

PRRSV Elimination in a Farrow-to-Finish Pig Herd Using Herd Closure and Rollover Approach

 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. PRRSV Viral RNA and Antibody Detection
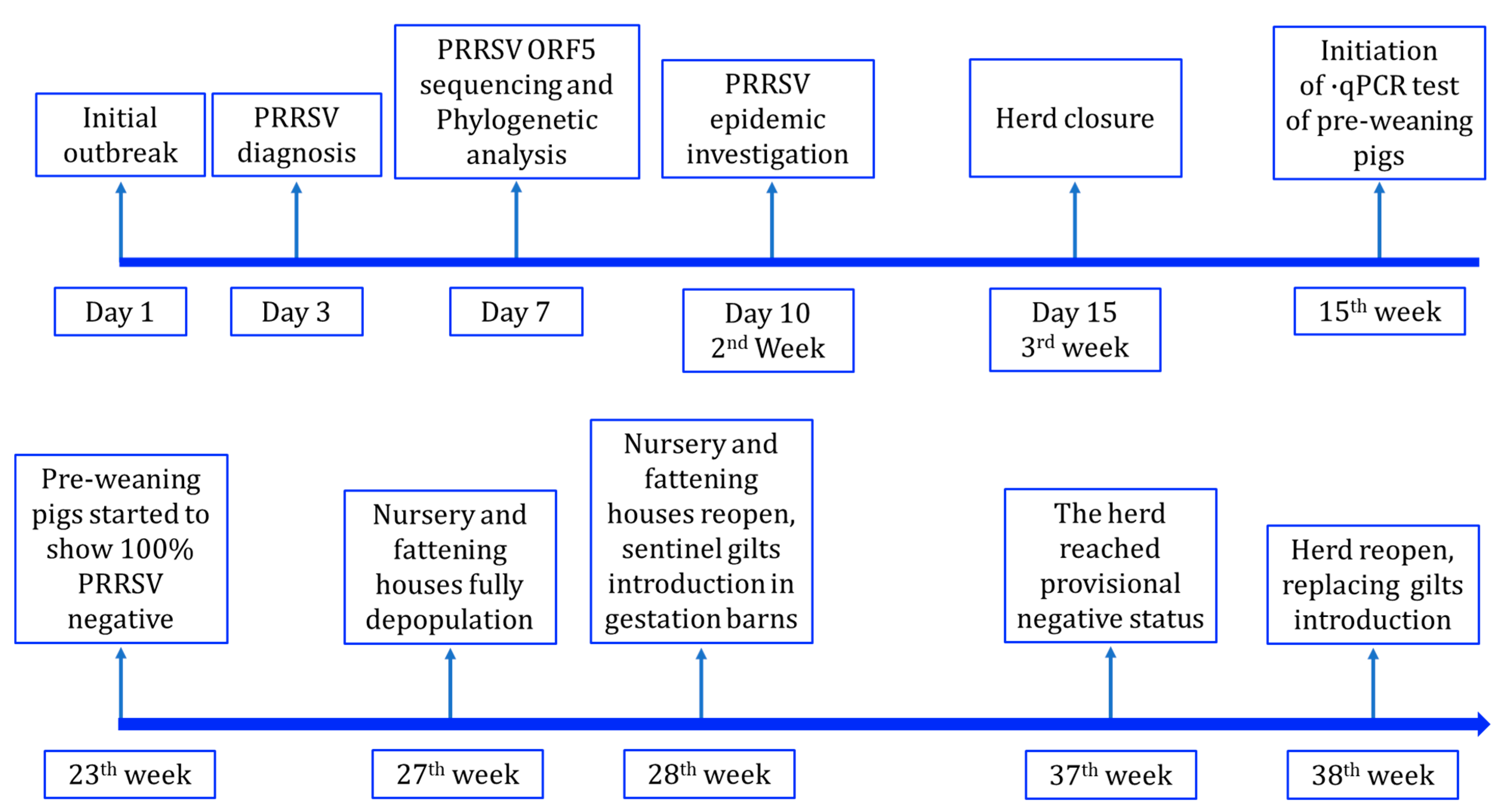
2.2. PRRSV Elimination Protocol
2.3. PRRSV Diagnosis and Surveillance Program
2.4. Production Data Collection
3. Results
3.1. The Herd Background
3.2. The Impact of PRRSV Outbreak
3.3. Diagnostic Results
3.4. The Time to Achieve Provisional Negative Status
3.5. The Time of Production Backing to Baseline
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, Y.; Zhang, Q.; Cao, Z.; Tang, Y.D.; Xia, D.; Wang, G.; Shan, H. Recent Advances in Porcine Reproductive and Respiratory Syndrome Virus NADC30-like Research in China: Molecular Characterization, Pathogenicity, and Control. Front. Microbiol. 2021, 12, 791313. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Corzo, C.A.; Mondaca, E.; Wayne, S.; Torremorell, M.; Dee, S.; Davies, P.; Morrison, R.B. Control and elimination of porcine reproductive and respiratory syndrome virus. Virus Res. 2010, 154, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Holtkamp, D.J.; Kliebenstein, J.B.; Neumann, E.; Zimmerman, J.J. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J. Swine Health Prod. 2013, 21, 72–84. [Google Scholar]
- Jiang, Y.; Li, G.; Yu, L.; Li, L.; Zhang, Y.; Zhou, Y.; Tong, W.; Liu, C.; Gao, F.; Tong, G. Genetic Diversity of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) From 1996 to 2017 in China. Front. Microbiol. 2020, 11, 618. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, L.; Xu, Z.; Deng, H.; Li, F.; Sun, X.; Zhou, Y.; Zhu, L. Emergence and spread of NADC34-like PRRSV in Southwest China. Transbound. Emerg. Dis. 2022, 69, e3416–e3424. [Google Scholar] [CrossRef]
- Li, X.; Bao, H.; Wang, Y.; Tian, K. Widespread of NADC30-like PRRSV in China: Another Pandora’s box for Chinese pig industry as the outbreak of highly pathogenic PRRSV in 2006? Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2017, 49, 12–13. [Google Scholar] [CrossRef]
- Holtkamp, D.J.; Polson, D.D.; Torremorell, M.; Morrison, B.; Classen, D.M.; Becton, L.; Henry, S.; Rodibaugh, M.T.; Rowland, R.R.; Snelson, H.; et al. Terminology for classifying swine herds by porcine reproductive and respiratory syndrome virus status. J. Swine Health Prod. 2011, 19, 44–56. [Google Scholar]
- Holtkamp, D.; Torremorell, M.; Corzo, C.A.; Linhares, D.C.L.; Almeida, M.N.; Yeske, P.; Polson, D.D.; Becton, L.; Snelson, H.; Donovan, T.; et al. Proposed modifications to porcine reproductive and respiratory syndrome virus herd classification. J. Swine Health Prod. 2021, 29, 261–270. [Google Scholar]
- Zhao, H.Z.; Wang, F.X.; Han, X.Y.; Guo, H.; Liu, C.Y.; Hou, L.N.; Wang, Y.X.; Zheng, H.; Wang, L.; Wen, Y.J. Recent advances in the study of NADC34-like porcine reproductive and respiratory syndrome virus in China. Front. Microbiol. 2022, 13, 950402. [Google Scholar] [CrossRef]
- Trevisan, G.; Sharma, A.; Gauger, P.; Harmon, K.M.; Zhang, J.; Main, R.; Zeller, M.; Linhares, L.C.M.; Linhares, D.C.L. PRRSV2 genetic diversity defined by RFLP patterns in the United States from 2007 to 2019. J. Vet. Diagn. Investig. 2021, 33, 920–931. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xu, Y.; Feng, W. Porcine Reproductive and Respiratory Syndrome Virus: Immune Escape and Application of Reverse Genetics in Attenuated Live Vaccine Development. Vaccines 2021, 9, 480. [Google Scholar] [CrossRef] [PubMed]
- Rupasinghe, R.; Lee, K.; Liu, X.; Gauger Phillip, C.; Zhang, J.; Martínez-López, B. Molecular Evolution of Porcine Reproductive and Respiratory Syndrome Virus Field Strains from Two Swine Production Systems in the Midwestern United States from 2001 to 2020. Microbiol. Spectr. 2022, 10, e02634-21. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.N.; Trible, B.R.; Chen, N.; Rowland, R.R.R. GP5 of porcine reproductive and respiratory syndrome virus (PRRSV) as a target for homologous and broadly neutralizing antibodies. Vet. Microbiol. 2017, 209, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Wang, Y.; Xu, X.; Sun, Z.; Xiao, Y.; Ji, G.; Li, Y.; Tan, F.; Li, X.; Tian, K. Commercial vaccines provide limited protection to NADC30-like PRRSV infection. Vaccine 2016, 34, 5540–5545. [Google Scholar] [CrossRef]
- Jiang, Y.-F.; Xia, T.-Q.; Zhou, Y.-J.; Yu, L.-X.; Yang, S.; Huang, Q.-F.; Li, L.-W.; Gao, F.; Qu, Z.-H.; Tong, W.; et al. Characterization of three porcine reproductive and respiratory syndrome virus isolates from a single swine farm bearing strong homology to a vaccine strain. Vet. Microbiol. 2015, 179, 242–249. [Google Scholar] [CrossRef]
- Zhou, L.; Ge, X.; Yang, H. Porcine Reproductive and Respiratory Syndrome Modified Live Virus Vaccine: A “Leaky” Vaccine with Debatable Efficacy and Safety. Vaccines 2021, 9, 362. [Google Scholar] [CrossRef]
- Scott Dee, J.C.; Halbur, P.; Keffaber, K.; Lautner, B.; McCaw, M.; Poison, D.; Rodibaugh, M.; Sanford, E.; Yeske, P. Control of porcine reproductive and respiratory syndrome PRRS virus. J. Swine Health Prod. 1996, 4, 95–98. [Google Scholar]
- Batista, L.; Pijoan, C.; Torremorell, M. Experimental injection of gilts with porcine reproductive and respiratory syndrome virus (PRRSV) during acclimatization. J. Swine Health Prod. 2002, 10, 147–150. [Google Scholar]
- Linhares, D.C.L. Evaluation of Immune Management Strategies to Control and Eliminate Porcine Reproductive and Respiratory Syndrome virus (PRRSv); University of Minnesota: Minneapolis, MN, USA, 2013. [Google Scholar]
- Szabo, I.; Molnar, T.; Nemes, I.; Abonyi, T.; Terjek, Z.; Balint, A. PRRSV eradication on large-scale fattening pig farms in Hungary between 2014 and 2019. Acta Vet. Hung. 2019, 67, 529–542. [Google Scholar] [CrossRef]
- Pertich, A.; Barna, Z.; Makai, O.; Farkas, J.; Molnar, T.; Balint, A.; Szabo, I.; Albert, M. Elimination of porcine reproductive and respiratory syndrome virus infection using an inactivated vaccine in combination with a roll-over method in a Hungarian large-scale pig herd. Acta Vet. Scand. 2022, 64, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Qu, X.; Wang, X.; Li, Z.; Yang, S.; Sun, L.; Zhou, B. Production Performance of Four Pig Herds Infected With Porcine Reproductive and Respiratory Syndrome Using the “Load-Close-Exposure” Approach in China. Front. Vet. Sci. 2022, 9, 882971. [Google Scholar] [CrossRef]
- Linhares, D.C.; Cano, J.P.; Torremorell, M.; Morrison, R.B. Comparison of time to PRRSv-stability and production losses between two exposure programs to control PRRSv in sow herds. Prev. Vet. Med. 2014, 116, 111–119. [Google Scholar] [CrossRef]
- Rathkjen, P.H.; Dall, J. Control and eradication of porcine reproductive and respiratory syndrome virus type 2 using a modified-live type 2 vaccine in combination with a load, close, homogenise model: An area elimination study. Acta Vet. Scand. 2017, 59, 4. [Google Scholar] [CrossRef] [PubMed]
- Szabo, I.; Bognar, L.; Molnar, T.; Nemes, I.; Balint, A. PRRS eradication from swine farms in five regions of Hungary. Acta Vet. Hung. 2020, 68, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Toman, M.; Celer, V.; Smola, J. Successful elimination of PRRS virus from an infected farrow-to-finish herd by vaccination. Veterinární Med. 2017, 62, 553–558. [Google Scholar] [CrossRef]
- Chen, P.; Tan, X.; Lao, M.; Wu, X.; Zhao, X.; Zhou, S.; Yu, J.; Zhu, J.; Yu, L.; Tong, W.; et al. The Novel PRRSV Strain HBap4-2018 with a Unique Recombinant Pattern Is Highly Pathogenic to Piglets. Virol. Sin. 2021, 36, 1611–1625. [Google Scholar] [CrossRef]
- Guo, J.; Liu, Z.; Tong, X.; Wang, Z.; Xu, S.; Chen, Q.; Zhou, J.; Fang, L.; Wang, D.; Xiao, S. Evolutionary Dynamics of Type 2 Porcine Reproductive and Respiratory Syndrome Virus by Whole-Genome Analysis. Viruses 2021, 13, 2469. [Google Scholar] [CrossRef]
- Opriessnig, T.; Halbur, P.G.; Yoon, K.J.; Pogranichniy, R.M.; Harmon, K.M.; Evans, R.; Key, K.F.; Pallares, F.J.; Thomas, P.; Meng, X.J. Comparison of molecular and biological characteristics of a modified live porcine reproductive and respiratory syndrome virus (PRRSV) vaccine (ingelvac PRRS MLV), the parent strain of the vaccine (ATCC VR2332), ATCC VR2385, and two recent field isolates of PRRSV. J. Virol. 2002, 76, 11837–11844. [Google Scholar]
- Calderon Diaz, J.A.; Fitzgerald, R.M.; Shalloo, L.; Rodrigues da Costa, M.; Niemi, J.; Leonard, F.C.; Kyriazakis, I.; Garcia Manzanilla, E. Financial Analysis of Herd Status and Vaccination Practices for Porcine Reproductive and Respiratory Syndrome Virus, Swine Influenza Virus, and Mycoplasma hyopneumoniae in Farrow-to-Finish Pig Farms Using a Bio-Economic Simulation Model. Front. Vet. Sci. 2020, 7, 556674. [Google Scholar] [CrossRef]
- Zhang, Z.; Qu, X.; Zhang, H.; Tang, X.; Bian, T.; Sun, Y.; Zhou, M.; Ren, F.; Wu, P. Evolutionary and recombination analysis of porcine reproductive and respiratory syndrome isolates in China. Virus Genes 2020, 56, 354–360. [Google Scholar] [CrossRef]
- Moura, C.A.A.; Philips, R.; Silva, G.S.; Holtkamp, D.J.; Linhares, D.C.L. Comparison of virus detection, productivity, and economic performance between lots of growing pigs vaccinated with two doses or one dose of PRRS MLV vaccine, under field conditions. Prev. Vet. Med. 2022, 204, 105669. [Google Scholar] [CrossRef]
- Linhares, D.C.L.; Betlach, C.; Morrison, R.B. Effect of immunologic solutions on sows and gilts on time to stability, and production losses in breeding herds infected with 1-7-4 PRRSv. Prev. Vet. Med. 2017, 144, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Linhares, D.C.; Cano, J.P.; Wetzell, T.; Nerem, J.; Torremorell, M.; Dee, S.A. Effect of modified-live porcine reproductive and respiratory syndrome virus (PRRSv) vaccine on the shedding of wild-type virus from an infected population of growing pigs. Vaccine 2012, 30, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Rowland, R.R.; Morrison, R.B. Challenges and opportunities for the control and elimination of porcine reproductive and respiratory syndrome virus. Transbound. Emerg. Dis. 2012, 59 (Suppl. 1), 55–59. [Google Scholar] [CrossRef] [PubMed]
- Rajic, A.; Dewey, C.; Deckert, A.; Friendship, R.; Martin, D.; Yoo, D. Production of PRRSV-negative pigs commingled from multiple, vaccinated, serologically stable, PRRSV-positive breeding herds. J. Swine Health Prod. 2001, 9, 179–184. [Google Scholar]
- Dong, J.G.; Yu, L.Y.; Wang, P.P.; Zhang, L.Y.; Liu, Y.L.; Liang, P.S.; Song, C.X. A new recombined porcine reproductive and respiratory syndrome virus virulent strain in China. J. Vet. Sci. 2018, 19, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Zhu, W.; Chen, Y.; Wei, C.; Zhou, P.; Zhang, M.; Huang, Z.; Sun, L.; Su, S.; Zhang, G. Molecular epidemiology of PRRSV in South China from 2007 to 2011 based on the genetic analysis of ORF5. Microb. Pathog. 2013, 63, 30–36. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sampling Time | qPCR Results | Positive Ratio | Sampling Time | qPCR Results | Positive Ratio | Sampling Time | qPCR Results | Positive Ratio |
|---|---|---|---|---|---|---|---|---|
| 15th week * | 3/52 | 5.77% | 25th week | 0/57 | 0.00% | 35th week | 0/48 | 0.00% |
| 16th week | 2/55 | 3.64% | 26th week | 0/52 | 0.00% | 36th week | 0/52 | 0.00% |
| 17th week | 3/49 | 6.12% | 27th week | 0/50 | 0.00% | 37th week | 0/51 | 0.00% |
| 18th week | 0/51 | 0.00% | 28th week | 0/55 | 0.00% | 38th week | 0/51 | 0.00% |
| 19th week | 2/53 | 3.77% | 29th week | 0/53 | 0.00% | 42nd week | 0/57 | 0.00% |
| 20th week | 1/55 | 1.82% | 30th week | 0/52 | 0.00% | 46th week | 0/56 | 0.00% |
| 21st week | 0/61 | 0.00% | 31st week | 0/52 | 0.00% | 50th week | 0/57 | 0.00% |
| 22nd week | 1/57 | 1.75% | 32nd week | 0/54 | 0.00% | 54th week | 0/49 | 0.00% |
| 23rd week | 0/50 | 0.00% | 33rd week | 0/63 | 0.00% | 58th week | 0/54 | 0.00% |
| 24th week | 0/51 | 0.00% | 34th week | 0/57 | 0.00% | 62nd week | 0/51 | 0.00% |
| Sample Collection Time | Sows and Gilts | Pre-Weaning Pigs | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S:P Value | Distribution of S:P Values | S:P Value | Distribution of S:P Values | |||||||||
| Mean | Range | <0.4 | 0.4–1.0 | 1.1–2.0 | >2.0 | Mean | Range | <0.4 | 0.4–1.0 | 1.1–2.0 | >2.0 | |
| 5th week * | 1.97 | 0.87–2.83 | 0/30 | 3/30 | 9/30 | 16/30 | 1.66 | 0.22–2.37 | 6/30 | 11/30 | 9/30 | 4/30 |
| 14th week | 1.713 | 0.43–1.8 | 1/30 | 9/30 | 24/30 | 0/30 | 0.67 | 0.24–1.84 | 16/50 | 21/50 | 8/50 | 5/50 |
| 26th week | 0.78 | 0.23–1.7 | 11/30 | 17/30 | 5/30 | 0/30 | 0.49 | 0.16–0.87 | 42/50 | 8/50 | 0/50 | 0/50 |
| 38th week | 0.35 | 0.28–1.1 | 20/30 | 7/30 | 3/30 | 0/30 | 0.26 | 0.08–0.52 | 48/50 | 2/50 | 0/50 | 0/50 |
| 50th week | 0.31 | 0.31–0.89 | 29/30 | 1/30 | 0/30 | 0/30 | 0.21 | 0.03–0.37 | 50/50 | 0/50 | 0/50 | 0/50 |
| 62nd week | 0.16 | 0.12–0.34 | 30/30 | 0/30 | 0/30 | 0/30 | 0.17 | 0.04–0.33 | 50/50 | 0/50 | 0/50 | 0/50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, R.; Zhang, T.; Lai, R.; Ding, Z.; Zhuang, Y.; Liu, H.; Cao, H.; Gao, X.; Luo, J.; Chen, Z.; et al. PRRSV Elimination in a Farrow-to-Finish Pig Herd Using Herd Closure and Rollover Approach. Viruses 2023, 15, 1239. https://doi.org/10.3390/v15061239
Hu R, Zhang T, Lai R, Ding Z, Zhuang Y, Liu H, Cao H, Gao X, Luo J, Chen Z, et al. PRRSV Elimination in a Farrow-to-Finish Pig Herd Using Herd Closure and Rollover Approach. Viruses. 2023; 15(6):1239. https://doi.org/10.3390/v15061239
Chicago/Turabian StyleHu, Ruiming, Tiansheng Zhang, Rongbin Lai, Zhen Ding, Yu Zhuang, Hao Liu, Huabin Cao, Xiaona Gao, Junrong Luo, Zheng Chen, and et al. 2023. "PRRSV Elimination in a Farrow-to-Finish Pig Herd Using Herd Closure and Rollover Approach" Viruses 15, no. 6: 1239. https://doi.org/10.3390/v15061239
APA StyleHu, R., Zhang, T., Lai, R., Ding, Z., Zhuang, Y., Liu, H., Cao, H., Gao, X., Luo, J., Chen, Z., Zhang, C., Liu, P., Guo, X., Hu, G., Ding, N., & Deng, S. (2023). PRRSV Elimination in a Farrow-to-Finish Pig Herd Using Herd Closure and Rollover Approach. Viruses, 15(6), 1239. https://doi.org/10.3390/v15061239