
Current Achievements and Future Prospects in Virus Elimination Technology for Functional Chrysanthemum

 ,
,
Abstract
:1. Introduction
2. Research Progress on Chrysanthemum Viral Diseases
Main Virus Species Infecting Functional Chrysanthemum
3. Research Progress in Chrysanthemum Virus Detection Technology
3.1. Indicator Plant Assay
3.2. Electron Microscope Observation
3.3. Enzyme-Linked Immunosorbent Assay
3.4. Molecular Biology Technology
3.4.1. Reverse Transcription Polymerase Chain Reaction (RT-PCR) and Its Related Detection Techniques
3.4.2. Loop-Mediated Isothermal Amplification (LAMP)
4. Research Progress on Chrysanthemum Virus Elimination Methods
4.1. Shoot Tip Elimination
4.2. Heat Treatment Elimination
4.3. Antiviral Agent Treatment Elimination
4.4. Ultralow-Temperature Treatment
5. Research Progress on the Propagation of Virus-Free Chrysanthemum Seedlings
5.1. Sterile Seedling Culture
5.2. Propagation of Virus-Free Seedlings
5.3. Rooting Induction
5.4. Seedling Cultivation and Transplanting
5.5. Evaluation of the Elimination Effect
6. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, S.L.; Dai, S.L. The Chinese Chrysanthemum Book; China Forestry Publishing House: Beijing, China, 2013. [Google Scholar]
- Zhang, D.C. Edible Chrysanthemum. Chin. Veg. 2017, 6, 61. [Google Scholar]
- Wang, T.C. Chrysanthemum and its cultivation. Xinjiang For. 2020, 4, 37–38. [Google Scholar]
- Su, J.; Jiang, J.; Zhang, F.; Liu, Y.; Ding, L.; Chen, S.; Chen, F. Current achievements and future prospects in the genetic breeding of chrysanthemum: A review. Hortic. Res. 2019, 6, 109. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.; Wang, T.; Guo, Q.; Yang, F.; Chen, J.; Zhang, W. Combined metabolomic and transcriptomic analysis reveals redirection of the phenylpropanoid metabolic flux in different colored medicinal Chrysanthemum morifolium. Ind. Crops Prod. 2021, 164, 113343. [Google Scholar] [CrossRef]
- Zhou, L.J.; Wang, Y.; Wang, Y.; Song, A.; Jiang, J.; Chen, S.; Ding, B.; Guan, Z.; Chen, F. Transcription factor CmbHLH16 regulates petal anthocyanin homeostasis under different lights in chrysanthemum. Plant Physiol. 2022, 190, 1134–1152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.M.; Chen, R.; Xian, X.L. Research progress of functional chrysanthemum in China. Chin. J. Agric. Sci. 2022, 38, 38–46. [Google Scholar]
- Lu, C.; Li, Y.; Wang, J.; Qu, J.; Chen, Y.; Chen, X.; Huang, H.; Dai, S. Flower color classification and correlation between color space values with pigments in potted multiflora chrysanthemum. Sci. Hortic. 2021, 283, 110082. [Google Scholar] [CrossRef]
- Wang, T.; Guo, Q.S.; Mao, P.F. Flavonoid accumulation during florescence in three Chrysanthemum morifolium Ramat. ‘Hangbai’ genotypes. Biochem. Syst. Ecol. 2014, 55, 79–83. [Google Scholar] [CrossRef]
- Yu, Q.; Chen, W.; Zhong, J.; Qing, D.; Yan, C. Structural elucidation of three novel oligosaccharides from ‘Kunlun’ Chrysanthemum flower tea and their bioactivities. Food Chem. Toxicol. 2021, 149, 112032. [Google Scholar] [CrossRef]
- Yuan, H.; Jiang, S.; Liu, Y.; Daniyal, M.; Jian, Y.; Peng, C.; Shen, J.; Liu, S.; Wang, W. The flower head of Chrysanthemum morifolium Ramat: A paradigm of flowers serving as Chinese dietary herbal medicine. J. Ethnopharmacol. 2020, 261, 113043. [Google Scholar] [CrossRef]
- Wang, R.R. Study on Cryopreservation Technology and Virulence Elimination of Germplasm Resources of Chrysanthemum, Jujube and Potato. Ph.D. Thesis, Northwest A&F University, Xianyang, China, 2014. [Google Scholar]
- Gao, H. Effects of trace elements in Chrysanthemum on anti-inflammatory activity. J. Tradit. Chin. Med. Manag. 2006, 1, 24–25. [Google Scholar]
- Jiang, J.R. Reserch into the Effect and Mechanism of WCWE on HepG2 Cells. Ph.D. Thesis, Chengdu University of Traditional Chinese Medicine, Chengdu, China, 2017. [Google Scholar]
- Liu, D.M.; Zhu, S.M.; Yang, H.K.; Zhou, J.; Dong, X.R. Optimization of the extraction process of antioxidant active components from Huangshan Tributary Chrysanthemum and analysis of it by HPLC-MS. Sci. Technol. Food Ind. 2020, 41, 150–156. [Google Scholar]
- Jiang, H.D. Study on Cardiovascular System and Active Components of ‘Hangbai’ Chrysanthemum. Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2004. [Google Scholar]
- Yuan, L.H. Research progress on antitumor effect of apigenin. China Public Health 2004, 2, 117–118. [Google Scholar]
- Hadizadeh, H.; Samiei, L.; Shakeri, A. Chrysanthemum, an ornamental genus with considerable medicinal value: A comprehensive review. S. Afr. J. Bot. 2022, 144, 23–43. [Google Scholar] [CrossRef]
- Zhong, X.; Yang, L.; Li, J.; Tang, Z.; Wu, C.; Zhang, L.; Zhou, X.; Wang, Y.; Wang, Z. Integrated next-generation sequencing and comparative transcriptomic analysis of leaves provides novel insights into the ethylene pathway of Chrysanthemum morifolium in response to a Chinese isolate of chrysanthemum virus B. Virol. J. 2022, 19, 182. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.D.; Miao, M.S. Modern research and functional characteristics of chrysanthemum. J. Tradit. Chin. Med. 2014, 29, 551–553. [Google Scholar]
- Wang, Y.H. Establishment of Virus-Free System of Three Chrysanthemum Cultivars for Tea. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2019. [Google Scholar]
- Zhan, G.; Sun, M.Y.; Li, J.; Qu, F.L.; Long, M. Amino acid composition and nutritional value evaluation of five chrysanthemum species from different habitats. J. Anhui Agric. Univ. 2019, 46, 908–914. [Google Scholar]
- Cheng, C.G.; Li, D.T.; Liu, X.H.; Hong, Q.H. Comparison of Trace elements of Chrysanthemum and Chrysanthemum. Spectrosc. Spectr. Anal. 2006, 1, 156–158. [Google Scholar]
- Li, J.; Wu, X.; Liu, H.; Wang, X.; Yi, S.; Zhong, X.; Wang, Y.; Wang, Z. Identification and Molecular Characterization of a Novel Carlavirus Infecting Chrysanthemum morifolium in China. Viruses 2023, 15, 1029. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, X.; Ge, B.; Li, M.; Hong, B. A multiplex RT-PCR for simultaneous detection and identification of five viruses and two viroids infecting chrysanthemum. Arch. Virol. 2015, 160, 1145–1152. [Google Scholar] [CrossRef]
- Gobatto, D.; Oliveira, L.A.; Siqueira Franco, D.A.; Velásquez, N.; Daròs, J.A.; Eiras, M. Surveys in the chrysanthemum production areas of Brazil and Colombia reveal that weeds are potential reservoirs of chrysanthemum stunt viroid. Viruses 2019, 11, 355. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Gupta, A.; Jain, S.; Singh, D.; Verma, N. Chrysanthemum production, viral diseases and their management. In Virus Diseases of Ornamental Plants; Raj, S.K., Gaur, R.K., Yin, Z., Eds.; Springer: Singapore, 2021; pp. 261–275. [Google Scholar]
- Chirkov, S.N.; Sheveleva, A.; Snezhkina, A.; Kudryavtseva, A.; Krasnov, G.; Zakubanskiy, A.; Mitrofanova, I. Highly divergent isolates of chrysanthemum virus B and chrysanthemum virus R infecting chrysanthemum in Russia. PeerJ 2022, 10, e12607. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Xi, Y.; Shen, C.; Wang, M.; Wang, H. Occurrence of Nigrospora sphaerica causing leaf blight on Chrysanthemum morifolium in China. Crop Prot. 2022, 157, 105982. [Google Scholar] [CrossRef]
- Mitiouchkina, T.Y.; Firsov, A.P.; Titova, S.M.; Pushin, A.S.; Shulga, O.A.; Dolgov, S.V. Different Approaches to Produce Transgenic Virus B Resistant Chrysanthemum. Agronomy 2018, 8, 28. [Google Scholar] [CrossRef]
- Liu, P.L.; Wan, Q.; Guo, Y.P.; Yang, J.; Rao, G.Y. Phylogeny of the genus Chrysanthemum L.: Evidence from single-copy nuclear gene and chloroplast DNA sequences. PLoS ONE 2012, 7, e48970. [Google Scholar] [CrossRef] [PubMed]
- Dowrick, G.; El-Bayoumi, A. The origin of new forms of the garden Chrysanthemum. Euphytica 1966, 15, 32–38. [Google Scholar] [CrossRef]
- Nakano, M.; Hirakawa, H.; Fukai, E.; Toyoda, A.; Kajitani, R.; Minakuchi, Y.; Itoh, T.; Higuchi, Y.; Kozuka, T.; Bono, H.; et al. A chromosome-level genome sequence of Chrysanthemum seticuspe, a model species for hexaploid cultivated chrysanthemum. Commun. Biol. 2021, 4, 1167. [Google Scholar] [CrossRef]
- Eisa, E.A.; Tilly-Mándy, A.; Honfi, P.; Shala, A.Y.; Gururani, M.A. Chrysanthemum: A comprehensive review on recent developments on in vitro regeneration. Biology 2022, 11, 1774. [Google Scholar] [CrossRef]
- Guan, Y.; He, X.; Wen, D.; Chen, S.; Chen, F.; Chen, F.; Jiang, Y. Fusarium oxysporum infection on root elicit aboveground terpene production and salicylic acid accumulation in Chrysanthemum morifolium. Plant Physiol. Biochem. 2022, 190, 11–23. [Google Scholar] [CrossRef]
- Cho, W.K.; Jo, Y.; Jo, K.-M.; Kim, K.-H. A Current overview of two viroids that infect Chrysanthemums: Chrysanthemum stunt viroid and Chrysanthemum chlorotic mottle viroid. Viruses 2013, 5, 1099–1113. [Google Scholar] [CrossRef]
- Palukaitis, P. Chrysanthemum stunt viroid. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Elsevier: London, UK, 2017; pp. 181–190. ISBN 9780128014981. [Google Scholar]
- Verhoeven, J.T.J.; Hammond, R.W.; Stancanelli, G. Economic significance of viroids in ornamental crops. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Elsevier: London, UK, 2017; pp. 27–38. ISBN 9780128014981. [Google Scholar]
- Liu, H.H. Virus Pathogen Identification and Analysis of Biological Characters and Quality Characters of Virus-Free Chrysanthemum Seedlings. Master’s Thesis, Zhejiang University, Hangzhou, China, 2015. [Google Scholar]
- Xiao, X.; Zhu, W.; Du, C.; Shi, Y.D.; Wang, J.F. Effects of crop rotation and fertilization on microbial characteristics of ‘Chuzhou’ Chrysanthemum continuous cropping soil. Chin. J. Appl. Ecol. 2015, 26, 1779–1784. [Google Scholar]
- Gobatto, D.; Chaves, A.L.R.; Harakava, R.; Marque, J.M.; Daròs, J.A.; Eiras, M. Chrysanthemum stunt viroid in Brazil: Survey, identification, biological and molecular characterization and detection methods. J. Plant Pathol. 2014, 96, 111–119. [Google Scholar]
- Yoon, J.Y.; Baek, E.; Palukaitis, P. Are there strains of Chrysanthemum stunt viroid? J. Plant Pathol. 2012, 94, 697–701. [Google Scholar]
- Yoon, J.Y.; Palukaitis, P. Sequence comparisons of global Chrysanthemum stunt viroid variants: Multiple polymorphic positions scattered through the viroid genome. Virus Genes 2013, 46, 97–104. [Google Scholar] [CrossRef]
- Yan, K.; Du, X.; Mao, B. Production of virus-free chrysanthemum (Chrysanthemum morifolium Ramat) by tissue culture techniques. Methods Mol. Biol. 2022, 2400, 171–186. [Google Scholar] [PubMed]
- Liu, X.L.; Zhao, X.T.; Muhammad, I.; Ge, B.B.; Hong, B. Multiplex reverse transcription loop-mediated isothermal amplification for the simultaneous detection of CVB and CSVd in chrysanthemum. J. Virol. Methods 2014, 210, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Li, H.N. Study on Gene Expression of Citrus Clefts Virus Infection. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2014. [Google Scholar]
- Liu, X.L.; Wei, Q.; Hong, B.; Zhao, X.T. First report of potato virus Y strain N-Wilga infecting chrysanthemum in China. Plant Dis. 2014, 98, 1589. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, D.; Yu, C.; Wang, Z.; Dong, J.; Shi, K.; Yuan, X. Identification of three new isolates of tomato spotted wilt virus from different hosts in China: Molecular diversity, phylogenetic and recombination analyses. Virol. J. 2016, 13, 8. [Google Scholar] [CrossRef]
- Diener, T.O.; Lawson, R.H. Chrysanthemum stunt: A viroid disease. Virology 1973, 51, 94–101. [Google Scholar] [CrossRef]
- Hollings, M.; Stone, O.M. Some properties of chrysanthemum stunt, a virus with the characteristics of an uncoated ribonucleic acid. Ann. Appl. Biol. 1973, 74, 333–348. [Google Scholar] [CrossRef]
- Palukaitis, P.; Symons, R.H. Purification and characterization of the circular and linear forms of chrysanthemum stunt viroid. J. Gen. Virol. 1980, 46, 477–489. [Google Scholar] [CrossRef]
- Zhang, Z.Z.; Pan, S.; Li, S.F. First report of chrysanthemum chlorotic mottle viroid in chrysanthemum in China. Plant Dis. 2011, 95, 1320. [Google Scholar] [CrossRef] [PubMed]
- Singh, L.; Hallan, V.; Zaidi, A.A. Intermolecular interactions of chrysanthemum virus B coat protein: Implications for capsid assembly. Indian J. Virol. 2011, 22, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Matsyshita, Y. Chrysanthemum stunt viroid. Jpn. Agric. Res. Q. JARQ 2013, 47, 237–242. [Google Scholar] [CrossRef]
- Gottsberger, A.R.; Suárez-Mahecha, B. Detection of Citrus exocortis viroid on Solanum jasminoides plantlets from an Austrian nursery. Plant Pathol. 2010, 59, 1159. [Google Scholar] [CrossRef]
- Wang, R.; Dong, J.; Wang, Z.; Zhou, T.; Li, Y.; Ding, W.L. Complete nucleotide sequence of a new carlavirus in Chrysanthemums in China. Arch. Virol. 2018, 163, 1973–1976. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Jo, Y.; Lian, S.; Jo, K.M.; Chu, H.; Yoon, J.Y.; Choi, S.K.; Kim, K.H.; Cho, W.K. Comparative analysis of chrysanthemum transcriptome in response to three RNA viruses: Cucumber mosaic virus, Tomato spotted wilt virus and Potato virus X. Plant Mol Biol. 2015, 88, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.M.; Moran, J.R. The incidence of chrysanthemum virus B, tomato aspermy virus and tomato spotted wilt virus in commercial chrysanthemum flowercrops in Victoria. Australas. Plant Pathol. 1983, 12, 17. [Google Scholar] [CrossRef]
- Zeng, R. Quantitative Analysis of Genomic RNA of Cucumber Mosaic Virus. Master’s Thesis, Hunan Agricultural University, Changsha, China, 2007. [Google Scholar]
- Qu, J.; Guo, X.Q.; Ci, X.Y.; Qi, D.; Wen, F.J. Advances in molecular biology of potato X virus and its application as expression vector. Chin. J. Virol. 2003, 1, 87–92. [Google Scholar]
- Sun, Q. Study on RT-PCR Detection Technology of Virus-Free Potato Seedlings. Master’s Thesis, Shandong Agricultural University, Tai’an, China, 2003. [Google Scholar]
- Kondo, T.; Yamashita, K.; Sugiyama, S. First report of impatiens necrotic spot virus infecting chrysanthemum (Chrysanthemum morifolium) in Japan. J. Gen. Plant Pathol. 2011, 77, 263–265. [Google Scholar] [CrossRef]
- Sherman, J.M.; Moyer, J.W.; Daub, M.E. Tomato spotted wilt virus resistance in chrysanthemum expressing the viral nucleocapsid gene. Plant Dis. 1998, 82, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Y.; Cai, J.H.; Li, G.X.; Qin, B.X.; Zhou, X.P. Molecular characterization of a distinct Begomovirus infecting Euphorbia pulcherrima in China. J. Phytopathol. 2004, 152, 215–218. [Google Scholar] [CrossRef]
- An, X.L. Preliminary Study on Yellow Dwarf Disease of Potato in North Xinjiang. Master’s Thesis, Shihezi University, Shihezi, China, 2008. [Google Scholar]
- Raj, S.K.; Kumar, S.; Choudhari, S. Identification of Tomato aspermy virus as the cause of yellow mosaic and flower deformation of chrysanthemums in India. Australas. Plant Dis. Notes 2007, 2, 1–2. [Google Scholar] [CrossRef]
- Takeshita, M.; Nagai, N.; Okuda, M.; Matsuura, S.; Okuda, S.; Furuya, N.; Tsuchiya, K. Molecular and biological characterization of Chrysanthemum stem necrosis virus isolates from distinct regions in Japan. Eur. J. Plant Pathol. 2011, 131, 9–14. [Google Scholar] [CrossRef]
- Zhang, E.Z.; Zhao, W.; Zhang, L.L.; Han, Q.Y.; Zhang, J.H. Advances in cloning of Soybean Mosaic virus infection. Soybean Sci. 2017, 36, 808–812. [Google Scholar]
- Li, L.J. Rapid Detection and Biological Control of Tobacco Mosaic virus in Southern Hunan. Master’s Thesis, Hunan Agricultural University, Changsha, China, 2015. [Google Scholar]
- Matsui, C.; Hosaka, Y.; Kawase, S. Map of Plant Virus; Beijing Agricultural Press: Beijing, China, 1980; pp. 103–105. [Google Scholar]
- Liu, H.H.; Shen, X.G.; Mao, B.Z. Chrysanthemum virus disease and its prevention and treatment. Pharm. Biotechnol. 2015, 22, 91–94. [Google Scholar]
- Wang, T.L.; Qin, L.; Du, Y.X.; Sun, K.; Wang, S.; He, Z. Research progress of Chrysanthemum virus disease. Chin. J. Mod. Chin. Med. 2019, 23, 1655–1663. [Google Scholar]
- Chen, Y.C.; Nie, F.J.; Zhang, L.; Gong, L.; Gan, X.Y.; Shi, L.; Song, Y.X. Research progress of potato X. virus. Chang. Veg. 2016, 18, 39–44. [Google Scholar]
- Kumar, S.; Khan, M.S.; Raj, S.K.; Sharma, A.K. Elimination of mixed infection of Cucumber mosaic and Tomato aspermy virus from Chrysanthemum morifolium Ramat. cv. Pooja by shoot meristem culture. Sci. Hortic. 2009, 119, 108–112. [Google Scholar] [CrossRef]
- Li, M.Y.; Chen, D.L.; Li, X.M.; Wang, L.L.; Huang, C.L. Tomato spotted wilt virus (TSWV) occurred in the chrysanthemum of Li Mingyuan (85). Agric. Eng. Technol. 2018, 38, 61–64. [Google Scholar]
- Li, M.J.; Yang, X.C.; Tian, B. Identification of chlorotic mottle viroids in Chrysanthemum. Acta Microbiol. Sin. 1987, 2, 31–37. [Google Scholar]
- Li, M.Y.; Chen, D.L.; Li, X.M.; Jiang, M.Y. Identification of two species of Chrysanthemum virus from Li Mingyuan’s broken hand (81). J. Agric. Eng. Technol. 2017, 37, 62–64. [Google Scholar]
- Nie, X.; Singh, R.P. A novel usage of random primers for multiplex RT-PCR detection of virus and viroid in aphids, leaves, and tubers. J. Virol. Methods 2001, 91, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.N.; Pak, H.S.; Jung, J.A.; Kim, J.S. Occurrence of Tomato spotted wilt virus in Chrysanthemum (Dendranthema grandiflorum) in Korea. Plant Pathol. J. 2006, 22, 230. [Google Scholar] [CrossRef]
- Leng, X.X. Virus-free culture and detection of flower shoot tip. Biol. Bull. 2003, 03, 14. [Google Scholar]
- Zhao, S. Study on Virus Elimination and Detection Technology of Chrysanthemum. Master’s Thesis, Beijing Forestry University, Beijing, China, 2013. [Google Scholar]
- Noordam, D. Virus diseases of Chrysanthemum indicum in The Netherlands. Tijdschr. Plantenziekten 1952, 58, 121–190. [Google Scholar]
- Chen, J.F.; Zhao, J.Y. Detection of virus in Li Shaohua fruit trees and its application. Hebei For. Fruit Res. 2005, 20, 167–173. [Google Scholar]
- Cai, J.H.; Zhu, X.R. Identification of virus pathogens of several Chrysanthemum. Guangdong Agric. Sci. 1994, 6, 282–283. [Google Scholar]
- Chen, Y.F.; Li, M.F.; Li, G.F.; Wei, M.S.; Yi, F.Y.; Han, L.B. Identification of virus-free seedlings from mother strains of Chrysanthemum. Plant Quar. 2001, 06, 340–341. [Google Scholar]
- Supakitthanakorn, S.; Vichittragoontavorn, K.; Sunpapao, A.; Kunasakdakul, K.; Thapanapongworakul, P.; Ruangwong, O.U. Tobacco Mosaic Virus Infection of Chrysanthemums in Thailand: Development of Colorimetric Reverse-Transcription Loop-Mediated Isothermal Amplification (RT–LAMP) Technique for Sensitive and Rapid Detection. Plants 2022, 11, 1788. [Google Scholar] [CrossRef]
- Zhang, J.X.; Cao, X.F.; Song, T.; Ji, Y. Research progress of plant virus detection technology. Anal. Flower Towels Herbs 2011, 10, 79–80. [Google Scholar]
- Curry, A.; Appleton, H.; Barry, D. Application of transmission electron microscopy to the clinicalstudy of viral and bacterial infections: Present and future. Micron 2006, 37, 91–1061. [Google Scholar] [CrossRef] [PubMed]
- Kong, B.H.; Li, D.B. Infection of tomato infertility virus in chrysanthemum. J. Zhejiang Agric. Univ. 1990, 6, 55–60. [Google Scholar]
- Shu, X.Z.; Zhu, S.F.; Hu, W.Z.; Lu, J.Y.; Shen, S.L.; Wang, S.Q.; Chen, Y.F. Identification of tomato sterility virus infecting chrysanthemum and hyacinth. Chin. J. Virol. 1990, 2, 186–192. [Google Scholar]
- Wu, H.Y.; Kong, B.H.; Chen, H.R.; Liu, J.H. Investigation and identification of chrysanthemum virus disease in Kunming area. J. Yunnan Agric. Anthropol. 2002, 17, 24–27. [Google Scholar]
- Du, L.; Zhong, X.F.; Chen, J.H.; Yi, J.P. Research progress of plant virus detection technology. Agric. Prod. Process. Ind. (J.) 2007, 7, 48–51. [Google Scholar]
- Zhang, H.B.; Zhu, X.R.; Zhang, Y.K. Preliminary identification and serological detection of cucumber Mosaic virus infecting Chrysanthemum. Chin. J. Virol. 1995, 10, 367–370. [Google Scholar]
- Chen, J.Y.; Yang, H.; Fan, G.C.; Chen, J.Y. Preliminary serological detection and virus-free technology of Luo Yuan and Xiamen Cut Flower Chrysanthemum. Fujian J. Agric. Sci. 1998, 4, 13–15. [Google Scholar]
- Meng, R.X. Research progress of loop-mediated isothermal amplification technique in plant virus detection. Agric. Technol. Serv. 2022, 39, 72–77. [Google Scholar]
- Li, W.C.; Zhou, C.Y. Application of immunoelectron microscopy in the study of plant viruses. South. Agric. 2011, 05, 84–86. [Google Scholar]
- Guo, Q.N.; Zhang, W.F.; Jia, X.; Yin, Y.; Yan, W.Y.; Cui, Z.H.; Zhao, X.T. Research progress in elimination and detection of Chrysanthemum virus. J. Zhejiang Agric. Sci. 2016, 57, 2035–2039. [Google Scholar]
- Zhou, Y.Y. Study on Virus-Free Rapid Propagation Technology and Adaptability of Virus-Free Seedling of ‘Huai’ Chrysanthemum. Master’s Thesis, Henan Normal University, Xinxiang, China, 2014. [Google Scholar]
- Wu, H.Z.; Chen, H.R. Detection of Chrysanthemum B virus by RT-PCR. J. Southwest Agric. Univ. 2002, 24, 115–117. [Google Scholar]
- Tao, Y.; Wu, X.Q. Research progress of plant virus detection methods. Mol. Plant Breed. 2017, 15, 2901–2906. [Google Scholar]
- Liu, Z.B.; Gao, Q.R.; Wang, R.X. Application of multiplex PCR technique in plant biology research. Mol. Plant Breed. 2005, 3, 261–268. [Google Scholar]
- Song, A.; You, Y.; Chen, F.; Li, P.; Jiang, J.; Chen, S. A multiplex RT-PCR for rapid and simultaneous detection of viruses and viroids in chrysanthemum. Lett. Appl. Microbiol. 2013, 56, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L. Identification and Molecular Diagnostic Methods of Chrysanthemum Virus and Viroid Pathogens. Ph.D. Thesis, China Agricultural University, Beijing, China, 2014. [Google Scholar]
- Jin, Y.; Wen, J.Z. Research progress of plant virus detection methods. Heilongjiang Agric. Sci. 2005, 3, 37–40. [Google Scholar]
- Gu, Q.L.; Liu, H.; Zhang, S.H. Application of PCR related technology in plant virus quarantine detection. Biotechnol. Bull. 2011, 3, 78–81. [Google Scholar]
- Guan, Z.; Wu, D.; Song, A.; Chen, F.; Chen, S.; Fang, W. A highly sensitive method for the detection of Chrysanthemum virus B. Electron. J. Biotechnol. 2017, 26, 64–68. [Google Scholar] [CrossRef]
- Yan, C.N. Detection of CVB, TAV and CChMVd Viruses from Yan Chenna, Chrysanthemum and Establishment of Regeneration System of Leaf Disk of Chrysanthemum. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2008. [Google Scholar]
- Wu, D. Analysis of Active Components in ‘Chuzhou’ Chrysanthemum Virus Elimination and Virus-Free Seedling. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2016. [Google Scholar]
- Park, J.; Jung, Y.; Kil, E.-J.; Kim, J.; Tran, D.T.; Choi, S.-K.; Yoon, J.-Y.; Cho, W.K.; Lee, S. Loop-mediated isothermal amplification for the rapid detection of Chrysanthemum chlorotic mottle viroid (CChMVd). J. Virol. Methods 2013, 193, 232–237. [Google Scholar] [CrossRef]
- Liu, J.; Huang, C.L.; Wu, Z.Y.; Zhang, X.H.; Wang, Y.Q. Detection of tomato infertility virus in chrysanthemum by ring mediated isothermal amplification. Sci. Agric. Sin. 2010, 43, 1288–1294. [Google Scholar]
- Fukuta, S.; Nimi, Y.; Ohishi, K.; Yoshimura, Y.; Anai, N.; Hotta, M.; Fukaya, M.; Kato, T.; Oya, T.; Kanbe, M. Development of reverse transcription loop-mediated isothermal amplification (RT-LAMP) method for detection of two vinuses and Chrysanthemum stuntviroid. Annu. Rep. Kansai Plant Prot. Soc. 2005, 47, 31–36. [Google Scholar] [CrossRef]
- Hansen, A.J.; Hildebrandt, A.C. The distribution of Tobacco mosaic virus in plant callus cultures. Virology 1996, 28, 15. [Google Scholar] [CrossRef]
- Chinestra, S.C.; Curvetto, N.R.; Marinangeli, P.A. Production of virus-free plants of Lilium spp. from bulbs obtained in vitro and ex vitro. Sci. Hort. 2015, 194, 304–312. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, C.Q. Research progress of plant virus-free detection. J. Shandong Agric. Univ. (Nat. Sci. Ed.) 2003, 2, 209–212. [Google Scholar]
- Ratcliff, F.; Harrison, B.D.; Baulcombe, D.C. A similarity between viral defense and gene silencing in plants. Science 1997, 276, 1558. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.R.; Mao, B.Z. Research progress in detection and detoxification of plant viruses. Bull. Biotechnol. 2017, 33, 26–33. [Google Scholar]
- Jeon, S.M.; Savitri, W.D.; Park, K.I.; Chung, M.Y.; Kim, C.K. Elimination of Chrysanthemum stunt viroid (CSVd) from an Viroid-infected Chrysanthemum Through Shoot Tip Culture. Flower Res. 2012, 20, 218–222. [Google Scholar]
- Savitri, W.D.; Park, K.I.; Jeon, S.M.; Chung, M.Y.; Han, J.S.; Kim, C.K. Elimination of Chrysanthemum stunt viroid (CSVd) from Meristem Tip Culture Combined with Prolonged Cold Treatment. Hortic. Environ. Biotechnol. 2013, 54, 177–182. [Google Scholar] [CrossRef]
- Ram, R.; Verma, N.; Singh, A.K.; Singh, L.; Hallan, V.; Zaidi, A.A. Indexing and production of virus-free chrysanthemums. Biol. Plant. 2005, 49, 149–152. [Google Scholar] [CrossRef]
- You, Y.P.; Peng, S.Y.; Zhu, W.Y.; Wang, J.; Jiang, J.F.; Chen, S.M.; Fang, W.M.; Guan, Z.Y.; Chen, F.D. Elimination of Chrysanthemum B virus by shoot tip culture. J. Nanjing Agric. Univ. 2013, 36, 144–148. [Google Scholar]
- Hosokawa, M.; Otake, A.; Ohishi, K.; Ueda, E.; Hayashi, T.; Yazawa, S. Elimination of Chrysanthemum stunt viroid from an infected chrysanthemum cultivar by shoot regeneration from a leaf primordium-free shootapical meristem dome attached to a root tip. Plant Cell Rep. 2004, 22, 859–863. [Google Scholar] [CrossRef]
- Cai, H.; Kong, B.H.; Chen, H.R. Detection and identification of Phytophthora xanthogens (PY) from Changchun Flowers. Acta Microbiol. Sin. 2003, 43, 116–119. [Google Scholar]
- Zhao, S.; Li, Q.; Dai, S.L. Effects of different methods on the elimination of three viruses from Chrysanthemum. J. Northwest A F Univ. (Nat. Sci. Ed.) 2013, 41, 168–174. [Google Scholar]
- Ram, R.; Verma, N.; Singh, A.K.; Zaidi, A.A. Virus-free chrysanthemums: Production and quality management. Arch. Phytopathol. Plant Prot. 2009, 42, 940–949. [Google Scholar] [CrossRef]
- Yang, P.H.; Wang, R.R.; Liu, J.; Li, M.F. Research progress on virus-free and harmless technology of Chrysanthemum. Shaanxi Agric. Sci. 2011, 57, 89–92. [Google Scholar]
- Budiarto, K.; Marwoto, B.; Sanjaya, L.; Soedarjo, M.; Rahardjo, I.B. Elimination of CVB from a range of Chrysanthemum varieties by apical meristem culture following antiviral agent andheat treatments. Budiarto 2011, 18, 94–101. [Google Scholar]
- Wang, Q.C.; Panis, B.; Engelmann, F.; Lambardi, M.; Valkonen, J.P.T. Cryotherapy of shoot tips: A technique for pathogen eradication to produce healthy planting materials and prepare healthy plant genetic resources for cryopreservation. Ann. Appl. Biol. 2009, 154, 351–363. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, C.L.; Xu, M.L. Research progress of plant virus eradication using super low temperature processing. North. Hortic. 2013, 12, 184–187. [Google Scholar]
- Jeon, S.M.; Naing, A.H.; Kim, H.H.; Chung, M.Y.; Lim, K.B.; Kim, C.K. Elimination of Chrysanthemum stunt viroid and Chrysanthemum chlorotic mottle viroid from infected chrysanthemum by cryopreservation. Protoplasma 2016, 253, 1135–1144. [Google Scholar] [CrossRef]
- Verma, N.; Ram, R.; Hallan, V.A.; Kumar, K.; Zaidi, A.A. Production of Cucumber mosaic virus-free chrysanthemums bymeristem tip culture. Crop Prot. 2004, 23, 469–473. [Google Scholar] [CrossRef]
- Gong, J.G.; Yang, X.; Li, T.; Niu, Y.; Fan, J.Q. Tissue culture and propagation of ‘Chuzhou’ Chrysanthemum by leaf regeneration. J. Anhui Agric. Sci. 2013, 415, 7788–7789. [Google Scholar]
- Zhou, Y.; Wang, Y.; Song, Y.; Gao, Z.; Liu, Y.; Fan, L.; Hu, Q.; Gao, S. Stem Apex Detoxification Culture Markedly Improved Several Physiological Characters of Chrysanthemum ‘Yutai’. Plant Cell Tissue Organ Cult. 2014, 119, 369–381. [Google Scholar] [CrossRef]
- Chen, Q.; Xiao, J.M.; Sun, J. Research on production technology of virus-free Boju seedlings. Shanghai Agric. Sci. Technol. 2015, 2, 34–48. [Google Scholar]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ding, X.X. Research progress of tissue culture regeneration system of chrysanthemum. Liaoning For. Sci. Technol. 2018, 4, 64–65. [Google Scholar]
- Jahan, M.T.; Islam, M.R.; Islam, S.S.; Das, P.; Islam, M.M.; Kabir, M.H.; Mamun, A.N.K. Clonal Propagation of Chrysanthemum morifolium Ramat Using Various Explants Obtained from Field Grown Plants. GSC Biol. Pharm. Sci. 2021, 16, 87–93. [Google Scholar] [CrossRef]
- Deng, Y.M.; Chu, F.L.; Hua, R.Z. Study on virus-free technology of ‘Chuzhou’ Chrysanthemum tissue Culture. J. Anhui Agric. Sci. 2007, 14, 4130. [Google Scholar]
- Wang, K.C.; Zhang, X.Q.; Mao, Y.Y. Tissue culture of chrysanthemum petals. Chin. Herb. Med. 2000, 8, 70–72. [Google Scholar]
- Deng, N.F.; Wu, G.R. Study on Tissue culture and rapid propagation of Chrysanthemum petals. J. Hezhou Univ. 2007, 3, 144–145. [Google Scholar]
- Liu, Y.T.; Zhao, X.Z.; Yang, Y.X.; Yue, D.X.; Wei, W.C. Effects of different hormone concentrations on tissue culture of Edible Chrysanthemum. Acta Agric. Sin. 2014, 5, 67–69. [Google Scholar]
- Liu, Z.R. Tissue culture and rapid propagation of chrysanthemum. Chin. Flowers Bonsai 2003, 8, 36–37. [Google Scholar]
- Hou, Y.J.; Zhu, T.; Wang, X.T. Study on tissue culture of Chrysanthemum. J. Xinyang Norm. Univ. Nat. Sci. Ed. 2005, 3, 323–325. [Google Scholar]
- Fu, Y.L.; Wang, Y.Z.; Guo, Q.H.; Jia, H.Y. Tissue culture and rapid propagation of Compositum cerebrum. J. Anhui Agric. Sci. 2010, 24, 13367–13368. [Google Scholar]
- Huang, D.F.; Wang, J.X.; Mao, Y. Experiments on tissue culture and rapid propagation of a new variety of Cut Chrysanthemum. Shanghai Agric. Sci. Technol. 1992, 2, 38–39. [Google Scholar]
- Yuan, C.Z.; Li, B.; Yang, W.R.; Zhu, C.M. Effects of hormone on callus induction and cluster bud differentiation of Chrysanthemum. North. Hortic. 2010, 1, 162–164. [Google Scholar]
- Liu, P.; Liu, J.; Zhao, Y.H.; Liu, Q.X.; Zhang, W.G. Study on tissue culture, virus-free and rapid propagation of Chrysanthemum. J. Inn. Mong. Univ. Natl. Nat. Sci. Ed. 2005, 4, 410–413. [Google Scholar]
- Zhao, W.P.; Wang, Y.Q.; Liu, Y.; Huang, B.B.; Cai, H. Research progress of tissue culture technology of chrysanthemum. Anhui Agric. Sci. Bull. 2019, 25, 36–38. [Google Scholar]
- Jiang, X.W.; Liu, G.F.; Bao, M.Z. Establishment of rapid and efficient regeneration system for leaves and stem segments of 9 cultivars of Chrysanthemum. J. Huazhong Agric. Univ. 2003, 2, 162–166. [Google Scholar]
- Wang, F. Research on Resources, Seedling Breeding and Quality Analysis of Chrysanthemum. Master’s Thesis, Anhui Agricultural University, Hefei, China, 2013. [Google Scholar]
- Chen, X.L.; Chen, X.B.; Chen, Q.L. Application of tissue culture and rapid propagation technology of chrysanthemum in practical production. Fujian Hot Crop Sci. Technol. 2009, 4, 28–29. [Google Scholar]
- Wu, D.; Song, A.P.; Shi, Y.D.; Chen, F.D.; Fang, W.M.; Chen, S.M.; Guan, Z.Y. Virus removal and quality analysis of virus-free ‘Chuzhou’ Chrysanthemum seedlings. J. Nanjing Agric. Univ. 2017, 40, 983–992. [Google Scholar]
- Luo, H.; Wen, X.H.; Zhou, Y.Y.; Dai, S.L. Establishment of in vitro Regeneration System of Aromatic Coreopsis chinensis. Chin. Bull. Bot. 2020, 55, 318–328. [Google Scholar]
- Wang, S.; Chen, D.L.; Huang, J.L.; Pan, X.F.; Li, Y.H. Research progress of PCR-based detection of plant viruses. Plant Quar. 2021, 35, 1–8. [Google Scholar]
- Pei, Y.N.; Xian, S.; Qi, Y.H.; Zhang, L.; Wang, D.F.; Niu, Y.B. Sequencing and analysis of the complete genome of Chrysanthemum R virus isolated from Chrysanthemum. Chin. J. Biochem. Mol. Biol. 2020, 36, 217–224. [Google Scholar]
- Niu, E.B.; Chen, L.J.; Niu, Y.B. First Report of Zucchini yellow mosaic virus in Chrysanthemum. Plant Dis. 2015, 99, 1289. [Google Scholar] [CrossRef]
- Duarte, L.M.L.; Rivas, E.B.; Alexandre, M.A.V.; Ávila, A.C.; Nagata, T.; Chagas, C.M. Chrysanthemum Stem Necrosis Caused by a Possible Novel Tospovirus. J. Phytopathol. 1995, 143, 569–571. [Google Scholar] [CrossRef]
- Jonghe, K.; Morio, S.; Maes, M. First outbreak of Chrysanthemum stem necrosis virus (CSNV) on potted Chrysanthemum in Belgium. New Dis. Rep. 2013, 28, 14. [Google Scholar] [CrossRef]
- Matsuura, S.; Kubota, K.; Okuda, M. First Report of Chrysanthemum stem necrosis virus on Chrysanthemums in Japan. Plant Dis. 2007, 91, 468. [Google Scholar] [CrossRef] [PubMed]
- Hollings, M.; Stone, O.M. Attempts to eliminate chrysanthemum stunt from chrysanthemum by meristem-tip culture after heat-treatment. Ann. Appl. Biol. 1970, 65, 311–315. [Google Scholar] [CrossRef]
- Ebata, M.; Matsushita, Y.; Morimoto, M.; Mochizuki, T. Distribution of chrysanthemum chlorotic mottle viroid in shoot meristem and flower buds of chrysanthemum. Eur. J. Plant Pathol. 2019, 154, 555–563. [Google Scholar] [CrossRef]
- Mehle, N.; Dobnik, D.; Ravnikar, M.; Pompe Novak, M. Validated reverse transcription droplet digital PCR serves as a higher order method for absolute quantification of Potato virus Y strains. Anal. Bioanal. Chem. 2018, 410, 3815–3825. [Google Scholar] [CrossRef]
- Pandey, B.; Mallik, I.; Gudmestad, N.C. Development and application of a real-time reverse-transcription PCR and droplet digital PCR assays for the direct detection of Potato mop top virus in soil. Phytopathology 2020, 110, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Leichtfried, T.; Reisenzein, H.; Steinkellner, S.; Gottsberger, R.A. Transmission studies of the newly described apple chlorotic fruit spot viroid using a combined RT-qPCR and droplet digital PCR approach. Arch. Virol. 2020, 165, 2665–2671. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Y. Detection of GI and GII Norovirus in Fruits and Vegetables by Reverse Transcription Microdrop Digital PCR. Master’s Thesis, Jinan University, Guangzhou, China, 2018. [Google Scholar]
- Chu, Y.X.; Zhu, W.M. Advances in tomato breeding for resistance to yellow leaf curl virus. Genom. Appl. Biol. 2009, 28, 563–568. [Google Scholar]
- Zhang, J.X. Screening of Drought Resistant Germplasm and Discovery of Key Drought Resistant Genes of Generalized Chrysanthemum. Master’s Thesis, Beijing Forestry University, Beijing, China, 2020. [Google Scholar]
- Xu, T.T.; Ma, J.; Chi, T.H.; Ye, D.; Guan, Z.Y.; Fang, W.M.; Chen, F.D.; Zhang, F. Evaluation of cold resistance of cut flower chrysanthemum at seedling stage and related molecular marker mining. J. Nucl. Agric. 2019, 33, 199–207. [Google Scholar]
- Fu, X. Correlation Analysis and QTL Mapping of Molecular Markers of Aphid Resistance in Cut Flower Chrysanthemum. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2020. [Google Scholar]
- Ogawa, T.; Toguri, T.; Kudoh, H.; Okamura, M.; Momma, T.; Yoshioka, M.; Kato, K.; Hagiwara, Y.; Sano, T. Double-stranded RNA-specific ribonuclease confers tolerance against Chrysanthemum stunt viroid and Tomato spotted wilt virus in transgenic chrysanthemum Plants. Breed. Sci. 2005, 55, 49–55. [Google Scholar] [CrossRef]
- Tran, D.T.; Cho, S.; Hoang, P.M.; Kim, J.; Kil, E.J.; Lee, T.K.; Rhee, Y.; Lee, S. A Codon-optimized nucleic acid hydrolyzing single-chain antibody confers resistance to Chrysanthemums against Chrysanthemum stunt viroid infection. Plant Mol. Biol. Rep. 2016, 34, 221–232. [Google Scholar] [CrossRef]
- Torata, Y.; Asano, S.; Naka, K.; Yoshida, K.; Inda, K.; Matsushita, Y. Breeding of small-flowered chrysanthemum parental line with resistance to Chrysanthemum stunt viroid. Hortic. Res. 2018, 17, 524. [Google Scholar]
- Naka, T.; Asano, S.; Torata, Y.; Tsujimoto, N.; Matsushita, Y. Inheritance of resistance to CSVd in double flowered Chrysanthemum cultivars. Hortic. Res. 2016, 15, 444. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
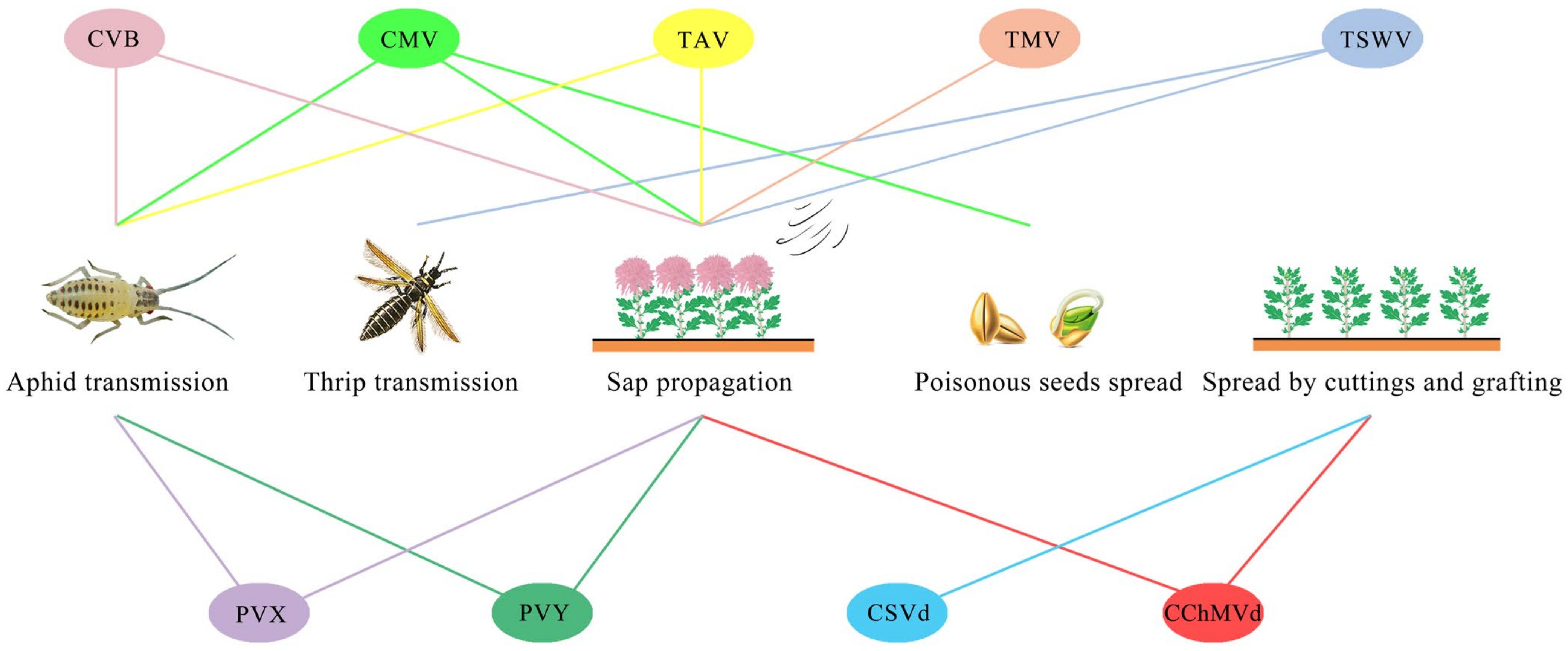
| Virus Name | Genus | Mode of Transmission | Range of Infection | References |
|---|---|---|---|---|
| CVB | Carlavirus | Aphid and sap | Compositae | [54,55,56] |
| CMV | Cucumovirus | Aphid, sap and poisonous seed | Compositae and Legume | [57] |
| TAV | Cucumovirus | Aphid and sap | Compositae, Solanaceae and Chenopodiaceae | [36,58] |
| TMV | Tobamovirus | Sap | Compositae and Solanaceae | [59] |
| PVX | Potexvirus | Aphid and sap | Compositae, Solanaceae, and Legumes | [60] |
| PVY | Potyvirus | Aphid and sap | Compositae, Solanaceae, Legumes, and Quinoa | [61,62] |
| TSWV | Orthotospovirus | Sap and thrips | Compositae, Solanaceae, Legumes, and Cucurbitaceae | [47,63,64] |
| CSVd | Pospiviroid | Cuttage and grafting | Compositae, Solanaceae, and Aracanthaceae | [65] |
| CChMVd | Pelamoviroid | Cuttage, grafting and sap | Compositae | [66,67] |
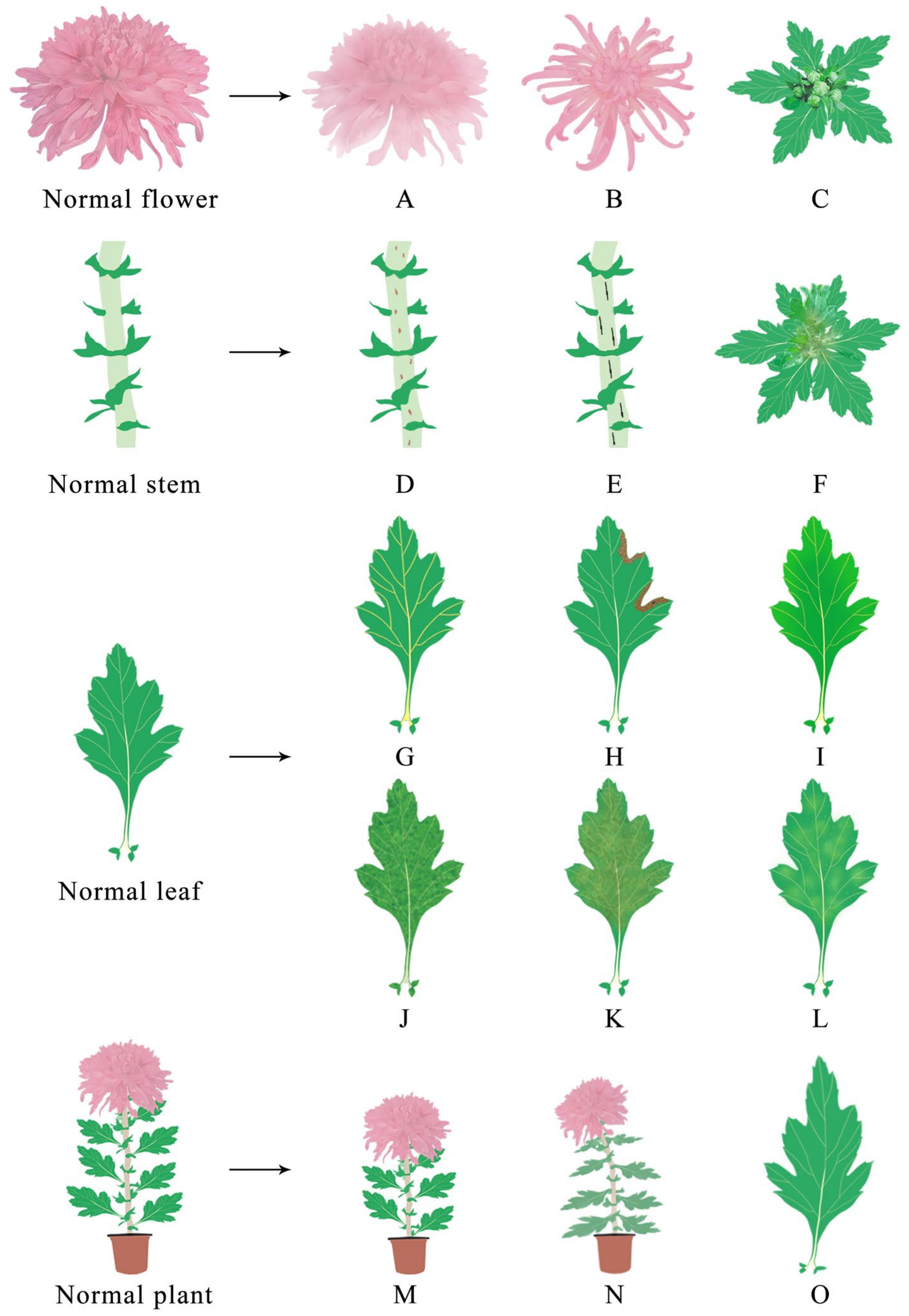
| Site of Infection | Virus Name | Symptom | References |
|---|---|---|---|
| Flower | CVB | Floral deformity | [68] |
| TSWV | No flowers and color changing | [69] | |
| CSVd | Color changing and early flowering | [8,9] | |
| Stem | TSWV | Brown streaks, black stripes and upper stem rot | [69] |
| Leaf | CMV | Vein uplift and mosaic | [59,70] |
| TSWV | Yellow brown ring spots | [69] | |
| CVB | Mosaic and necrosis spots | [68] | |
| TAV | Mosaic, deformation and necrosis spots | [71] | |
| TMV | Mosaic, necrosis spots, mottled, chlorisis and etiolation | [72] | |
| PVX | Mosaic, necrosis spots, mottled, chlorosis and etiolation | [73] | |
| PVY | Mottled, chlorisis and etiolation | [59] | |
| CSVd | Deformation and mottled | [5,6] | |
| CChMVd | Chlorosis and etiolation | [72,74] | |
| Plant | TAV | Dwarfism | [71] |
| CSVd | Dwarfism | [4] | |
| TSWV | Dwarfism and wilt | [69] |
| Detecting Techniques | Technological Type | The Species of Virus Detected | Detection Speed * | References |
|---|---|---|---|---|
| Indicator plant assay | —— | TAV, CMV, TMV, CVB | + | [84,85] |
| Electron microscope observation | Negative staining technology | TAV, CMV, TMV, CVB | ++ | [86,89,90,91] |
| ELISA | —— | TAV, CMV, TMV, PVX, PVY, CVB | ++ | [85,93,94] |
| Molecular Biology Technology | Single RT-PCR | TAV, TMV, CVB | +++ | [86,98,99] |
| Multiple RT-PCR | TAV, CVB, CMV, TMV, PVY, CChMVd, CSVd | ++++ | [102,103] | |
| Fluorescence quantitative RT-PCR | TAV, CVB, CMV, TMV, PVY, CChMVd, CSVd | +++ | [103] | |
| Nest-PCR | TAV, CVB, CMV, TMV, PVY | +++ | [107,108] | |
| RT-LAMP | TAV, TMV, CChMVd | +++ | [86,109,110] | |
| Multiple RT-LAMP | CVB, CMV, CSVd | ++++ | [103,111] |
| Virus Name | Elimination Method | Shoot Tip Size (mm) | Rate of Elimination (%) | Detection Techniques | References |
|---|---|---|---|---|---|
| CVB | Shoot tip | 0.3~1 | 0 | RT-PCR | [119] |
| Shoot tip | <0.2 | 43.6 | DAS-ELISA | [126] | |
| Shoot tip | 0.3~0.7 | 43.33 | RT-PCR | [120] | |
| Shoot tip with heat treatment | 0.3~1 | 20 | RT-PCR | [119] | |
| 36.3 | DAS-ELISA | ||||
| Shoot tip with heat treatment | 0.4~0.5 | 100 | ELISA | [123] | |
| Shoot tip, antiviral agents and heat treatment | <0.2 | 100 | DAS-ELISA | [126] | |
| Ultralow-temperature | 2.0 | 21.9 | RT-PCR | [12] | |
| 24.4 | DAS-ELISA | ||||
| Antiviral agents | 0.3~1 | 26.7 | RT-PCR | [119] | |
| 40 | DAS-ELISA | ||||
| CMV | Shoot tip | 0.3 | 72 | RT-PCR | [130] |
| 84 | DAS-ELISA | ||||
| Shoot tip | 0.3 | 65.6 | RT-PCR | [74] | |
| 78.1 | DAS-ELISA | ||||
| Shoot tip with heat treatment | 0.4~0.5 | 100 | ELISA | [84] | |
| TAV | Shoot tip | 0.3 | 65.6 | RT-PCR | [74] |
| 78.1 | DAS-ELISA | ||||
| TMV | Shoot tip, antiviral agents and heat treatment | 0.3~0.4 | 51.7 | RT-PCR | [124] |
| 65.9 | ELISA | ||||
| CSVd | Shoot tip | 0.3 | 14.3 | Nest PCR | [121] |
| Antiviral agents with 4 °C treatment | 30~50 | 100 | RT-PCR | [118] | |
| Ultralow-temperature | 30~50 | 20 | Nest PCR | [129] | |
| CChMVd | Ultralow-temperature | 30~50 | 20 | Nest PCR | [129] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, K.; Chen, Q.; Pan, B.; Sun, Y.; Xu, Y.; Chen, D.; Liu, H.; Luo, C.; Chen, X.; Li, H.; et al. Current Achievements and Future Prospects in Virus Elimination Technology for Functional Chrysanthemum. Viruses 2023, 15, 1770. https://doi.org/10.3390/v15081770
Gao K, Chen Q, Pan B, Sun Y, Xu Y, Chen D, Liu H, Luo C, Chen X, Li H, et al. Current Achievements and Future Prospects in Virus Elimination Technology for Functional Chrysanthemum. Viruses. 2023; 15(8):1770. https://doi.org/10.3390/v15081770
Chicago/Turabian StyleGao, Kang, Qingbing Chen, Bo Pan, Yahui Sun, Yuran Xu, Dongliang Chen, Hua Liu, Chang Luo, Xi Chen, Haiying Li, and et al. 2023. "Current Achievements and Future Prospects in Virus Elimination Technology for Functional Chrysanthemum" Viruses 15, no. 8: 1770. https://doi.org/10.3390/v15081770
APA StyleGao, K., Chen, Q., Pan, B., Sun, Y., Xu, Y., Chen, D., Liu, H., Luo, C., Chen, X., Li, H., & Huang, C. (2023). Current Achievements and Future Prospects in Virus Elimination Technology for Functional Chrysanthemum. Viruses, 15(8), 1770. https://doi.org/10.3390/v15081770