
Significant Broad-Spectrum Antiviral Activity of Bi121 against Different Variants of SARS-CoV-2
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Production and Standardization of Bi121
2.2. Enrichment of Polyphenolic Rich Bi121
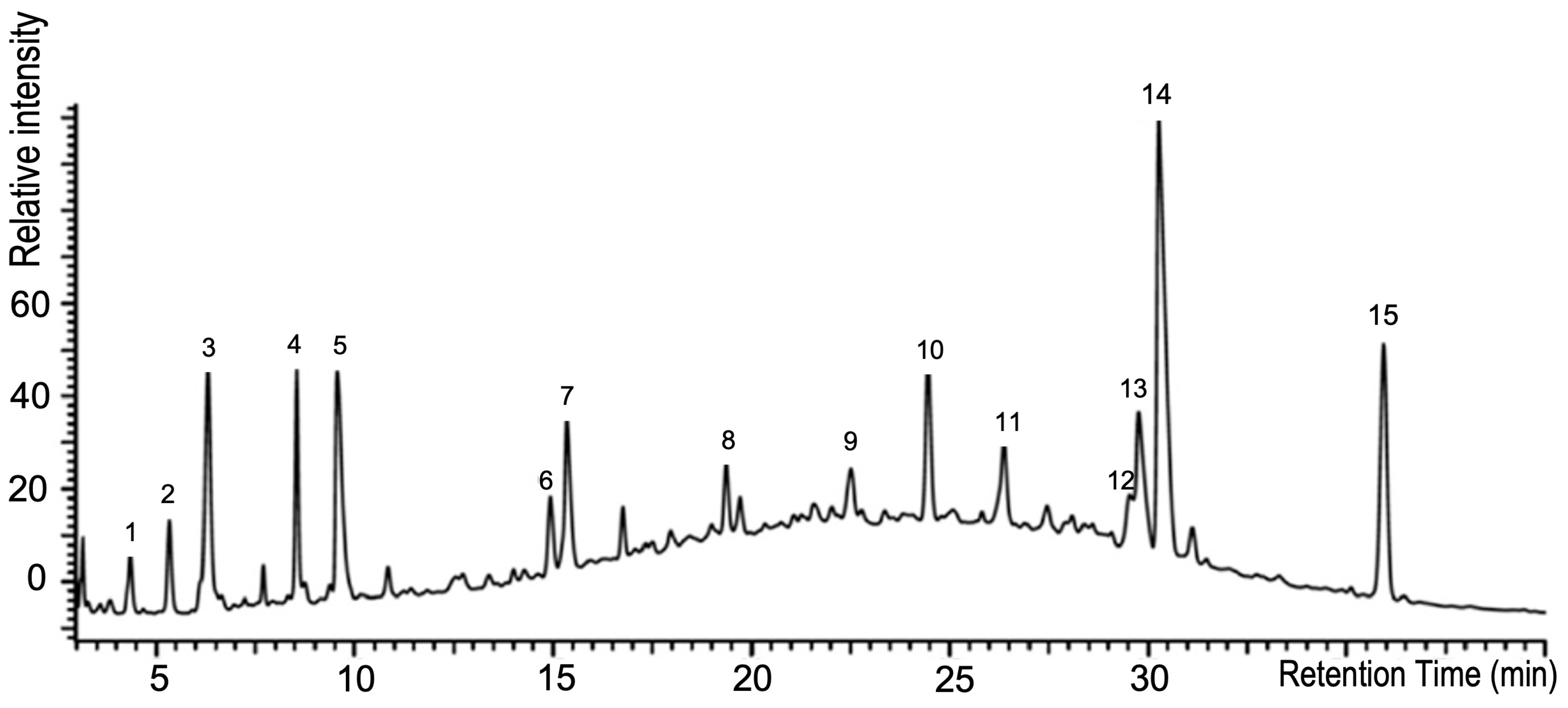
2.3. Fractionation of Bi121 Polyphenol Fractions Using Reversed-Phase UHPLC
2.4. Nanospray LC/MS/MS Analysis and Identification of Active Compounds
2.5. Cell Lines and Virus
2.6. rVSV-ΔG-SARS-CoV-2 S Luciferase Pseudotype Assay and Cytotoxicity Assay
2.7. Cytotoxicity Assay
2.8. Antiviral Effect of Bi121
2.9. Quantitative Real Time-PCR (qRT-PCR)
2.10. Plaque Assay
2.11. Tissue Culture Infectious Dose 50 (TCID50)
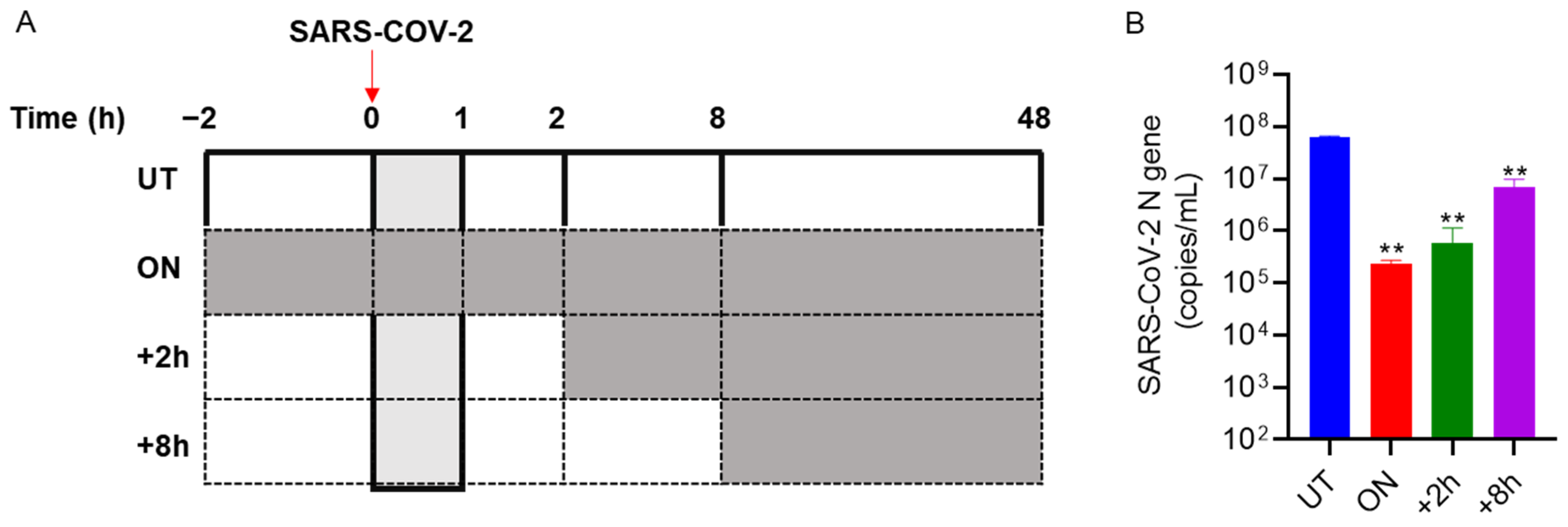
2.12. Time of Addition Assay
2.13. Target Prediction
2.14. Computational Validation
2.15. Statistical Analysis
3. Results
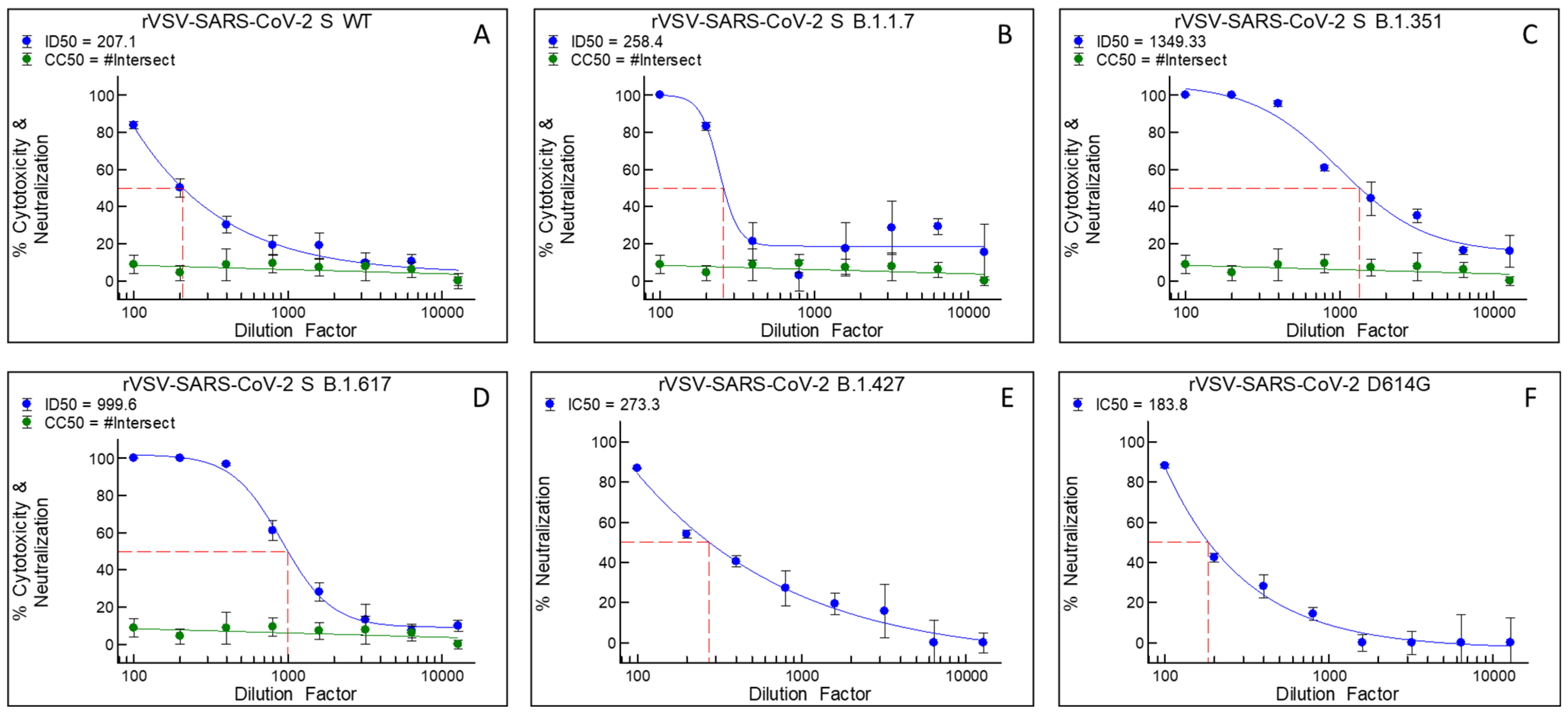
3.1. Bi121 Neutralized Pseudotyped-SARS-CoV-2 S (Spike Protein) Representing Various Viral Mutations
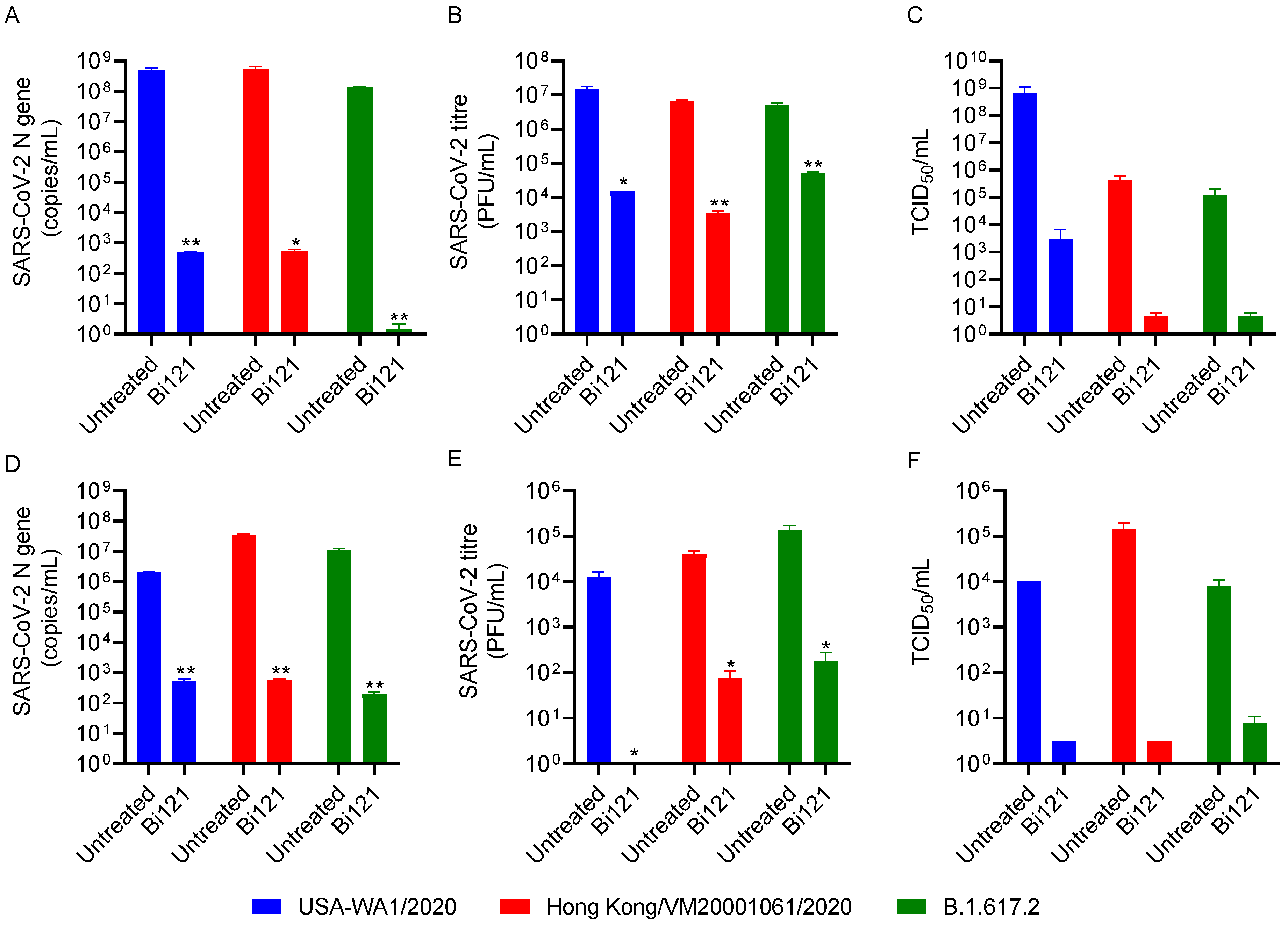
3.2. Bi121 Neutralized Three Variants of SARS-CoV-2 in Two Different Cell Lines
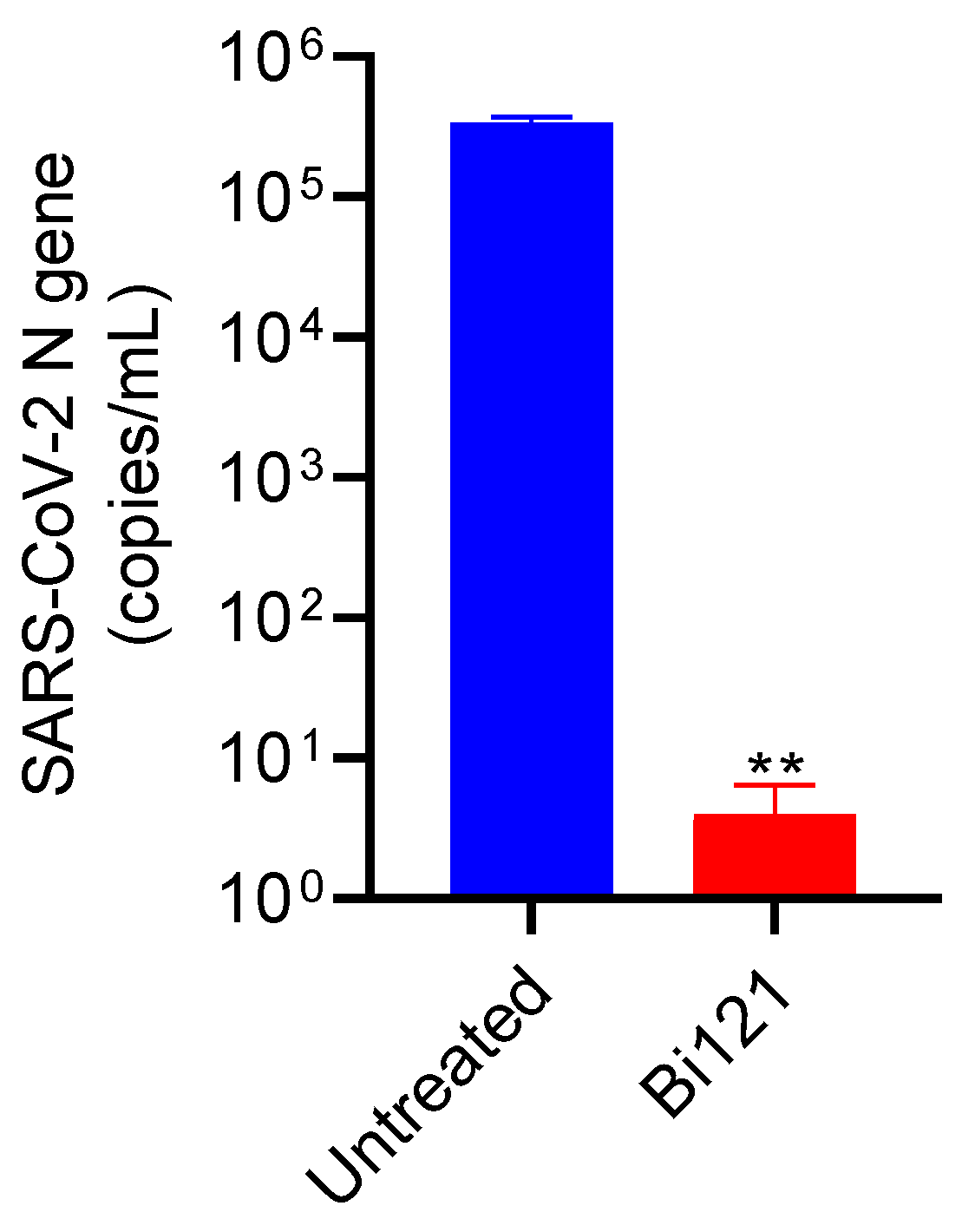
3.3. The Anti-SARS-CoV-2 Activity of Bi121 against Omicron Strain
3.4. Bi121 Interfered with the Early Stages of SARS-CoV-2 Infection
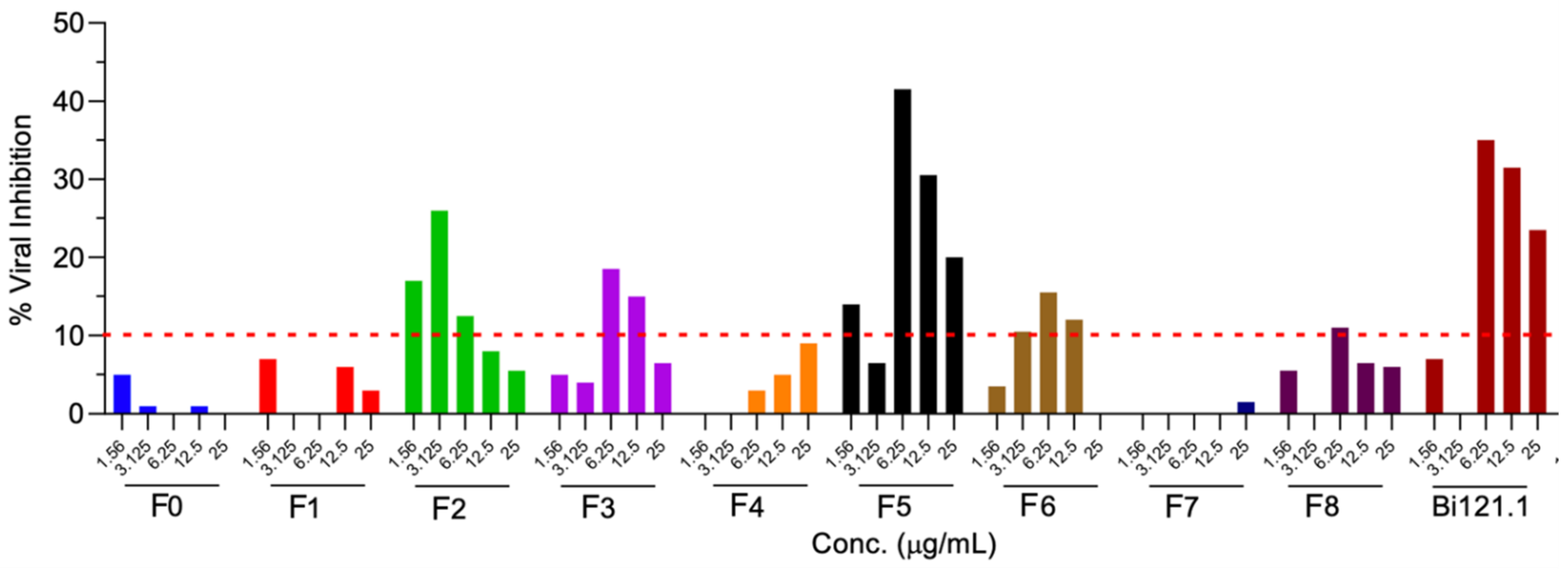
3.5. Antiviral Activity of Various Fractionated Polyphenolic Compounds from Bi121
3.6. Identification of Potential Polyphenol and Flavonoid Fractions from the Bi121 Extract Using LC/MS/MS
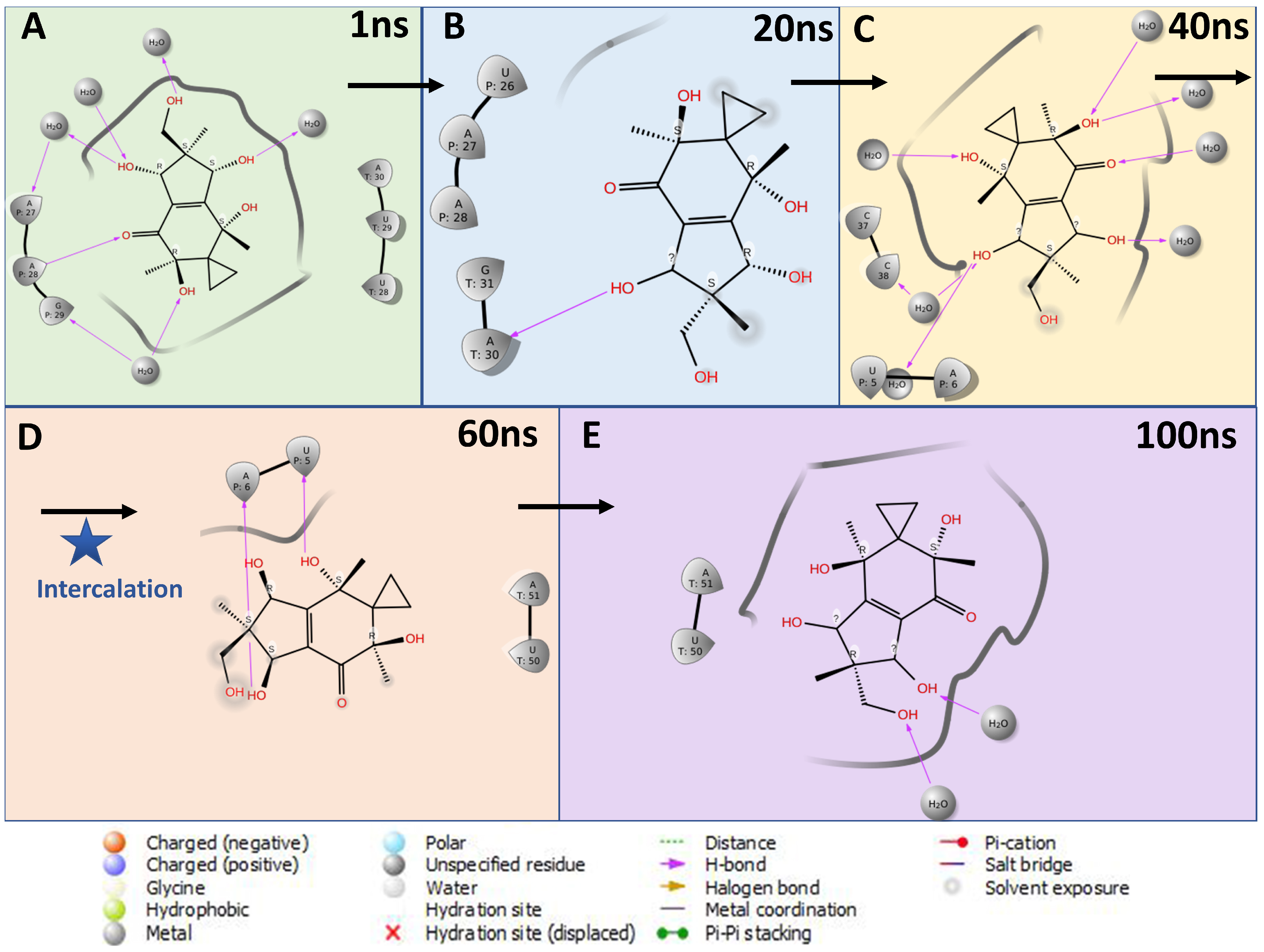
3.7. Target Prediction and Computational Target Validation
4. Discussion
5. Summary
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Declaration of Interest
References
- WHO. WHO COVID-19 Dashboard; World Health Organization: Geneva, Switzerland, 2023. Available online: https://covid19.who.int/ (accessed on 10 May 2023).
- Rochman, N.D.; Wolf, Y.I.; Faure, G.; Mutz, P.; Zhang, F.; Koonin, E.V. Ongoing global and regional adaptive evolution of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2021, 118, 336644. [Google Scholar] [CrossRef] [PubMed]
- Tabibzadeh, A.; Esghaei, M.; Soltani, S.; Yousefi, P.; Taherizadeh, M.; Safarnezhad Tameshkel, F.; Golahdooz, M.; Panahi, M.; Ajdarkosh, H.; Zamani, F.; et al. Evolutionary study of COVID-19, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) as an emerging coronavirus: Phylogenetic analysis and literature review. Vet. Med. Sci. 2021, 7, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Kannan, S.R.; Spratt, A.N.; Sharma, K.; Chand, H.S.; Byrareddy, S.N.; Singh, K. Omicron SARS-CoV-2 variant: Unique features and their impact on pre-existing antibodies. J. Autoimmun. 2022, 126, 102779. [Google Scholar] [CrossRef] [PubMed]
- Yamasoba, D.; Uriu, K.; Plianchaisuk, A.; Kosugi, Y.; Pan, L.; Zahradnik, J. Genotype to Phenotype Japan (G2P-Japan) Consortium; Ito, J.; Sato, K. Virological characteristics of the SARS-CoV-2 omicron XBB.1.16 variant. Lancet Infect. Dis, 2023; Online ahead of print. [Google Scholar]
- Afshar, Z.M.; Hosseinzadeh, D.; Hosseinzadeh, R.; Babazadeh, A.; Allahgholipour, A.; Sio, T.T.; Sullman, M.J.M.; Carson-Chahhoud, K.; Barary, M.; Ebrahimpour, S. The use of remdesivir in patients with COVID-19. Infect. Disord. Drug. Targets 2023. [Google Scholar] [CrossRef]
- Gottlieb, R.L.; Vaca, C.E.; Paredes, R.; Mera, J.; Webb, B.J.; Perez, G.; Oguchi, G.; Ryan, P.; Nielsen, B.U.; Brown, M.; et al. Early Remdesivir to Prevent Progression to Severe COVID-19 in Outpatients. N. Engl. J. Med. 2021, 386, 305–315. [Google Scholar] [CrossRef]
- Hammond, J.; Leister-Tebbe, H.; Gardner, A.; Abreu, P.; Bao, W.; Wisemandle, W.; Baniecki, M.; Hendrick, V.M.; Damle, B.; Simón-Campos, A.; et al. Oral Nirmatrelvir for High-Risk, Nonhospitalized Adults with COVID-19. N. Engl. J. Med. 2022, 386, 1397–1408. [Google Scholar] [CrossRef]
- Jayk Bernal, A.; Gomes Da Silva, M.M.; Musungaie, D.B.; Kovalchuk, E.; Gonzalez, A.; Delos Reyes, V.; Martín-Quirós, A.; Caraco, Y.; Williams-Diaz, A.; Brown, M.L.; et al. Molnupiravir for Oral Treatment of COVID-19 in Nonhospitalized Patients. N. Engl. J. Med. 2021, 386, 509–520. [Google Scholar] [CrossRef]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef]
- Anjani Kumar, S.; Rathi, B.; Poonam. Recent updates on the biological efficacy of approved drugs and potent synthetic compounds against SARS-CoV-2. RSC Adv. 2023, 13, 3677–3687. [Google Scholar] [CrossRef]
- Martinez, M.A. Efficacy of repurposed antiviral drugs: Lessons from COVID-19. Drug. Discov. Today 2022, 27, 1954–1960. [Google Scholar] [CrossRef]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell. Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, R. Structural Basis of the Potential Binding Mechanism of Remdesivir to SARS-CoV-2 RNA-Dependent RNA Polymerase. J. Phys. Chem. B. 2020, 124, 6955–6962. [Google Scholar] [CrossRef] [PubMed]
- Bafna, K.; Cioffi, C.L.; Krug, R.M.; Montelione, G.T. Structural similarities between SARS-CoV2 3CLpro and other viral proteases suggest potential lead molecules for developing broad spectrum antivirals. Front. Chem. 2022, 10, 948553. [Google Scholar] [CrossRef] [PubMed]
- Zhan, P.; Yu, B.; Ouyang, L. Drug repurposing: An effective strategy to accelerate contemporary drug discovery. Drug. Discov. Today 2022, 27, 1785–1788. [Google Scholar] [CrossRef] [PubMed]
- Brendler, T.; Van Wyk, B.E. A historical, scientific and commercial perspective on the medicinal use of Pelargonium sidoides (Geraniaceae). J. Ethnopharmacol. 2008, 119, 420–433. [Google Scholar] [CrossRef] [PubMed]
- Kolodziej, H. Fascinating metabolic pools of Pelargonium sidoides and Pelargonium reniforme, traditional and phytomedicinal sources of the herbal medicine Umckaloabo. Phytomedicine 2007, 14 (Suppl. S6), 9–17. [Google Scholar] [CrossRef]
- Careddu, D.; Pettenazzo, A. Pelargonium sidoides extract EPs 7630: A review of its clinical efficacy and safety for treating acute respiratory tract infections in children. Int. J. Gen. Med. 2018, 11, 91–98. [Google Scholar] [CrossRef]
- Gokce, S.; Dortkardesler, B.E.; Yurtseven, A.; Kurugol, Z. Effectiveness of Pelargonium sidoides in pediatric patients diagnosed with uncomplicated upper respiratory tract infection: A single-blind, randomized, placebo-controlled study. Eur. J. Pediatr. 2021, 180, 3019–3028. [Google Scholar] [CrossRef]
- Michaelis, M.; Doerr, H.W.; Cinatl, J., Jr. Investigation of the influence of EPs(R) 7630, a herbal drug preparation from Pelargonium sidoides, on replication of a broad panel of respiratory viruses. Phytomedicine 2011, 18, 384–386. [Google Scholar] [CrossRef]
- Theisen, L.L.; Muller, C.P. EPs(R) 7630 (Umckaloabo(R)), an extract from Pelargonium sidoides roots, exerts anti-influenza virus activity in vitro and in vivo. Antivir. Res. 2012, 94, 147–156. [Google Scholar] [CrossRef]
- Helfer, M.; Koppensteiner, H.; Schneider, M.; Rebensburg, S.; Forcisi, S.; Muller, C.; Schmitt-Kopplin, P.; Schindler, M.; Brack-Werner, R. The root extract of the medicinal plant Pelargonium sidoides is a potent HIV-1 attachment inhibitor. PLoS ONE 2014, 9, e87487. [Google Scholar] [CrossRef]
- Schnitzler, P.; Schneider, S.; Stintzing, F.C.; Carle, R.; Reichling, J. Efficacy of an aqueous Pelargonium sidoides extract against herpesvirus. Phytomedicine 2008, 15, 1108–1116. [Google Scholar] [CrossRef] [PubMed]
- Papies, J.; Emanuel, J.; Heinemann, N.; Kulic, Z.; Schroeder, S.; Tenner, B.; Lehner, M.D.; Seifert, G.; Muller, M.A. Antiviral and Immunomodulatory Effects of Pelargonium sidoides DC. Root Extract EPs(R) 7630 in SARS-CoV-2-Infected Human Lung Cells. Front. Pharm. 2021, 12, 757666. [Google Scholar] [CrossRef] [PubMed]
- Whitt, M.A. Generation of VSV pseudotypes using recombinant DeltaG-VSV for studies on virus entry, identification of entry inhibitors, and immune responses to vaccines. J. Virol. Methods 2010, 169, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Case, J.B.; Bailey, A.L.; Kim, A.S.; Chen, R.E.; Diamond, M.S. Growth, detection, quantification, and inactivation of SARS-CoV-2. Virology 2020, 548, 39–48. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty Per Cent Endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- NCBI. 2022. PubChem Compound Summary for CID 9994839, Neoilludin B. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Neoilludin-B (accessed on 20 July 2022).
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Wang, X.; Shen, Y.; Wang, S.; Li, S.; Zhang, W.; Liu, X.; Lai, L.; Pei, J.; Li, H. PharmMapper 2017 update: A web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Res. 2017, 45, W356–W360. [Google Scholar] [CrossRef]
- Wang, L.; Ma, C.; Wipf, P.; Liu, H.; Su, W.; Xie, X.Q. TargetHunter: An in silico target identification tool for predicting therapeutic potential of small organic molecules based on chemogenomic database. AAPS J. 2013, 15, 395–406. [Google Scholar] [CrossRef]
- Gong, J.; Cai, C.; Liu, X.; Ku, X.; Jiang, H.; Gao, D.; Li, H. ChemMapper: A versatile web server for exploring pharmacology and chemical structure association based on molecular 3D similarity method. Bioinformatics 2013, 29, 1827–1829. [Google Scholar] [CrossRef]
- Kinnings, S.L.; Jackson, R.M. ReverseScreen3D: A structure-based ligand matching method to identify protein targets. J. Chem. Inf. Model. 2011, 51, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Balvociute, M.; Huson, D.H. SILVA, RDP, Greengenes, NCBI and OTT—how do these taxonomies compare? BMC Genom. 2017, 18, 114. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.W.; Prlic, A.; Altunkaya, A.; Bi, C.; Bradley, A.R.; Christie, C.H.; Costanzo, L.D.; Duarte, J.M.; Dutta, S.; Feng, Z.; et al. The RCSB protein data bank: Integrative view of protein, gene and 3D structural information. Nucleic Acids Res. 2017, 45, D271–D281. [Google Scholar] [PubMed]
- Molecular Operating Environment. Molecular Operating Environment (MOE) 2019.01; Chemical Computing Group ULC: Montreal, QC, Canada, 2019; H3A 2R7. [Google Scholar]
- Vilar, S.; Cozza, G.; Moro, S. Medicinal chemistry and the molecular operating environment (MOE): Application of QSAR and molecular docking to drug discovery. Curr. Top. Med. Chem. 2008, 8, 1555–1572. [Google Scholar] [CrossRef]
- Schrödinger Release 2020. Desmond Molecular Dynamics System. Maestro-Desmond Interoperability Tools; Schrödinger: New York, NY, USA; D. E. Shaw Research: New York, NY, USA, 2020. [Google Scholar]
- Bowers, K.J.; Chow, D.E.; Xu, H.; Dror, R.O.; Eastwood, M.P.; Gregersen, B.A.; Klepeis, J.L.; Kolossvary, I.; Moraes, M.A.; Sacerdoti, F.D.; et al. Scalable Algorithms for Molecular Dynamics Simulations on Commodity Clusters SC’06. In Proceedings of the 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 11–17 November 2006; p. 43. [Google Scholar]
- Le, P.; Nodwell, M.B.; Eirich, J.; Sieber, S.A. A Chemical Proteomic Analysis of Illudin-Interacting Proteins. Chemistry 2019, 25, 12644–12651. [Google Scholar] [CrossRef]
- Lehmann, V.K.; Huang, A.; Ibanez-Calero, S.; Wilson, G.R.; Rinehart, K.L.; Illudin, S. The sole antiviral compound in mature fruiting bodies of Omphalotus illudens. J. Nat. Prod. 2003, 66, 1257–1258. [Google Scholar] [CrossRef]
- Yedavalli, V.S.; Zhang, N.; Cai, H.; Zhang, P.; Starost, M.F.; Hosmane, R.S.; Jeang, K.T. Ring expanded nucleoside analogues inhibit RNA helicase and intracellular human immunodeficiency virus type 1 replication. J. Med. Chem. 2008, 51, 5043–5051. [Google Scholar] [CrossRef]
- Kelner, M.J.; Mcmorris, T.C.; Estes, L.; Rutherford, M.; Montoya, M.; Goldstein, J.; Samson, K.; Starr, R.; Taetle, R. Characterization of illudin S sensitivity in DNA repair-deficient Chinese hamster cells. Unusually high sensitivity of ERCC2 and ERCC3 DNA helicase-deficient mutants in comparison to other chemotherapeutic agents. Biochem. Pharm. 1994, 48, 403–409. [Google Scholar] [CrossRef]
- Ozkan, R.T.N. Structure–activity relationship studies on aza analogues of illudins. Journal of Molecular Structure. J. Mol. Struct. 2001, 572, 15–24. [Google Scholar] [CrossRef]
- Zoldakova, M. Biochemical Studies of Targeted and Bimodal Analogues of the Natural Anticancer Compounds Combretastatin A-4 and Illudin M. Ph.D. Thesis, University of Bayreuth, Bayreuth, Germany, 2011. [Google Scholar]
- Swart, M.; Fonseca Guerra, C.; Bickelhaupt, F.M. Hydrogen bonds of RNA are stronger than those of DNA, but NMR monitors only presence of methyl substituent in uracil/thymine. J. Am. Chem. Soc. 2004, 126, 16718–16719. [Google Scholar] [CrossRef]
- Nakamura, M.; Fukunaga, Y.; Sasa, K.; Ohtoshi, Y.; Kanaori, K.; Hayashi, H.; Nakano, H.; Yamana, K. Pyrene is highly emissive when attached to the RNA duplex but not to the DNA duplex: The structural basis of this difference. Nucleic Acids Res. 2005, 33, 5887–5895. [Google Scholar] [CrossRef] [PubMed]
- Giovanetti, M.; Benedetti, F.; Campisi, G.; Ciccozzi, A.; Fabris, S.; Ceccarelli, G.; Tambone, V.; Caruso, A.; Angeletti, S.; Zella, D.; et al. Evolution patterns of SARS-CoV-2: Snapshot on its genome variants. Biochem. Biophys. Res. Commun. 2021, 538, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Matthys, H.; Eisebitt, R.; Seith, B.; Heger, M. Efficacy and safety of an extract of Pelargonium sidoides (EPs 7630) in adults with acute bronchitis. A randomised, double-blind, placebo-controlled trial. Phytomedicine 2003, 10 (Suppl. S4), 7–17. [Google Scholar] [CrossRef]
- Gescher, K.; Hensel, A.; Hafezi, W.; Derksen, A.; Kühn, J. Oligomeric proanthocyanidins from Rumex acetosa L. inhibit the attachment of herpes simplex virus type-1. Antivir. Res. 2011, 89, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Tsukuda, S.; Watashi, K.; Hojima, T.; Isogawa, M.; Iwamoto, M.; Omagari, K.; Suzuki, R.; Aizaki, H.; Kojima, S.; Sugiyama, M.; et al. A new class of hepatitis B and D virus entry inhibitors, proanthocyanidin and its analogs, that directly act on the viral large surface proteins. Hepatology 2017, 65, 1104–1116. [Google Scholar] [CrossRef] [PubMed]
- Kuramoto, M.; Tsukihara, T.; Ono, N. Neoilludins A and B, new bioactive components from Lampteromyces japonicus. Chem. Lett. 1999, 28, 1113–1114. [Google Scholar] [CrossRef]
- Kinder, F.R., Jr.; Bair, K.W. Total Synthesis of (.+-.)-Illudin M. J. Org. Chem. 1994, 59, 6965–6967. [Google Scholar] [CrossRef]
- Eneh, S.C.; Okonji, O.C.; Chiburoma, A.G.; Francisca Ogochukwu, O.; Tuwleh, L.; Gideon, I.; Okonji, E.F.; Bushabu, F.N.; Mgbere, O. Marburg virus disease amid COVID-19 in West Africa: An emerging and re-emerging zoonotic epidemic threat, future implications and way forward. Adv. Infect. Dis. 2023, 10, 20499361231168520. [Google Scholar] [CrossRef]
- Patel, P.R.; Shah, S.U. Ebola Virus; StatPearls: Treasure Island, FL, USA, 2023. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| rVSV-ΔG-SARS-CoV-2 S | Specific Mutations in the Spike Protein of Pseudotyped Strains |
|---|---|
| Wuhan wild type | - |
| B.1.1.7 (Alpha) | A570D, D614G, D1118H, delH69-V70, N501Y, P681H, S982A, T716I, delY144 |
| B.1.351 (Beta) | K417N, E484K, N501Y, D614G, A701V |
| B.1.617 (Delta) | L452R, E484Q |
| D614G | D614G |
| B.1.427 | L452R, D614G |
| Fraction Identity | Putative Active Molecule | |
|---|---|---|
| SARS-CoV-2 Antiviral Activity (10–20% Viral Inhibition) | Presence/Absence of 298.33 Da (Bi121.2) Small Molecule Using MS | |
| Bi121 Fraction F0 | No | No |
| Bi121 Fraction F1 | No | No |
| Bi121 Fraction F2 | Yes | Yes |
| Bi121 Fraction F3 | Yes | Yes |
| Bi121 Fraction F4 | No | No |
| Bi121 Fraction F5 | Yes | Yes |
| Bi121 Fraction F6 | No | No |
| Bi121 Fraction F7 | No | No |
| Bi121 Fraction F8 | No | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subhadra, B.; Agrawal, R.; Pal, V.K.; Chenine, A.-L.; Mattathil, J.G.; Singh, A. Significant Broad-Spectrum Antiviral Activity of Bi121 against Different Variants of SARS-CoV-2. Viruses 2023, 15, 1299. https://doi.org/10.3390/v15061299
Subhadra B, Agrawal R, Pal VK, Chenine A-L, Mattathil JG, Singh A. Significant Broad-Spectrum Antiviral Activity of Bi121 against Different Variants of SARS-CoV-2. Viruses. 2023; 15(6):1299. https://doi.org/10.3390/v15061299
Chicago/Turabian StyleSubhadra, Bobban, Ragini Agrawal, Virender Kumar Pal, Agnes-Laurence Chenine, Jeffy George Mattathil, and Amit Singh. 2023. "Significant Broad-Spectrum Antiviral Activity of Bi121 against Different Variants of SARS-CoV-2" Viruses 15, no. 6: 1299. https://doi.org/10.3390/v15061299
APA StyleSubhadra, B., Agrawal, R., Pal, V. K., Chenine, A. -L., Mattathil, J. G., & Singh, A. (2023). Significant Broad-Spectrum Antiviral Activity of Bi121 against Different Variants of SARS-CoV-2. Viruses, 15(6), 1299. https://doi.org/10.3390/v15061299