
The Extract of Scutellaria baicalensis Attenuates the Pattern Recognition Receptor Pathway Activation Induced by Influenza A Virus in Macrophages

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Virus and SBE Solution
2.2. Samples Preparation and RNA Extraction
2.3. Generation of cDNA Library and RNA Sequencing
2.4. Sequencing Data Quality Control and Genome Mapping
2.5. Gene Expression Analysis
2.6. Gene Ontology (GO) Term and Pathway Enrichments Analysis of DEGs
2.7. Real-Time Quantitative PCR (RT-qPCR)
2.8. Immunofluorescence under Confocal Laser Scanning Microscopy
2.9. Western Blot Assay
2.10. Quantification and Statistical Analysis
3. Results
3.1. SBE Plays Dual Role of Antivirus and Inflammatory Inhibition in Macrophage Infected with PR8 Strain
3.2. Transcriptome Analysis of the Cellular Gene Expression after Virus Infection and Intervention Effect of SBE
3.3. Differentially Expressed Genes (DEGs) Analysis
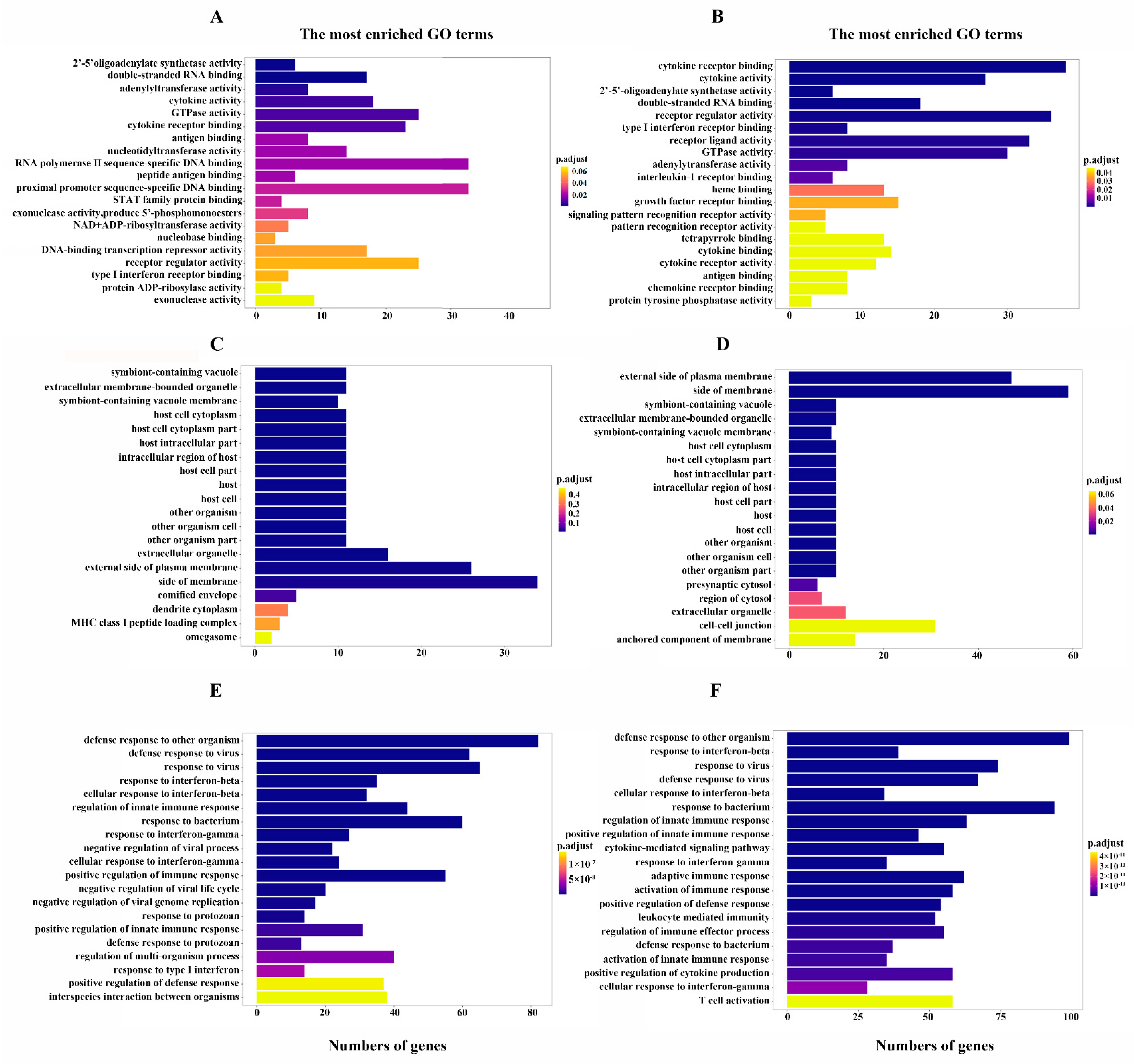
3.4. SBE Inhibits the Activation of Innate Immune Response after Virus Infection Revealed by GO Enrichment Analysis
3.5. SBE Interferes with Pattern Recognition Receptor Signaling Pathway Based on KEGG Pathway Enrichment Analysis
3.6. Validation of Key DEGs within PRR Signaling Pathway and Detection of Downstream Effect
4. Discussion
4.1. The Over-Activation of PRR Pathways Leads to the Aggravation of Inflammation Induced by IAV Infection
4.2. SBE Effectively Reverses the Over-Transcription of Genes in PRR Signaling Pathways and Inflammatory Response Induced by IAV through Multiple Targets
4.3. Multi-Target Reversal Drug Screening and Development Based on Gene Transcription Map of IAV Infection
4.4. Intervention Macrophage and “Cytokine Storm” Is Crucial for Treatment of Viral Pneumonia
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paules, C.; Subbarao, K. Influenza. Lancet 2017, 390, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Govorkova, E.A. H5N1 influenza-continuing evolution and spread. N. Engl. J. Med. 2006, 355, 2174–2177. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wu, Y. Bat-derived influenza-like viruses H17N10 and H18N11. Trends Microbiol. 2014, 22, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Mauad, T.; Hajjar, L.A. Lung pathology in fatal novel human influenza A (H1N1) infection. Am. J. Respir. Crit. Care Med. 2010, 181, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Rello, J.; Pop-Vicas, A. Clinical review: Primary influenza viral pneumonia. Crit. Care 2009, 13, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uyeki, T.M.; Hui, D.S. Influenza. Lancet 2020, 400, 693–706. [Google Scholar] [CrossRef]
- Sparrow, E.; Wood, J.G. Global production capacity of seasonal and pandemic influenza vaccines in 2019. Vaccine 2021, 39, 512–520. [Google Scholar] [CrossRef]
- Peteranderl, C.; Herold, S. Human Influenza Virus Infections. Semin. Respir. Crit. Care Med. 2016, 37, 487–500. [Google Scholar] [CrossRef]
- Zhao, T.; Tang, H. Scutellaria baicalensis Georgi. (Lamiaceae): A review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. J. Pharm. Pharmacol. 2019, 71, 1353–1369. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.L.; Wang, S. A comprehensive review on phytochemistry, pharmacology, and flavonoid biosynthesis of Scutellaria baicalensis. Pharm. Biol. 2018, 56, 465–484. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Wang, D. Whole genome and transcriptome reveal flavone accumulation in Scutellaria baicalensis roots. Front. Plant Sci. 2022, 13, 1000469. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Li, P. Pharmacophylogenetic study of Scutellaria baicalensis and its substitute medicinal species based on the chloroplast genomics, metabolomics, and active ingredient. Front. Plant Sci. 2022, 13, 951824. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.W.; Lin, T.Y. Therapeutic efficacy of Scutellaria baicalensis Georgi against psoriasis-like lesions via regulating the responses of keratinocyte and macrophage. Biomed. Pharmacother. 2022, 155, 113798. [Google Scholar] [CrossRef] [PubMed]
- Zhi, H.J.; Zhu, H.Y. In vivo effect of quantified flavonoids-enriched extract of Scutellaria baicalensis root on acute lung injury induced by influenza A virus. Phytomedicine 2019, 57, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Zhi, H.; Jin, X. Exploring the effective materials of flavonoids-enriched extract from Scutellaria baicalensis roots based on the metabolic activation in influenza A virus induced acute lung injury. J. Pharm. Biomed. Anal. 2020, 177, 112876. [Google Scholar] [CrossRef]
- Xu, H.R.; Li, Y.L. Effect of Scutellariae Radix on expression of inflammatory cytokine protein and gene in lung of mice with viral pneumonia caused by influenza virus FM1 infection. Zhongguo Zhong Yao Za Zhi 2019, 44, 5166–5173. [Google Scholar] [PubMed]
- Zhang, Q.; Yang, B. Effect of total flavonoids of Scutellaria baicalensis Georgi on expression of influenza A virus nucleoprotein in HeLa cells. Nan Fang Yi Ke Da Xue Xue Bao 2012, 32, 966–969. [Google Scholar] [PubMed]
- Jin, J.; Chen, Y. The inhibitory effect of sodium baicalin on oseltamivir-resistant influenza A virus via reduction of neuraminidase activity. Arch. Pharm. Res. 2018, 41, 664–676. [Google Scholar] [CrossRef]
- Li, R.; Wang, L. Baicalin inhibits influenza virus A replication via activation of type I IFN signaling by reducing miR-146a. Mol. Med. Rep. 2019, 20, 5041–5049. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Nikrad, M.P. Innate immune response of human alveolar macrophages during influenza A infection. PLoS ONE 2012, 7, e29879. [Google Scholar] [CrossRef] [Green Version]
- Vangeti, S.; Yu, M. Respiratory Mononuclear Phagocytes in Human influenza A virus infection: Their Role in immune Protection and As Targets of the virus. Front. Immunol. 2018, 9, 1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, B.; Wu, Y. Uncoupling of macrophage inflammation from self-renewal modulates host recovery from respiratory viral infection. Immunity 2021, 54, 1200–1218.e9. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, G.; Zhou, Y. Innate Immune Sensing of Influenza A Virus. Viruses 2020, 12, 755. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhang, L. Host cellular signaling induced by influenza virus. Sci. China Life Sci. 2011, 54, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Chhibber, S. Acute lung inflammation in Klebsiella pneumoniae B5055-induced pneumonia and sepsis in BALB/c mice: A comparative study. Inflammation 2011, 34, 452–462. [Google Scholar] [CrossRef]
- Guirgis, F.W.; Khadpe, J.D. Persistent organ dysfunction after severe sepsis: A systematic review. J. Crit. Care 2014, 29, 320–326. [Google Scholar] [CrossRef]
- Le Goffic, R.; Balloy, V. Detrimental contribution of the Toll-like receptor (TLR)3 to influenza A virus-induced acute pneumonia. PLoS Pathog. 2006, 2, e53. [Google Scholar] [CrossRef] [Green Version]
- de Marcken, M.; Dhaliwal, K. TLR7 and TLR8 activate distinct pathways in monocytes during RNA virus infection. Sci. Signal. 2019, 12, eaaw1347. [Google Scholar] [CrossRef]
- Pichlmair, A.; Schulz, O. RIG-I-mediated antiviral responses to single-stranded RNA bearing 5′-phosphates. Science 2006, 314, 997–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benitez, A.A.; Panis, M. In Vivo RNAi Screening Identifies MDA5 as a Significant Contributor to the Cellular Defense against Influenza A Virus. Cell Rep. 2015, 11, 1714–1726. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.G.; Dash, P. The Intracellular Sensor NLRP3 Mediates Key Innate and Healing Responses to Influenza A Virus via the Regulation of Caspase-1. Immunity 2009, 30, 566–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriyama, M.; Koshiba, T. Influenza A virus M2 protein triggers mitochondrial DNA-mediated antiviral immune responses. Nat. Commun. 2019, 10, 4624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullen, L.M.; Chamberlain, G. Pattern recognition receptors as potential therapeutic targets in inflammatory rheumatic disease. Arthritis Res. Ther. 2015, 17, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.P.; Christopher, M.E. Activation of toll-like receptor signaling pathway for protection against influenza virus infection. Vaccine 2009, 27, 3481–3483. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.P.; Christopher, M.E. Antiviral role of toll-like receptor-3 agonists against seasonal and avian influenza viruses. Curr. Pharm. Des. 2009, 15, 1269–1274. [Google Scholar] [CrossRef] [PubMed]
- Green, R.R.; Wilkins, C. Transcriptional analysis of antiviral small molecule therapeutics as agonists of the RLR pathway. Genom. Data 2016, 7, 290–292. [Google Scholar] [CrossRef] [Green Version]
- Pattabhi, S.; Wilkins, C.R. Targeting Innate Immunity for Antiviral Therapy through Small Molecule Agonists of the RLR Pathway. J. Virol. 2015, 90, 2372–2387. [Google Scholar] [CrossRef] [Green Version]
- Li, C.R.; Lin, G. Pharmacological effects and pharmacokinetics properties of Radix Scutellariae and its bioactive flavones. Biopharm. Drug Dispos. 2011, 32, 427–445. [Google Scholar] [CrossRef]
- Zhu, H.Y.; Han, L. Baicalin inhibits autophagy induced by influenza A virus H3N2. Antivir. Res. 2015, 113, 62–70. [Google Scholar] [CrossRef]
- Chu, M.; Xu, L. Role of Baicalin in Anti-Influenza Virus A as a Potent Inducer of IFN-Gamma. Biomed. Res. Int. 2015, 2015, 263630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.H.; Yu, W.Y. Mosla scabra flavonoids ameliorate the influenza A virus-induced lung injury and water transport abnormality via the inhibition of PRR and AQP signaling pathways in mice. J. Ethnopharmacol. 2016, 179, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.X.; Li, R.F. Total alkaloids from Alstonia scholaris inhibit influenza a virus replication and lung immunopathology by regulating the innate immune response. Phytomedicine 2020, 77, 153272. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Hu, Z.L. Deep sequencing of the mouse lung transcriptome reveals distinct long non-coding RNAs expression associated with the high virulence of H5N1 avian influenza virus in mice. Virulence 2018, 9, 1092–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bavagnoli, L.; Campanini, G. Identification of a novel antiviral micro-RNA targeting the NS1 protein of the H1N1 pandemic human influenza virus and a corresponding viral escape mutation. Antivir. Res. 2019, 171, 104593. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Zhang, Y.B. CircRNA expression profiling and bioinformatics analysis indicate the potential biological role and clinical significance of circRNA in influenza A virus-induced lung injury. J. Biosci. 2021, 46, 38. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.S.; Andiappan, A.K. RNA Sequencing of H3N2 Influenza Virus-Infected Human Nasal Epithelial Cells from Multiple Subjects Reveals Molecular Pathways Associated with Tissue Injury and Complications. Cells 2019, 8, 986. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Zhang, K. Global transcriptome analysis of H5N1 influenza virus-infected human cells. Hereditas 2019, 156, 10. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.X.; Nagy, A. RNA-Seq Analysis of Influenza A Virus-Induced Transcriptional Changes in Mice Lung and Its Possible Implications for the Virus Pathogenicity in Mice. Viruses 2021, 13, 2031. [Google Scholar] [CrossRef]
- Ural, B.B.; Yeung, S.T. Identification of a nerve-associated, lung-resident interstitial macrophage subset with distinct localization and immunoregulatory properties. Sci. Immunol. 2020, 5, eaax8756. [Google Scholar] [CrossRef]
- Nicol, M.Q.; Dutia, B.M. The role of macrophages in influenza A virus infection. Future Virol. 2014, 9, 847–862. [Google Scholar] [CrossRef]
- Gu, Y.; Zuo, X. The Mechanism behind Influenza Virus Cytokine Storm. Viruses 2021, 13, 1362. [Google Scholar] [CrossRef] [PubMed]
- Kalil, A.C.; Thomas, P.G. Influenza virus-related critical illness: Pathophysiology and epidemiology. Critical Care 2019, 23, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Us, D. Cytokine storm in avian influenza. Mikrobiyol. Bul. 2008, 42, 365–380. [Google Scholar]
- Sun, J.; Madan, R. Effector T cells control lung inflammation during acute influenza virus infection by producing IL-10. Nat. Med. 2009, 15, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.L.; Zhang, X. Intensive Cytokine induction in Pandemic H1N1 Influenza Virus Infection Accompanied by Robust Production of IL-10 and IL-6. PLoS ONE 2011, 6, e28680. [Google Scholar] [CrossRef] [Green Version]
- Xiahou, Z.K.; Wang, X.L. NMI and IFP35 serve as proinflammatory DAMPs during cellular infection and injury. Nat. Commun. 2017, 8, 950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Xu, N. IFP35 as a promising biomarker and therapeutic target for the syndromes induced by SARS-CoV-2 or influenza virus. Cell Rep. 2021, 37, 110126. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B.; Maddur, M.S. Innate immune sensing and response to influenza. Curr. Top. Microbiol. Immunol. 2015, 386, 23–71. [Google Scholar]
- Liu, G.; Chen, S. The Establishment and Validation of the Human U937 Cell Line as a Cellular Model to Screen Immunomodulatory Agents Regulating Cytokine Release Induced by Influenza Virus Infection. Virol. Sin. 2019, 34, 648–661. [Google Scholar] [CrossRef] [Green Version]
- Li, R.F.; Zhou, X.B. Novel fatty acid in cordyceps suppresses influenza A (H1N1) virus induced proinflammatory response through regulating innate signaling pathways. ACS Omega 2021, 6, 1505–1515. [Google Scholar] [CrossRef] [PubMed]
- Ramos, I.; Fernandez-Sesma, A. Modulating the innate immune response to influenza A virus: Potential therapeutic use of anti-inflammatory drugs. Front. Immunol. 2015, 6, 361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.L.; Ke, C. Scutellaria baicalensis pith-decayed root inhibits macrophage-related inflammation through the NF-κB/NLRP3 pathway to alleviate LPS-induced acute lung injury. Planta Med. 2023, 89, 493–507. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Ma, L.; Su, R.; Guo, R.; Zhou, N.; Liu, M.; Wu, J.; Wang, Y.; Hao, Y. The Extract of Scutellaria baicalensis Attenuates the Pattern Recognition Receptor Pathway Activation Induced by Influenza A Virus in Macrophages. Viruses 2023, 15, 1524. https://doi.org/10.3390/v15071524
Yang M, Ma L, Su R, Guo R, Zhou N, Liu M, Wu J, Wang Y, Hao Y. The Extract of Scutellaria baicalensis Attenuates the Pattern Recognition Receptor Pathway Activation Induced by Influenza A Virus in Macrophages. Viruses. 2023; 15(7):1524. https://doi.org/10.3390/v15071524
Chicago/Turabian StyleYang, Mingrui, Luyao Ma, Rina Su, Rui Guo, Na Zhou, Menghua Liu, Jun Wu, Yi Wang, and Yu Hao. 2023. "The Extract of Scutellaria baicalensis Attenuates the Pattern Recognition Receptor Pathway Activation Induced by Influenza A Virus in Macrophages" Viruses 15, no. 7: 1524. https://doi.org/10.3390/v15071524
APA StyleYang, M., Ma, L., Su, R., Guo, R., Zhou, N., Liu, M., Wu, J., Wang, Y., & Hao, Y. (2023). The Extract of Scutellaria baicalensis Attenuates the Pattern Recognition Receptor Pathway Activation Induced by Influenza A Virus in Macrophages. Viruses, 15(7), 1524. https://doi.org/10.3390/v15071524