

Modulation of Reoviral Cytolysis (I): Combination Therapeutics

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Chemotherapeutic Agents
2.3. Cell Viability Assay
2.4. Ras Activation Assay
2.5. FACS Analysis
2.6. Subcutaneous Tumor Xenograft Model in Nude Mice
2.7. Immunodetection of Reoviral Replication
3. Results
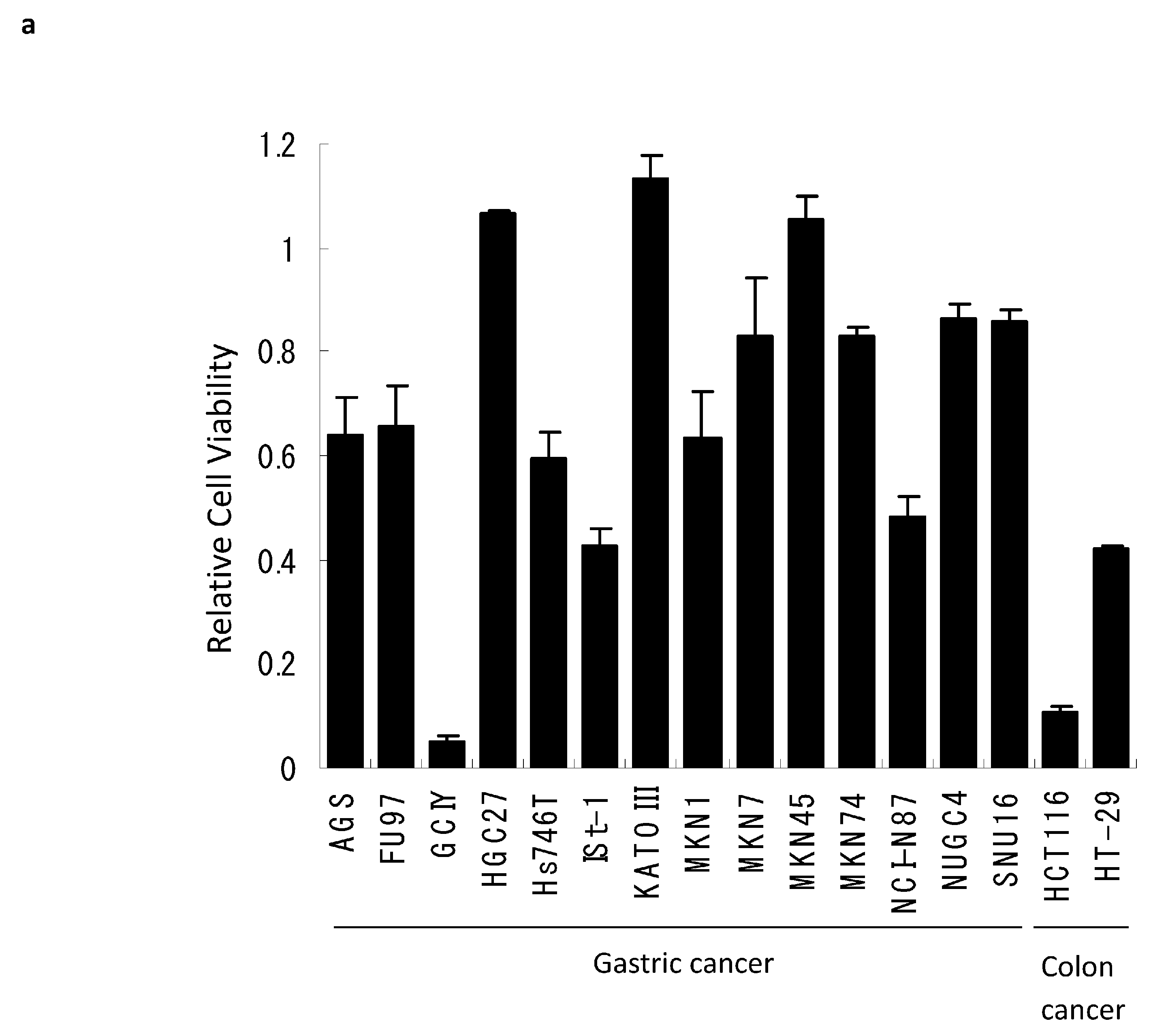
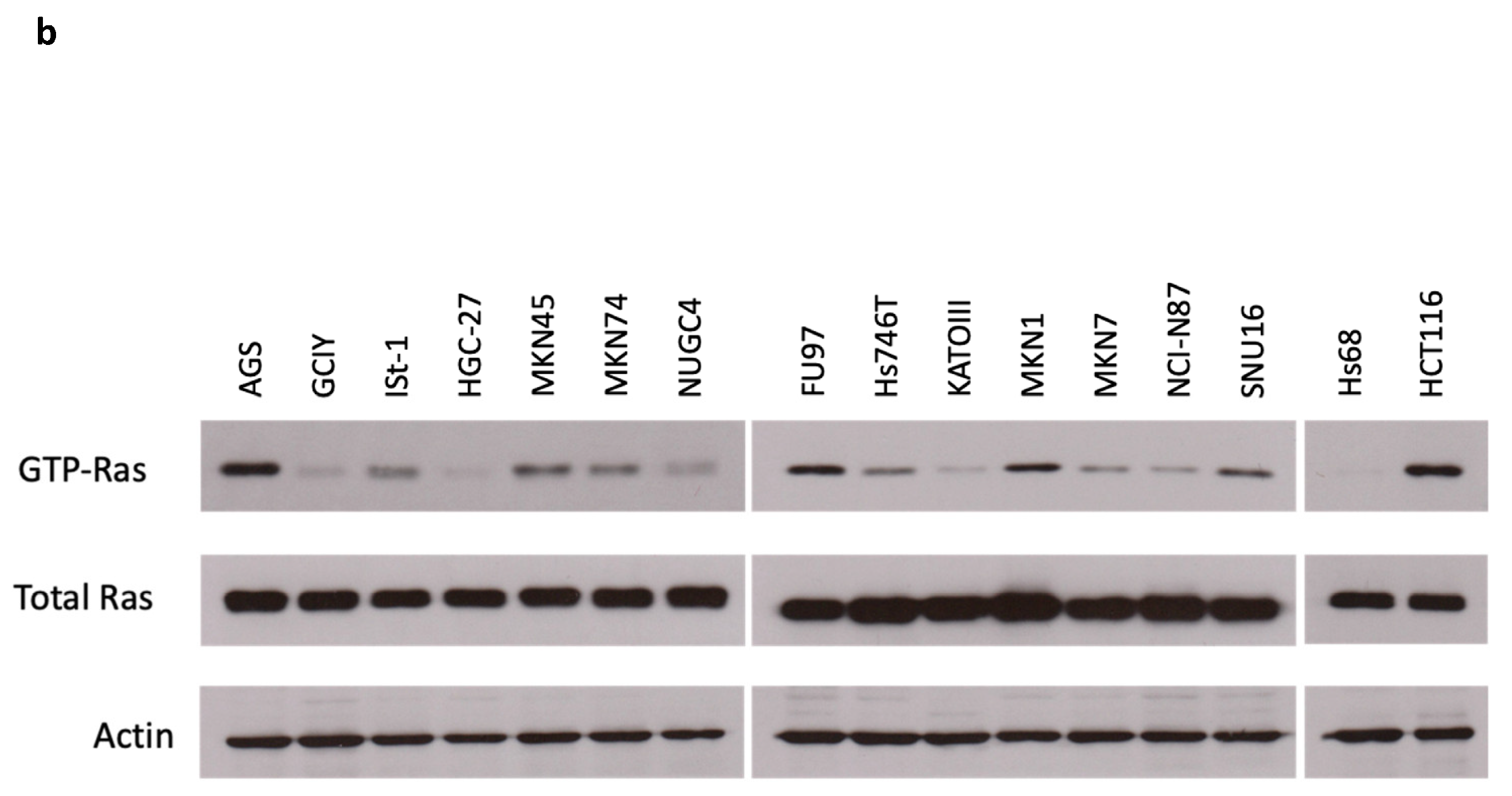
3.1. Reovirus Cytotoxicity in Gastric Cancer Cell Lines
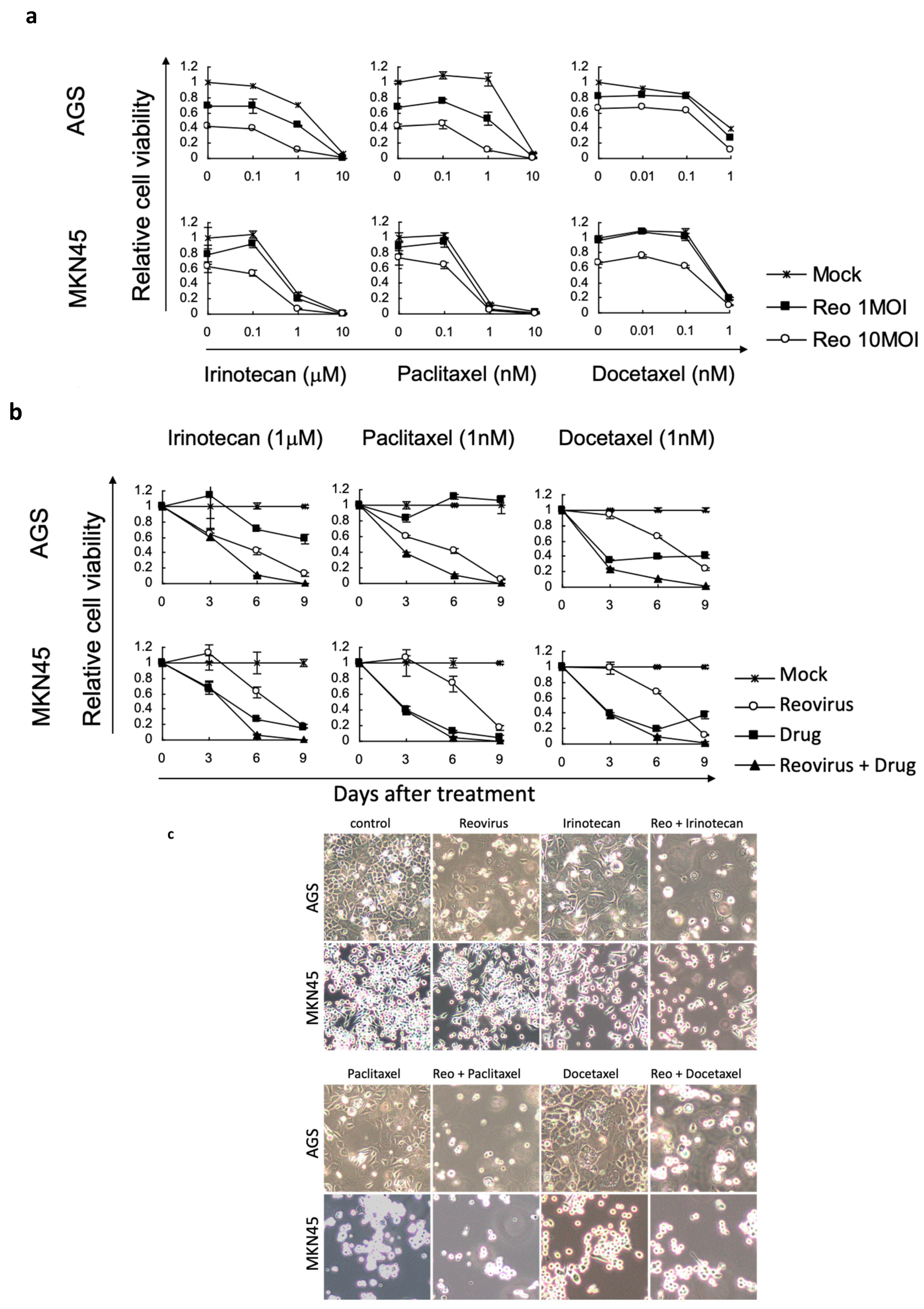
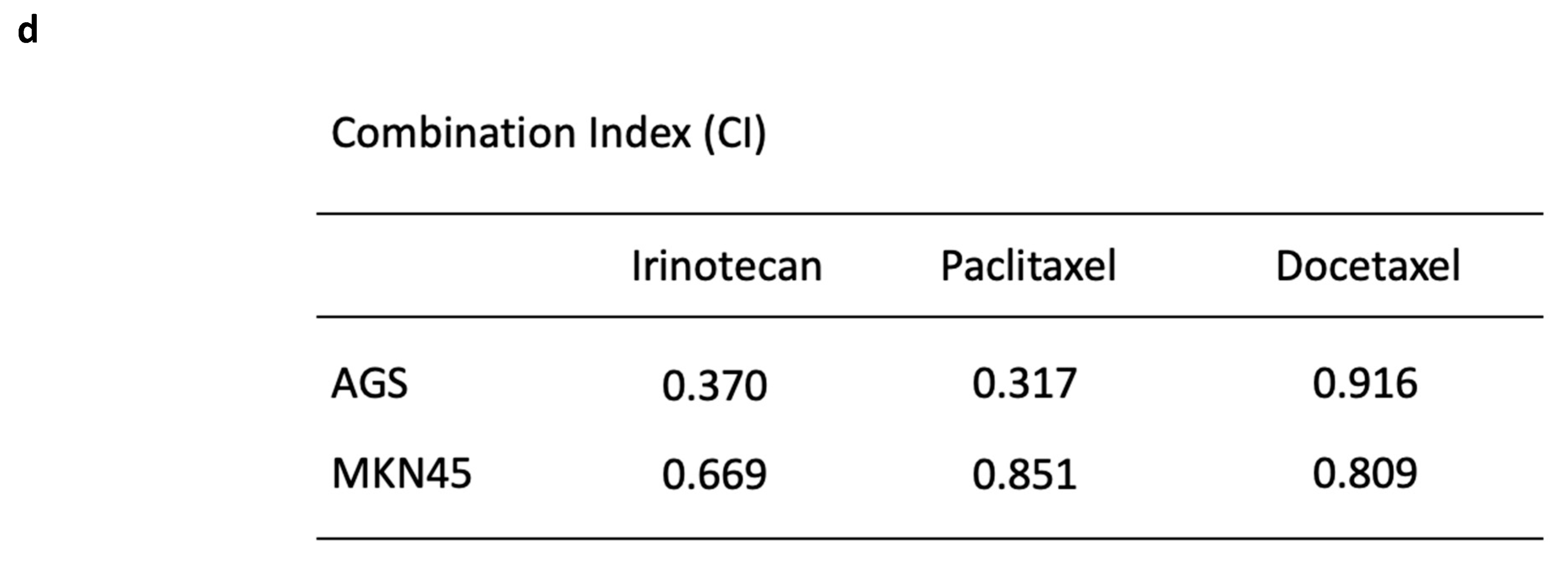
3.2. Reovirus Cytotoxicity with Chemotherapeutic Agents in Gastric Cancer Cell Lines
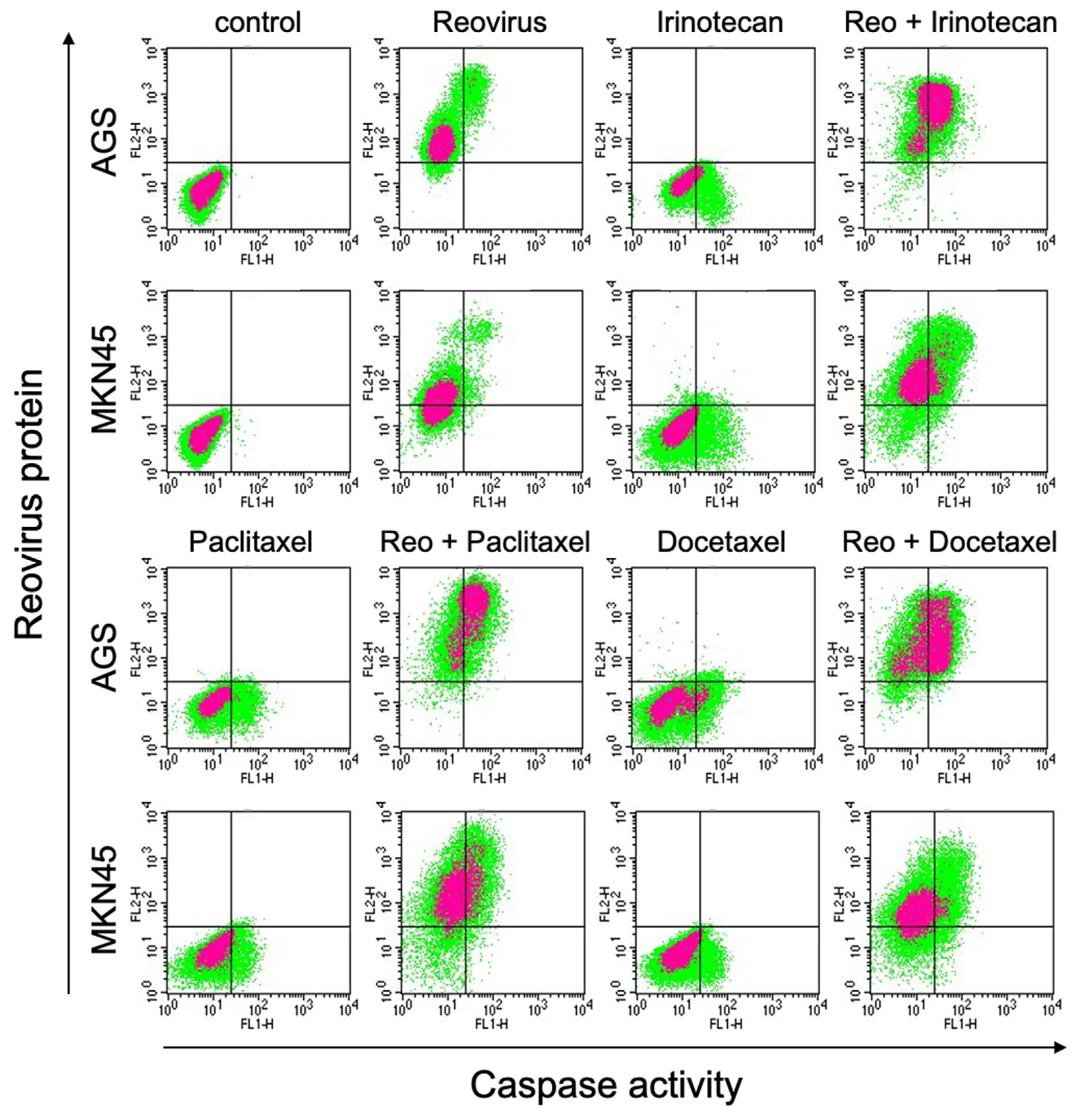
3.3. Combinations of Reovirus and Chemotherapeutic Agents Enhanced Reovirus Replication and Apoptosis
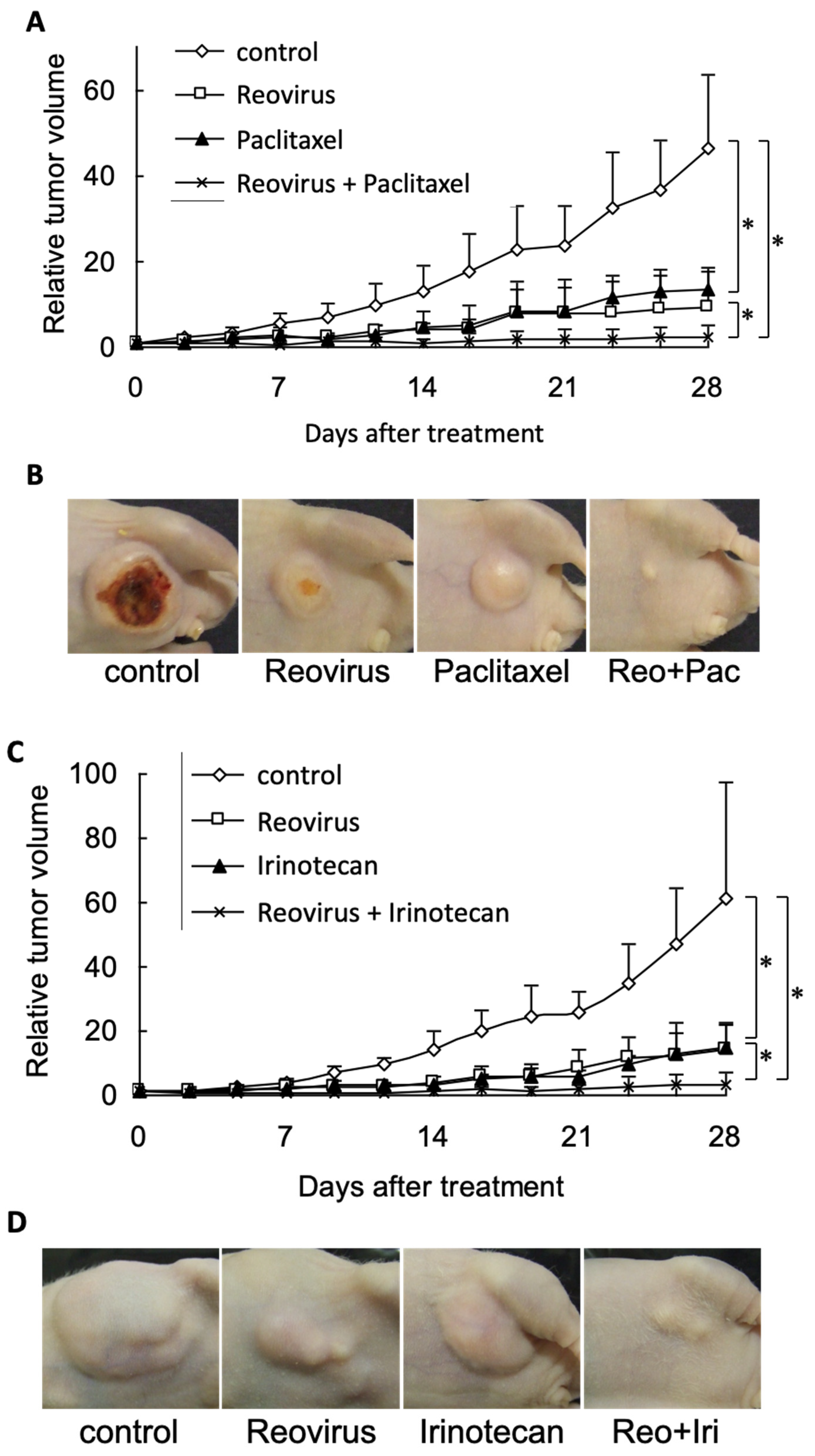
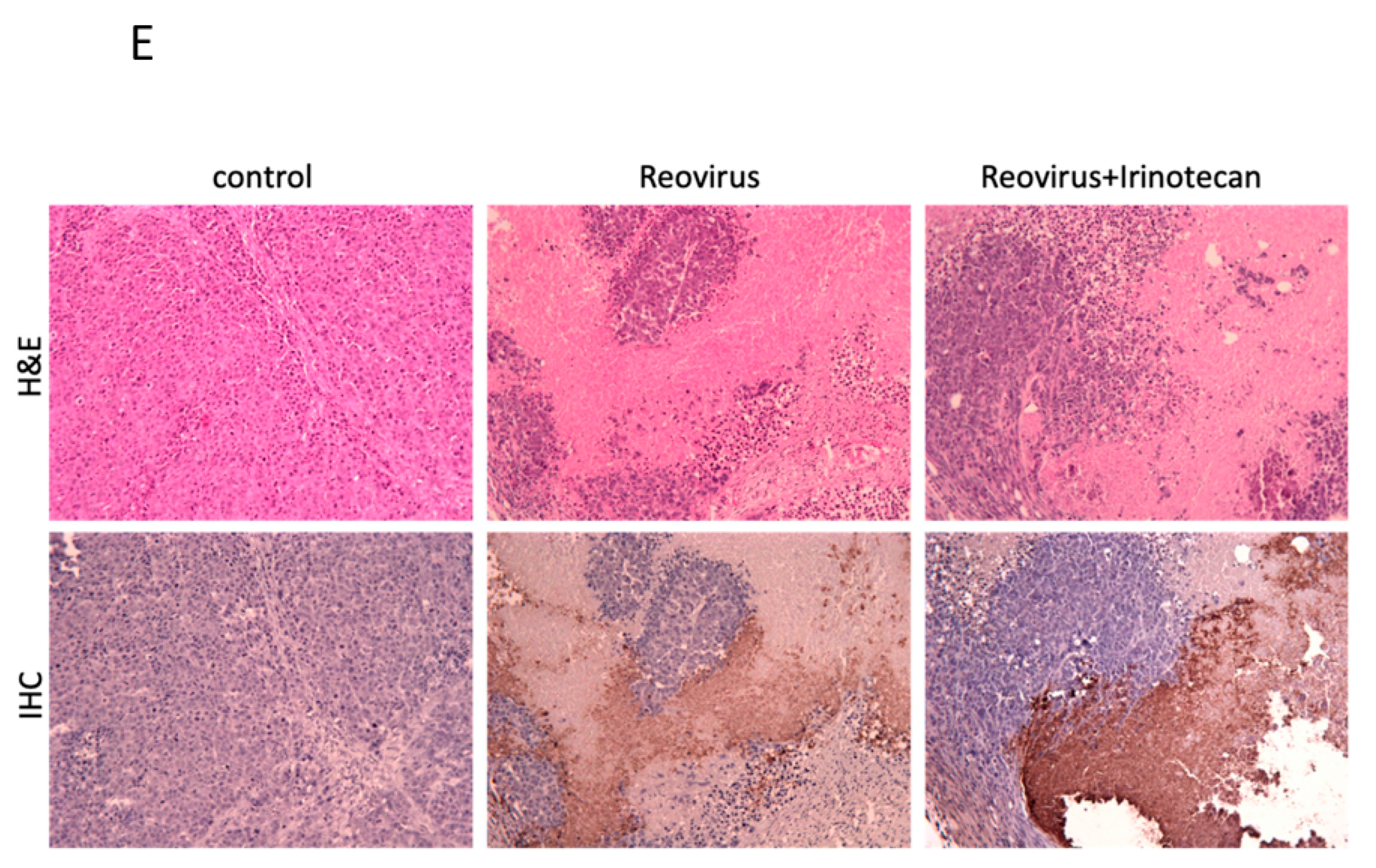
3.4. Combined Reovirus and Chemotherapeutic Agents Enhanced Anti-Tumor Effects in a Murine Gastric Cancer Xenograft Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Winer, E.; Gralow, J.; Diller, L.; Karlan, B.; Loehrer, P.; Pierce, L.; Demetri, G.; Ganz, P.; Kramer, B.; Kris, M.; et al. Clinical cancer advances 2008: Major research advances in cancer treatment, prevention, and screening—A report from the American Society of Clinical Oncology. J. Clin. Oncol. 2009, 27, 812–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozzetti, F.; Marubini, E.; Bonfanti, G.; Miceli, R.; Piano, C.; Gennari, L.; Italian Gastrointestinal Tumor Study Group. Subtotal versus total gastrectomy for gastric cancer: Five-year survival rates in a multicenter randomized Italian trial. Ann. Surg. 1999, 230, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Karpeh, M.S.; Leon, L.; Klimstra, D.; Brennan, M.F. Lymph node staging in gastric cancer: Is location more important than Number? An analysis of 1,038 patients. Ann. Surg. 2000, 232, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Wesolowski, R.; Lee, C.; Kim, R. Is there a role for second-line chemotherapy in advanced gastric cancer? Lancet Oncol. 2009, 10, 903–912. [Google Scholar] [CrossRef]
- Strong, J.E.; Coffey, M.C.; Tang, D.; Sabinin, P.; Lee, P.W. The molecular basis of viral oncolysis: Usurpation of the Ras signaling pathway by reovirus. EMBO J. 1998, 17, 3351–3362. [Google Scholar] [CrossRef] [Green Version]
- Duncan, M.R.; Stanish, S.M.; Cox, D.C. Differential sensitivity of normal and transformed human cells to reovirus infection. J. Virol. 1978, 28, 444–449. [Google Scholar] [CrossRef] [Green Version]
- Hashiro, G.; Loh, P.C.; Yau, J.T. The preferential cytotoxicity of reovirus for certain transformed cell lines. Arch. Virol. 1977, 54, 307–315. [Google Scholar] [CrossRef]
- Coffey, M.C.; Strong, J.E.; Forsyth, P.A.; Lee, P.W. Reovirus therapy of tumors with activated Ras pathway. Science 1998, 282, 1332–1334. [Google Scholar] [CrossRef]
- Hirasawa, K.; Nishikawa, S.G.; Norman, K.L.; Alain, T.; Kossakowska, A.; Lee, P.W. Oncolytic reovirus against ovarian and colon cancer. Cancer Res. 2002, 62, 1696–1701. [Google Scholar]
- Etoh, T.; Himeno, Y.; Matsumoto, T.; Aramaki, M.; Kawano, K.; Nishizono, A.; Kitano, S. Oncolytic viral therapy for human pancreatic cancer cells by reovirus. Clin. Cancer Res. 2003, 9, 1218–1223. [Google Scholar]
- Kim, M.; Egan, C.; Alain, T.; Urbanski, S.J.; Lee, P.W.; Forsyth, P.A.; Johnston, R.N. Acquired resistance to reoviral oncolysis in Ras-transformed fibrosarcoma cells. Oncogene 2007, 26, 4124–4134. [Google Scholar] [CrossRef]
- Oncolytics Biotech® Inc. Announces Receipt of Orphan Drug Designation from the U.S. FDA for Gastric Cancer. 2015. Available online: https://www.oncolyticsbiotech.com/press-releases (accessed on 5 June 2023).
- Toyoizumi, T.; Mick, R.; Abbas, A.E.; Kang, E.H.; Kaiser, L.R.; Molnar-Kimber, K.L. Combined therapy with chemotherapeutic agents and herpes simplex virus type 1 ICP34.5 mutant (HSV-1716) in human non-small cell lung cancer. Hum. Gene Ther. 1999, 10, 3013–3029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawa, A.; Nozawa, N.; Goshima, F.; Nagasaka, T.; Kikkawa, F.; Niwa, Y.; Nakanishi, T.; Kuzuya, K.; Nishiyama, Y. Oncolytic viral therapy for human ovarian cancer using a novel replication-competent herpes simplex virus type I mutant in a mouse model. Gynecol. Oncol. 2003, 91, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Cinatl, J., Jr.; Cinatl, J.; Michaelis, M.; Kabickova, H.; Kotchetkov, R.; Vogel, J.U.; Doerr, H.W.; Klingebiel, T.; Driever, P.H. Potent oncolytic activity of multimutated herpes simplex virus G207 in combination with vincristine against human rhabdomyosarcoma. Cancer Res. 2003, 63, 1508–1514. [Google Scholar]
- Petrowsky, H.; Roberts, G.D.; Kooby, D.A.; Burt, B.M.; Bennett, J.J.; Delman, K.A.; Stanziale, S.F.; Delohery, T.M.; Tong, W.P.; Federoff, H.J.; et al. Functional interaction between fluorodeoxyuridine-induced cellular alterations and replication of a ribonucleotide reductase-negative herpes simplex virus. J. Virol. 2001, 75, 7050–7058. [Google Scholar] [CrossRef] [Green Version]
- Bennett, J.J.; Adusumilli, P.; Petrowsky, H.; Burt, B.M.; Roberts, G.; Delman, K.A.; Zager, J.S.; Chou, T.C.; Fong, Y. Up-regulation of GADD34 mediates the synergistic anticancer activity of mitomycin C and a γ134.5 deleted oncolytic herpes virus (G207). FASEB J. 2004, 18, 1001–1003. [Google Scholar] [CrossRef] [PubMed]
- Aghi, M.; Rabkin, S.; Martuza, R.L. Effect of chemotherapy-induced DNA repair on oncolytic herpes simplex viral replication. J. Natl. Cancer Inst. 2006, 98, 38–50. [Google Scholar] [CrossRef]
- Sei, S.; Mussio, J.K.; Yang, Q.E.; Nagashima, K.; Parchment, R.E.; Coffey, M.C.; Shoemaker, R.H.; Tomaszewski, J.E. Synergistic antitumor activity of oncolytic reovirus and chemotherapeutic agents in non-small cell lung cancer cells. Mol. Cancer 2009, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Pandha, H.S.; Heinemann, L.; Simpson, G.R.; Melcher, A.; Prestwich, R.; Errington, F.; Coffey, M.; Harrington, K.J.; Morgan, R. Synergistic effects of oncolytic reovirus and cisplatin chemotherapy in murine malignant melanoma. Clin. Cancer Res. 2009, 15, 6158–6166. [Google Scholar] [CrossRef] [Green Version]
- Cho, I.R.; Koh, S.S.; Min, H.J.; Park, E.H.; Srisuttee, R.; Jhun, B.H.; Kang, C.D.; Kim, M.; Johnston, R.N.; Chung, Y.H. Reovirus infection induces apoptosis of TRAIL-resistant gastric cancer cells by down-regulation of Akt activation. Int. J. Oncol. 2010, 36, 1023–1030. [Google Scholar]
- Zhao, L.; Wientjes, M.G.; Au, J.L. Evaluation of combination chemotherapy: Integration of nonlinear regression, curve shift, isobologram, and combination index analyses. Clin. Cancer Res. 2004, 10, 7994–8004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, H.A.; Davydova, J.G.; Adachi, Y.; Takayama, K.; Barker, S.D.; Reynolds, P.N.; Krasnykh, V.N.; Kunisaki, C.; Shimada, H.; Curiel, D.T.; et al. Promoter-controlled infectivity-enhanced conditionally replicative adenoviral vectors for the treatment of gastric cancer. J. Gastroenterol. 2005, 40, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Mita, H.; Toyota, M.; Aoki, F.; Akashi, H.; Maruyama, R.; Sasaki, Y.; Suzuki, H.; Idogawa, M.; Kashima, L.; Yanagihara, K.; et al. A novel method, digital genome scanning detects KRAS gene amplification in gastric cancers: Involvement of overexpressed wild-type KRAS in downstream signaling and cancer cell growth. BMC Cancer 2009, 9, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Houdt, W.J.; Smakman, N.; van Den Wollenberg, D.J.M.; Emmink, B.L.; Veenendaal, L.M.; Van Diest, P.J.; Hoeben, R.C.; Borel Rinkes, I.H.M.; Kranenburg, O. Transient infection of freshly isolated human colorectal tumor cells by reovirus T3D intermediate subviral particles. Cancer Gene Ther. 2008, 15, 284–292. [Google Scholar] [CrossRef] [Green Version]
- Norman, K.L.; Hirasawa, K.; Yang, A.D.; Shields, M.A.; Lee, P.W. Reovirus oncolysis: The Ras/RalGEF/p38 pathway dictates host cell permissiveness to reovirus infection. Proc. Natl. Acad. Sci. USA 2004, 101, 11099–11104. [Google Scholar] [CrossRef] [Green Version]
- Barton, E.S.; Forrest, J.C.; Connolly, J.L.; Chappell, J.D.; Liu, Y.; Schnell, F.J.; Nusrat, A.; Parkos, C.A.; Dermody, T.S. Junction adhesion molecule is a receptor for reovirus. Cell 2001, 104, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Strong, J.E.; Lee, P.W. The v-erbB oncogene confers enhanced cellular susceptibility to reovirus infection. J. Virol. 1996, 70, 612–616. [Google Scholar] [CrossRef] [Green Version]
- Chappell, J.D.; Gunn, V.L.; Wetzel, J.D.; Baer, G.S.; Dermody, T.S. Mutations in type 3 reovirus that determine binding to sialic acid are contained in the fibrous tail domain of viral attachment protein sigma1. J. Virol. 1997, 71, 1834–1841. [Google Scholar] [CrossRef] [Green Version]
- Golden, J.W.; Linke, J.; Schmechel, S.; Thoemke, K.; Schiff, L.A. Addition of exogenous protease facilitates reovirus infection in many restrictive cells. J. Virol. 2002, 76, 7430–7443. [Google Scholar] [CrossRef] [Green Version]
- Alain, T.; Kim, T.S.; Lun, X.; Liacini, A.; Schiff, L.A.; Senger, D.L.; Forsyth, P.A. Proteolytic disassembly is a critical determinant for reovirus oncolysis. Mol. Ther. 2007, 15, 1512–1521. [Google Scholar] [CrossRef]
- Bischoff, J.R.; Kirn, D.H.; Williams, A.; Heise, C.; Horn, S.; Muna, M.; Ng, L.; Nye, J.A.; Sampson-Johannes, A.; Fattaey, A.; et al. An adenovirus mutant that replicates selectively in p53-deficient human tumor cells. Science 1996, 274, 373–376. [Google Scholar] [CrossRef]
- Rodriguez, R.; Schuur, E.R.; Lim, H.Y.; Henderson, G.A.; Simons, J.W.; Henderson, D.R. Prostate attenuated replication competent adenovirus (ARCA) CN706: A selective cytotoxic for prostate-specific antigen-positive prostate cancer cells. Cancer Res. 1997, 57, 2559–2563. [Google Scholar] [PubMed]
- Tsukuda, K.; Wiewrodt, R.; Molnar-Kimber, K.; Jovanovic, V.P.; Amin, K.M. An E2F-responsive replication-selective adenovirus targeted to the defective cell cycle in cancer cells: Potent antitumoral efficacy but no toxicity to normal cell. Cancer Res. 2002, 62, 3438–3447. [Google Scholar] [PubMed]
- Li, Y.; Yu, D.C.; Chen, Y.; Amin, P.; Zhang, H.; Nguyen, N.; Henderson, D.R. A hepatocellular carcinoma-specific adenovirus variant, CV890, eliminates distant human liver tumors in combination with doxorubicin. Cancer Res. 2001, 61, 6428–6436. [Google Scholar]
- Kirn, D.; Martuza, R.L.; Zwiebel, J. Replication-selective virotherapy for cancer: Biological principles, risk management and future directions. Nat. Med. 2001, 7, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Nemunaitis, J.; Ganly, I.; Khuri, F.; Arseneau, J.; Kuhn, J.; McCarty, T.; Landers, S.; Maples, P.; Romel, L.; Randlev, B.; et al. Selective replication and oncolysis in p53 mutant tumors with ONYX-015, an E1B-55kD gene-deleted adenovirus, in patients with advanced head and neck cancer: A phase II trial. Cancer Res. 2000, 60, 6359–6366. [Google Scholar]
- Jacobs, C.; Lyman, G.; Velez-García, E.; Sridhar, K.S.; Knight, W.; Hochster, H.; Goodnough, L.T.; Mortimer, J.E.; Einhorn, L.H.; Schacter, L. A phase III randomized study comparing cisplatin and fluorouracil as single agents and in combination for advanced squamous cell carcinoma of the head and neck. J. Clin. Oncol. 1992, 10, 257–263. [Google Scholar] [CrossRef]
- Vokes, E.E. Chemotherapy and integrated treatment approaches in head and neck cancer. Curr. Opin. Oncol. 1991, 3, 529–534. [Google Scholar] [CrossRef]
- Fujiwara, T.; Kagawa, S.; Kishimoto, H.; Endo, Y.; Hioki, M.; Ikeda, Y.; Sakai, R.; Urata, Y.; Tanaka, N.; Fujiwara, T. Enhanced antitumor efficacy of telomerase-selective oncolytic adenoviral agent OBP-401 with docetaxel: Preclinical evaluation of chemovirotherapy. Int. J. Cancer 2006, 119, 432–440. [Google Scholar] [CrossRef] [Green Version]
- Khuri, F.R.; Nemunaitis, J.; Ganly, I.; Arseneau, J.; Tannock, I.F.; Romel, L.; Gore, M.; Ironside, J.; MacDougall, R.H.; Heise, C.; et al. A controlled trial of intratumoral ONYX-015, a selectively-replicating adenovirus, in combination with cisplatin and 5-fluorouracil in patients with recurrent head and neck cancer. Nat. Med. 2000, 6, 879–885. [Google Scholar] [CrossRef]
- Cheong, S.C.; Wang, Y.; Meng, J.H.; Hill, R.; Sweeney, K.; Kirn, D.; Lemoine, N.R.; Hallden, G. E1A-expressing adenoviral E3B mutants act synergistically with chemotherapeutics in immunocompetent tumor models. Cancer Gene Ther. 2008, 15, 40–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenberg, D.P.; Adusumilli, P.S.; Hendershott, K.J.; Yu, Z.; Mullerad, M.; Chan, M.K.; Chou, T.C.; Fong, Y. 5-fluorouracil and gemcitabine potentiate the efficacy of oncolytic herpes viral gene therapy in the treatment of pancreatic cancer. J. Gastrointest. Surg. 2005, 9, 1068–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Post, D.E.; Fulci, G.; Chiocca, E.A.; Van Meir, E.G. Replicative oncolytic herpes simplex viruses in combination cancer therapies. Curr. Gene Ther. 2004, 4, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.F.; Gao, S.P.; Price, D.L.; Li, S.; Chou, T.C.; Singh, P.; Huang, Y.Y.; Fong, Y.; Wong, R.J. Synergy of a herpes oncolytic virus and paclitaxel for anaplastic thyroid cancer. Clin. Cancer Res. 2008, 14, 1519–1528. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, K.; Etoh, T.; Suzuki, K.; Mitui, M.T.; Nishizono, A.; Shiraishi, N.; Kitano, S. Efficacy of oncolytic reovirus against human gastric cancer with peritoneal metastasis in experimental animal model. Int. J. Oncol. 2010, 37, 1433–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.P.; Wang, C.Y.; Hsieh, P.Y.; Fang, J.H.; Chen, Y.L. Knockdown of serine/threonine-protein kinase 24 promotes tumorigenesis and myeloid-derived suppressor cell expansion in an orthotopic immunocompetent gastric cancer animal model. J. Cancer 2020, 11, 213–228. [Google Scholar] [CrossRef] [Green Version]
- Hamano, S.; Mori, Y.; Kataoka, H.; Tanaka, M.; Ebi, M.; Kubota, E.; Mizoshita, T.; Tanida, S.; Johnston, R.N.; Asai, K.; et al. Oncolytic reovirus combined with trastuzumab enhances antitumor efficacy through TRAIL signaling in human HER2-positive gastric cancer cells. Cancer Lett. 2015, 356, 846–854. [Google Scholar] [CrossRef]
- Alain, T.; Wong, J.F.; Urbanski, S.J.; Lee, P.W.; Muruve, D.A.; Johnston, R.N.; Forsyth, P.A.; Beck, P.L. Reovirus decreases azoxymethane-induced aberrant crypt foci and colon cancer in a rodent model. Cancer Gene Ther. 2007, 14, 867–872. [Google Scholar] [CrossRef]
- Mahalingam, D.; Wilkinson, G.A.; Eng, K.H.; Fields, P.; Raber, P.; Moseley, J.L.; Cheetham, K.; Coffey, M.; Nuovo, G.; Kalinski, P.; et al. Pembrolizumab in Combination with the Oncolytic Virus Pelareorep and Chemotherapy in Patients with Advanced Pancreatic Adenocarcinoma: A Phase Ib Study. Clin. Cancer Res. 2020, 26, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Mahalingam, D.; Goel, S.; Aparo, S.; Patel Arora, S.; Noronha, N.; Tran, H.; Chakrabarty, R.; Selvaggi, G.; Gutierrez, A.; Coffey, M.; et al. A Phase II Study of Pelareorep (REOLYSIN®) in Combination with Gemcitabine for Patients with Advanced Pancreatic Adenocarcinoma. Cancers 2018, 10, 160. [Google Scholar] [CrossRef] [Green Version]
- Shao, S.; Yang, X.; Zhang, Y.N.; Wang, X.J.; Li, K.; Zhao, Y.L.; Mou, X.Z.; Hu, P.Y. Oncolytic Virotherapy in Peritoneal Metastasis Gastric Cancer: The Challenges and Achievements. Front. Mol. Biosci. 2022, 28, 835300. [Google Scholar] [CrossRef]
- Mohamed, A.; Lin, Q.F.; Eaton, H.E.; Shmulevitz, M. p38 Mitogen-Activated Protein Kinase Signaling Enhances Reovirus Replication by Facilitating Efficient Virus Entry, Capsid Uncoating, and Postuncoating Steps. J. Virol. 2023, 97, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Zamarin, D.; Holmgaard, R.B.; Subudhi, S.K.; Park, J.S.; Mansour, M.; Palese, P.; Merghoub, T.; Wolchok, J.D.; Allison, J.P. Localized oncolytic virotherapy overcomes systemic tumor resistance to immune checkpoint blockade immunotherapy. Sci. Trans. Med. 2014, 6, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, A.; Johnston, R.N.; Shmulevitz, M. 2015. Potential for improving potency and specificity of reovirus oncolysis with next-generation reovirus variants. Viruses 2015, 7, 6251–6278. [Google Scholar] [CrossRef] [Green Version]
- Bourhill, T.; Mori, Y.; Rancourt, D.E.; Shmulevitz, M.; Johnston, R.N. Going (reo) viral: Factors promoting successful reoviral oncolytic infection. Viruses 2018, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Bourhill, T.; Rohani, L.; Kumar, M.; Bose, P.; Rancourt, D.; Johnston, R.N. Modulation of Reoviral Cytolysis (II): Cellular Stemness. Viruses 2023. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mori, Y.; Nishikawa, S.G.; Fratiloiu, A.R.; Tsutsui, M.; Kataoka, H.; Joh, T.; Johnston, R.N. Modulation of Reoviral Cytolysis (I): Combination Therapeutics. Viruses 2023, 15, 1472. https://doi.org/10.3390/v15071472
Mori Y, Nishikawa SG, Fratiloiu AR, Tsutsui M, Kataoka H, Joh T, Johnston RN. Modulation of Reoviral Cytolysis (I): Combination Therapeutics. Viruses. 2023; 15(7):1472. https://doi.org/10.3390/v15071472
Chicago/Turabian StyleMori, Yoshinori, Sandra G. Nishikawa, Andreea R. Fratiloiu, Mio Tsutsui, Hiromi Kataoka, Takashi Joh, and Randal N. Johnston. 2023. "Modulation of Reoviral Cytolysis (I): Combination Therapeutics" Viruses 15, no. 7: 1472. https://doi.org/10.3390/v15071472
APA StyleMori, Y., Nishikawa, S. G., Fratiloiu, A. R., Tsutsui, M., Kataoka, H., Joh, T., & Johnston, R. N. (2023). Modulation of Reoviral Cytolysis (I): Combination Therapeutics. Viruses, 15(7), 1472. https://doi.org/10.3390/v15071472