Results
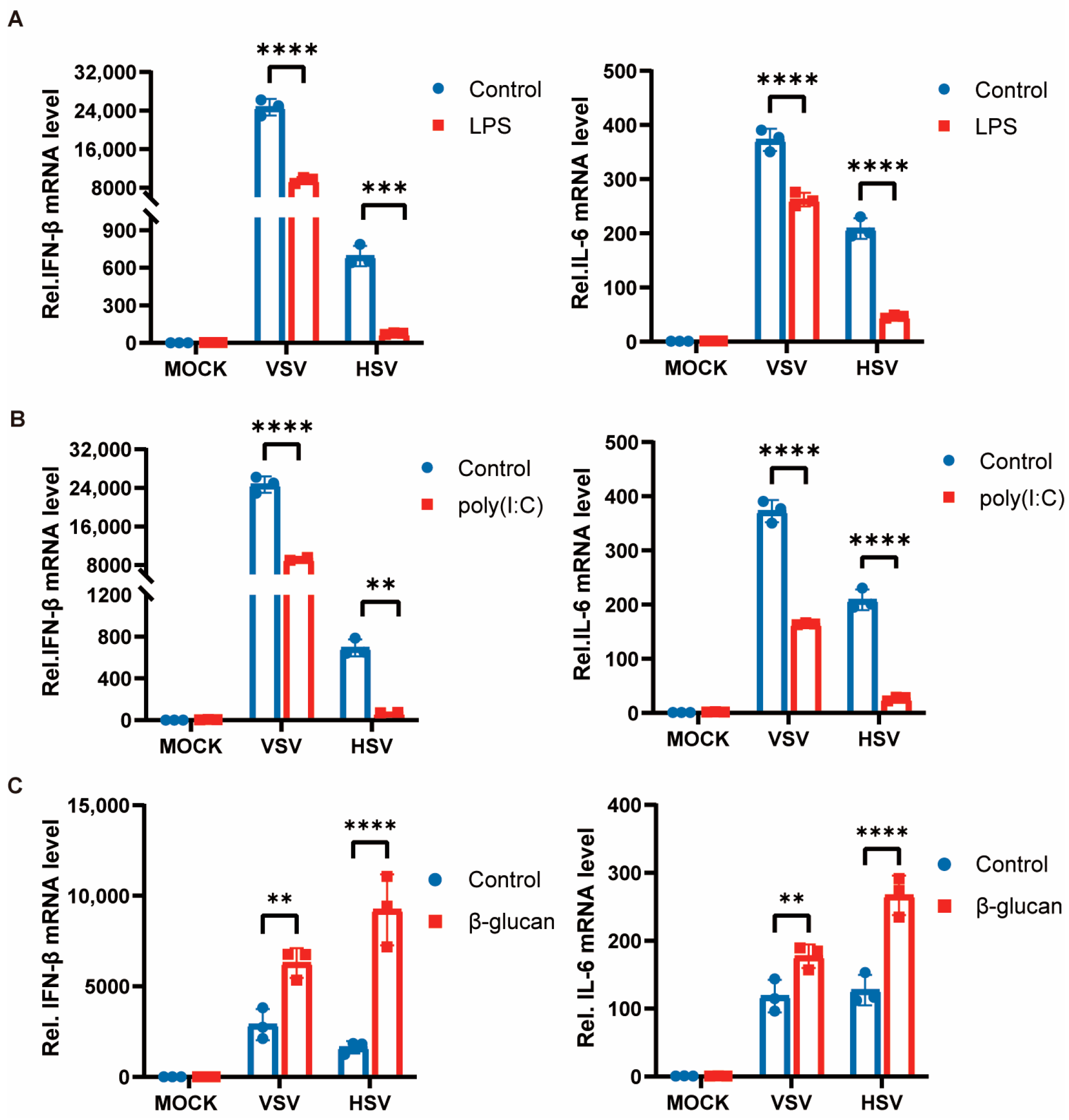
3.1. Trained Immunity Induced by β-Glucan Promotes Antiviral Immunity of Macrophages
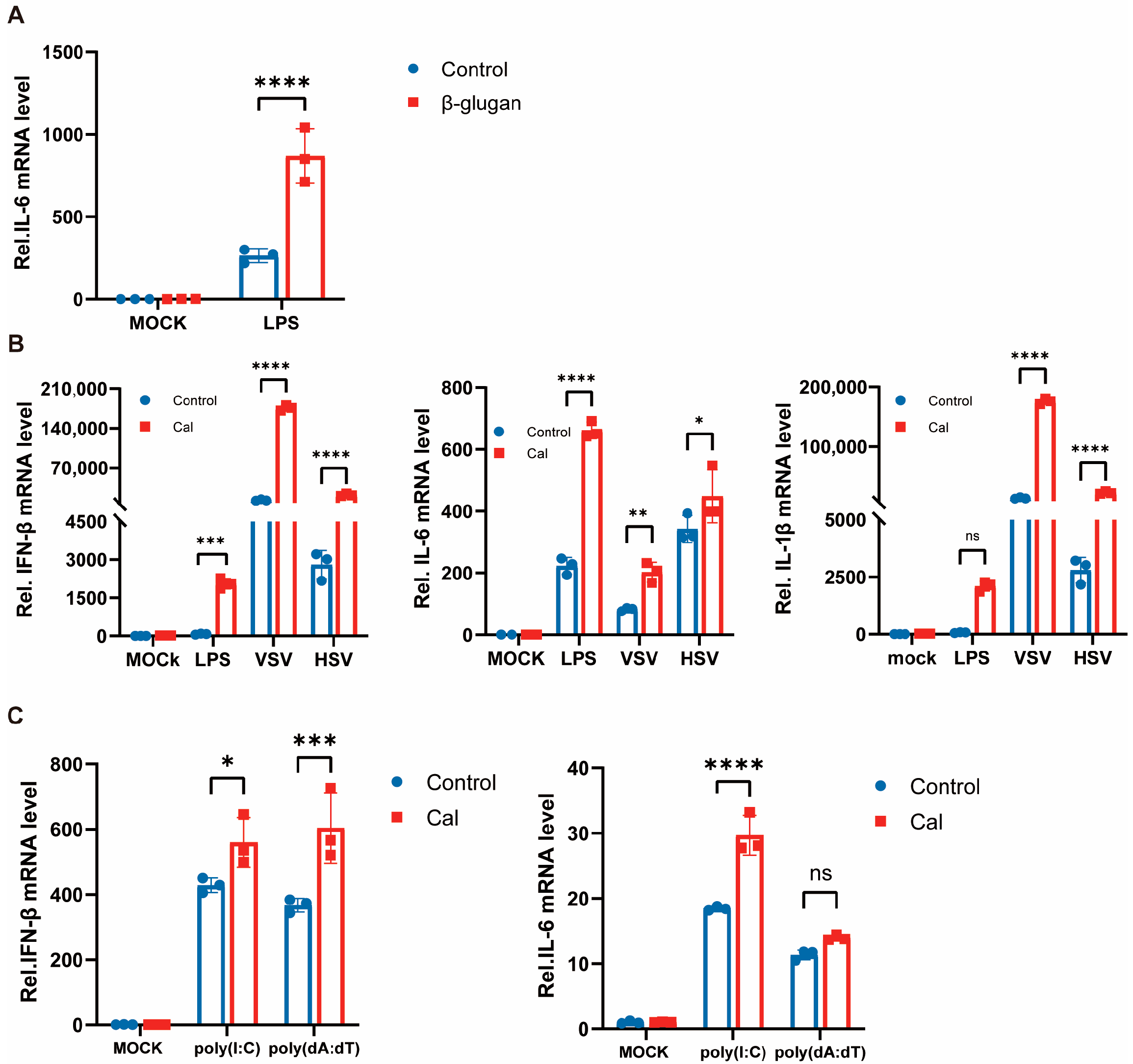
3.2. Trained Immunity Induced by Candida albicans Facilitates Antiviral Immunity of Macrophages
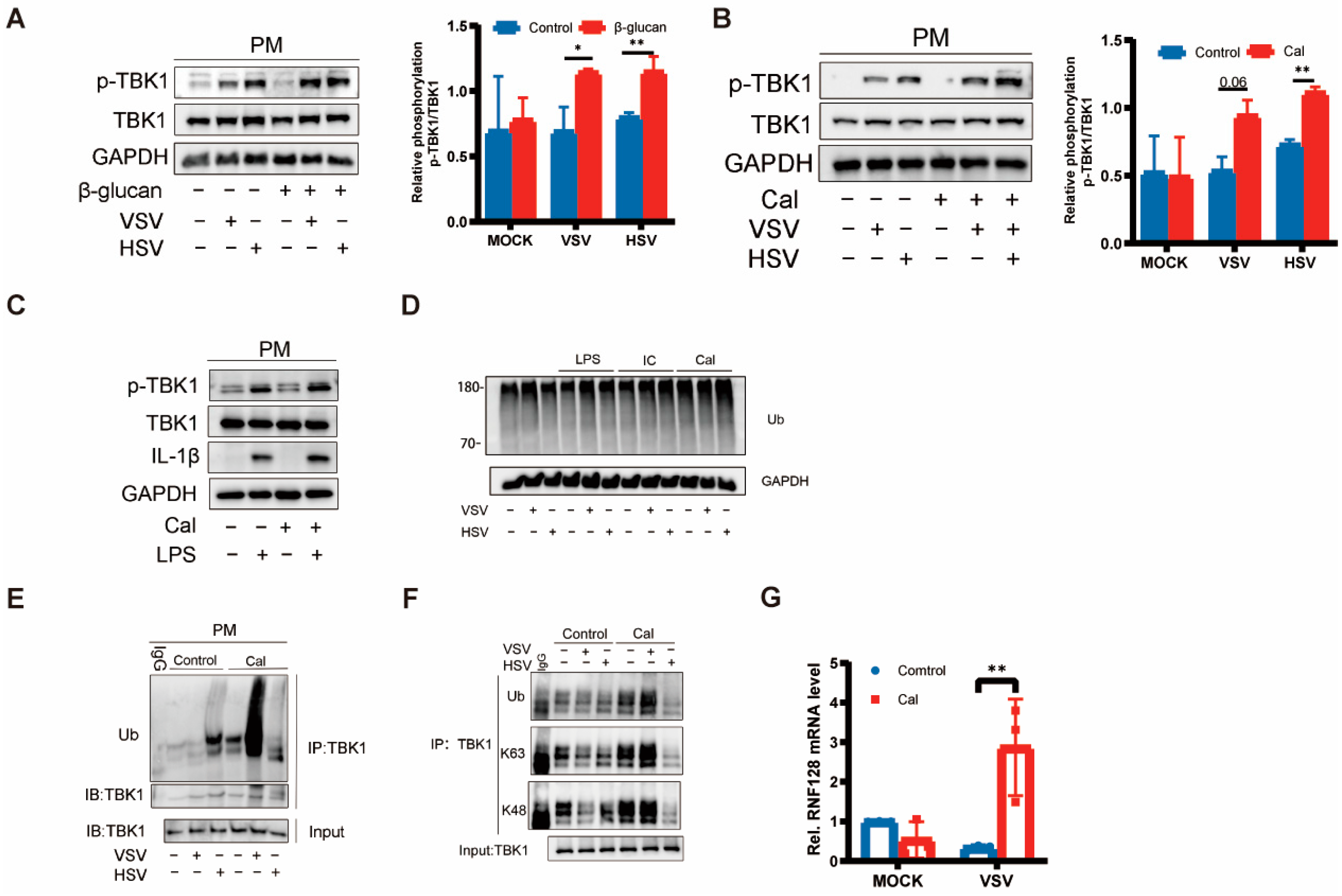
3.3. Trained Immunity Induced by β-Glucan Regulated TBK1 Activation
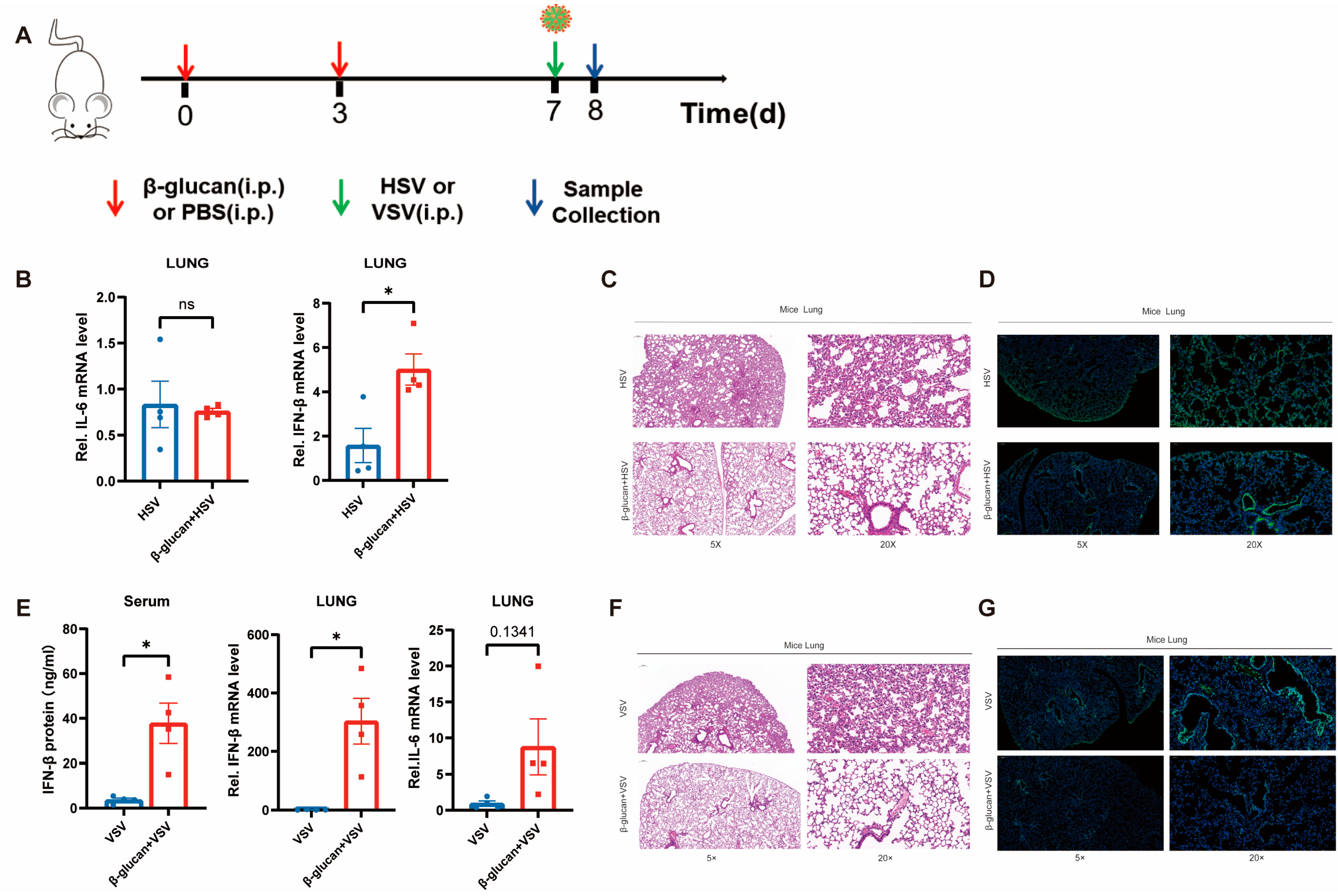
3.4. β-Glucan Enhances the Antiviral Innate Immunity in Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Netea, M.G.; Dominguez-Andres, J.; Barreiro, L.B.; Chavakis, T.; Divangahi, M.; Fuchs, E.; Joosten, L.A.B.; van der Meer, J.W.M.; Mhlanga, M.M.; Mulder, W.J.M.; et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 2020, 20, 375–388. [Google Scholar] [CrossRef]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.B.; Jacobs, C.; Xavier, R.J.; van der Meer, J.W.M.; van Crevel, R.; Netea, M.G. BCG-induced trained immunity in NK cells: Role for non-specific protec tion to infection. Clin. Immunol. 2014, 155, 213–219. [Google Scholar] [CrossRef]
- Quintin, J.; Saeed, S.; Martens, J.H.A.; Giamarellos-Bourboulis, E.J.; Ifrim, D.C.; Logie, C.; Jacobs, L.; Jansen, T.; Kullberg, B.-J.; Wijmenga, C.; et al. Candida albicans infection affords protection against reinfection via functional reprogramming of monocytes. Cell Host Microbe 2012, 12, 223–232. [Google Scholar] [CrossRef]
- Stothers, C.L.; Burelbach, K.R.; Owen, A.M.; Patil, N.K.; McBride, M.A.; Bohannon, J.K.; Luan, L.; Hernandez, A.; Patil, T.K.; Williams, D.L.; et al. β-Glucan Induces Distinct and Protective Innate Immune Memory in Differentiated Macrophages. J. Immunol. 2021, 207, 2785–2798. [Google Scholar] [CrossRef]
- Cheng, S.-C.; Quintin, J.; Cramer, R.A.; Shepardson, K.M.; Saeed, S.; Kumar, V.; Giamarellos-Bourboulis, E.J.; Martens, J.H.A.; Rao, N.A.; Aghajanirefah, A.; et al. mTOR- and HIF-1α-mediated aerobic glycolysis as metabolic basis for tr ained immunity. Science 2014, 345, 1250684. [Google Scholar] [CrossRef]
- de Laval, B.; Maurizio, J.; Kandalla, P.K.; Brisou, G.; Simonnet, L.; Huber, C.; Gimenez, G.; Matcovitch-Natan, O.; Reinhardt, S.; David, E.; et al. C/EBPβ-Dependent Epigenetic Memory Induces Trained Immunity in Hematop oietic Stem Cells. Cell Stem Cell 2020, 26, 657–674.e658. [Google Scholar] [CrossRef]
- Aegerter, H.; Kulikauskaite, J.; Crotta, S.; Patel, H.; Kelly, G.; Hessel, E.M.; Mack, M.; Beinke, S.; Wack, A. Influenza-induced monocyte-derived alveolar macrophages confer prolong ed antibacterial protection. Nat. Immunol. 2020, 21, 145–157. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef]
- Song, G.; Liu, B.; Li, Z.; Wu, H.; Wang, P.; Zhao, K.; Jiang, G.; Zhang, L.; Gao, C. E3 ubiquitin ligase RNF128 promotes innate antiviral immunity through K63-linked ubiquitination of TBK1. Nat. Immunol. 2016, 17, 1342–1351. [Google Scholar] [CrossRef]
- Li, L.; Luo, J.; Zhu, Z.; Xu, Q.; Wang, P.; Chang, B.; Wang, D.; Yu, L.; Lu, X.; Zhou, J.; et al. SRA Suppresses Antiviral Innate Immune Response in Macrophages by Limi ting TBK1 K63 Ubiquitination via Deubiquitinase USP15. Microbiol. Spectr. 2022, 10, e0202822. [Google Scholar] [CrossRef]
- Zhang, X.; Goncalves, R.; Mosser, D.M. The isolation and characterization of murine macrophages. Curr. Protoc. Immunol. 2008, 14, 14.11.11–14.11.14. [Google Scholar] [CrossRef]
- Netea, M.G.; Brown, G.D.; Kullberg, B.J.; Gow, N.A.R. An integrated model of the recognition of Candida albicans by the inna te immune system. Nat. Rev. Microbiol. 2008, 6, 67–78. [Google Scholar] [CrossRef]
- Tanaka, Y.; Chen, Z.J. STING specifies IRF3 phosphorylation by TBK1 in the cytosolic DNA sign aling pathway. Sci. Signal. 2012, 5, ra20. [Google Scholar] [CrossRef]
- Tsukamoto, H.; Takeuchi, S.; Kubota, K.; Kobayashi, Y.; Kozakai, S.; Ukai, I.; Shichiku, A.; Okubo, M.; Numasaki, M.; Kanemitsu, Y.; et al. Lipopolysaccharide (LPS)-binding protein stimulates CD14-dependent Tol l-like receptor 4 internalization and LPS-induced TBK1-IKKϵ-IRF3 axis activation. J. Biol. Chem. 2018, 293, 10186–10201. [Google Scholar] [CrossRef]
- Bo, Z.; Yan, Y.; Shu, L.; Wang, Y.Y.; Ying, L.; Diao, F.; Lei, C.; Xiao, H.; Lu, Z.; Tien, P. The adaptor protein MITA links virus-sensing receptors to IRF3 transcription factor activation. Immunity 2008, 29, 538–550. [Google Scholar]
- Liu, S.; Cai, X.; Wu, J.; Cong, Q.; Chen, X.; Li, T.; Du, F.; Ren, J.; Wu, Y.-T.; Grishin, N.V.; et al. Phosphorylation of innate immune adaptor proteins MAVS, STING, and TRI F induces IRF3 activation. Science 2015, 347, aaa2630. [Google Scholar] [CrossRef]
- Novakovic, B.; Habibi, E.; Wang, S.Y.; Arts, R.J.W.; Davar, R.; Megchelenbrink, W.; Kim, B.; Kuznetsova, T.; Kox, M.; Zwaag, J.; et al. beta-Glucan Reverses the Epigenetic State of LPS-Induced Immunological Tolerance. Cell 2016, 167, 1354–1368.e1314. [Google Scholar] [CrossRef]
- Tu, D.; Zhu, Z.; Zhou, A.Y.; Yun, C.-h.; Lee, K.-E.; Toms, A.V.; Li, Y.; Dunn, G.P.; Chan, E.; Thai, T.; et al. Structure and ubiquitination-dependent activation of TANK-binding kinase 1. Cell Rep. 2013, 3, 747–758. [Google Scholar] [CrossRef]
- Rice, P.J.; Adams, E.L.; Ozment-Skelton, T.; Gonzalez, A.J.; Goldman, M.P.; Lockhart, B.E.; Barker, L.A.; Breuel, K.F.; Deponti, W.K.; Kalbfleisch, J.H.; et al. Oral delivery and gastrointestinal absorption of soluble glucans stimu late increased resistance to infectious challenge. J. Pharmacol. Exp. Ther. 2005, 314, 1079–1086. [Google Scholar] [CrossRef]
- Ciarlo, E.; Heinonen, T.; Théroude, C.; Asgari, F.; Le Roy, D.; Netea, M.G.; Roger, T. Trained Immunity Confers Broad-Spectrum Protection Against Bacterial I nfections. J. Infect. Dis. 2020, 222, 1869–1881. [Google Scholar] [CrossRef]
- Shi, S.; Yin, L.; Shen, X.; Dai, Y.; Wang, J.; Yin, D.; Zhang, D.; Pan, X. β-Glucans from Trametes versicolor (L.) Lloyd Is Effective for Prevention of Influenza Virus Infection. Viruses 2022, 14, 237. [Google Scholar] [CrossRef]
- Mitroulis, I.; Ruppova, K.; Wang, B.; Chen, L.-S.; Grzybek, M.; Grinenko, T.; Eugster, A.; Troullinaki, M.; Palladini, A.; Kourtzelis, I.; et al. Modulation of Myelopoiesis Progenitors Is an Integral Component of Tra ined Immunity. Cell 2018, 172, 147–161.e112. [Google Scholar] [CrossRef]
- Saeed, S.; Quintin, J.; Kerstens, H.H.; Rao, N.A.; Aghajanirefah, A.; Matarese, F.; Cheng, S.C.; Ratter, J.; Berentsen, K.; van der Ent, M.A.; et al. Epigenetic programming of monocyte-to-macrophage differentiation and trained innate immunity. Science 2014, 345, 1251086. [Google Scholar] [CrossRef]
- Esteban, A.; Popp, M.W.; Vyas, V.K.; Strijbis, K.; Ploegh, H.L.; Fink, G.R. Fungal recognition is mediated by the association of dectin-1 and galectin-3 in macrophages. Proc. Natl. Acad. Sci. USA 2011, 108, 14270–14275. [Google Scholar] [CrossRef]
- Zhou, R.; Zhang, Q.; Xu, P. TBK1, a central kinase in innate immune sensing of nucleic acids and b eyond. Acta Biochim. Biophys. Sin. 2020, 52, 757–767. [Google Scholar] [CrossRef]
- Ma, X.; Helgason, E.; Phung, Q.T.; Quan, C.L.; Iyer, R.S.; Lee, M.W.; Bowman, K.K.; Starovasnik, M.A.; Dueber, E.C. Molecular basis of Tank-binding kinase 1 activation by transautophosph orylation. Proc. Natl. Acad. Sci. USA 2012, 109, 9378–9383. [Google Scholar] [CrossRef]
- Deng, M.; Tam, J.W.; Wang, L.; Liang, K.; Li, S.; Zhang, L.; Guo, H.; Luo, X.; Zhang, Y.; Petrucelli, A.; et al. TRAF3IP3 negatively regulates cytosolic RNA induced anti-viral signali ng by promoting TBK1 K48 ubiquitination. Nat. Commun. 2020, 11, 2193. [Google Scholar] [CrossRef]
- Liang, H.; Li, Y.; Li, M.; Zhou, W.; Chen, J.; Zhang, Z.; Yang, Y.; Ran, C.; Zhou, Z. The effect and underlying mechanism of yeast β-glucan on antiviral res istance of zebrafish against spring viremia of carp virus infection. Front. Immunol. 2022, 13, 1031962. [Google Scholar] [CrossRef]
- Vetvicka, V.; Vetvickova, J. Glucan supplementation enhances the immune response against an influen za challenge in mice. Ann. Transl. Med. 2015, 3, 22. [Google Scholar] [CrossRef]
- Murphy, E.J.; Masterson, C.; Rezoagli, E.; O’Toole, D.; Major, I.; Stack, G.D.; Lynch, M.; Laffey, J.G.; Rowan, N.J. β-Glucan extracts from the same edible shiitake mushroom Lentinus edod es produce differential in-vitro immunomodulatory and pulmonary cytopr otective effects—Implications for coronavirus disease (COVID-19) imm unotherapies. Sci. Total Environ. 2020, 732, 139330. [Google Scholar] [CrossRef]
- Basso, A.M.M.; De Castro, R.J.A.; de Castro, T.B.; Guimarães, H.I.; Polez, V.L.P.; Carbonero, E.R.; Pomin, V.H.; Hoffmann, C.; Grossi-de-Sa, M.F.; Tavares, A.H.; et al. Immunomodulatory activity of β-glucan-containing exopolysaccharides fr om Auricularia auricular in phagocytes and mice infected with Cryptoco ccus neoformans. Med. Mycol. 2020, 58, 227–239. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′ → 3′) | Reverse (5′ → 3′) |
|---|---|---|
| m-IL-6 | TTCCATCCAGTTGCCTTCTTG | AATTAAGCCTCCGACTTGTGAA |
| m-IFN-β | AGATCAACCTCACCTACAGG | TCAGAAACACTGTCTGCTGG |
| m-RNF128 | AGCTTCCATAATAAACACCT | CCTTTAACTGCCTCTGTAATAA |
| m-GAPDH | GAA GGG CTC ATG ACC ACA GT | GGA TGCAGG GAT GAT GTT CT |
| m-IL-1β | GTTCCCATTAGACAACTGCACTACAG | GTCGTTGCTTGGTTCTCCTTGTA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Li, Z.; Tian, M.; Cui, X.; Ma, J.; Liu, S.; Ye, C.; Yuan, L.; Qudus, M.S.; Afaq, U.; et al. β-Glucan Induces Training Immunity to Promote Antiviral Activity by Activating TBK1. Viruses 2023, 15, 1204. https://doi.org/10.3390/v15051204
Wang G, Li Z, Tian M, Cui X, Ma J, Liu S, Ye C, Yuan L, Qudus MS, Afaq U, et al. β-Glucan Induces Training Immunity to Promote Antiviral Activity by Activating TBK1. Viruses. 2023; 15(5):1204. https://doi.org/10.3390/v15051204
Chicago/Turabian StyleWang, Guolei, Zhiqiang Li, Mingfu Tian, Xianghua Cui, Jun’e Ma, Siyu Liu, Chenglin Ye, Li Yuan, Muhammad Suhaib Qudus, Uzair Afaq, and et al. 2023. "β-Glucan Induces Training Immunity to Promote Antiviral Activity by Activating TBK1" Viruses 15, no. 5: 1204. https://doi.org/10.3390/v15051204
APA StyleWang, G., Li, Z., Tian, M., Cui, X., Ma, J., Liu, S., Ye, C., Yuan, L., Qudus, M. S., Afaq, U., Wu, K., Liu, X., & Zhu, C. (2023). β-Glucan Induces Training Immunity to Promote Antiviral Activity by Activating TBK1. Viruses, 15(5), 1204. https://doi.org/10.3390/v15051204