
Humoral Immune Responses after an Omicron-Adapted Booster BNT162b2 Vaccination in Patients with Lymphoid Malignancies

 , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Antibody Quantification
2.3. Measurement of Virus-Neutralizing Capacity
2.4. Statistics and Modeling
3. Results
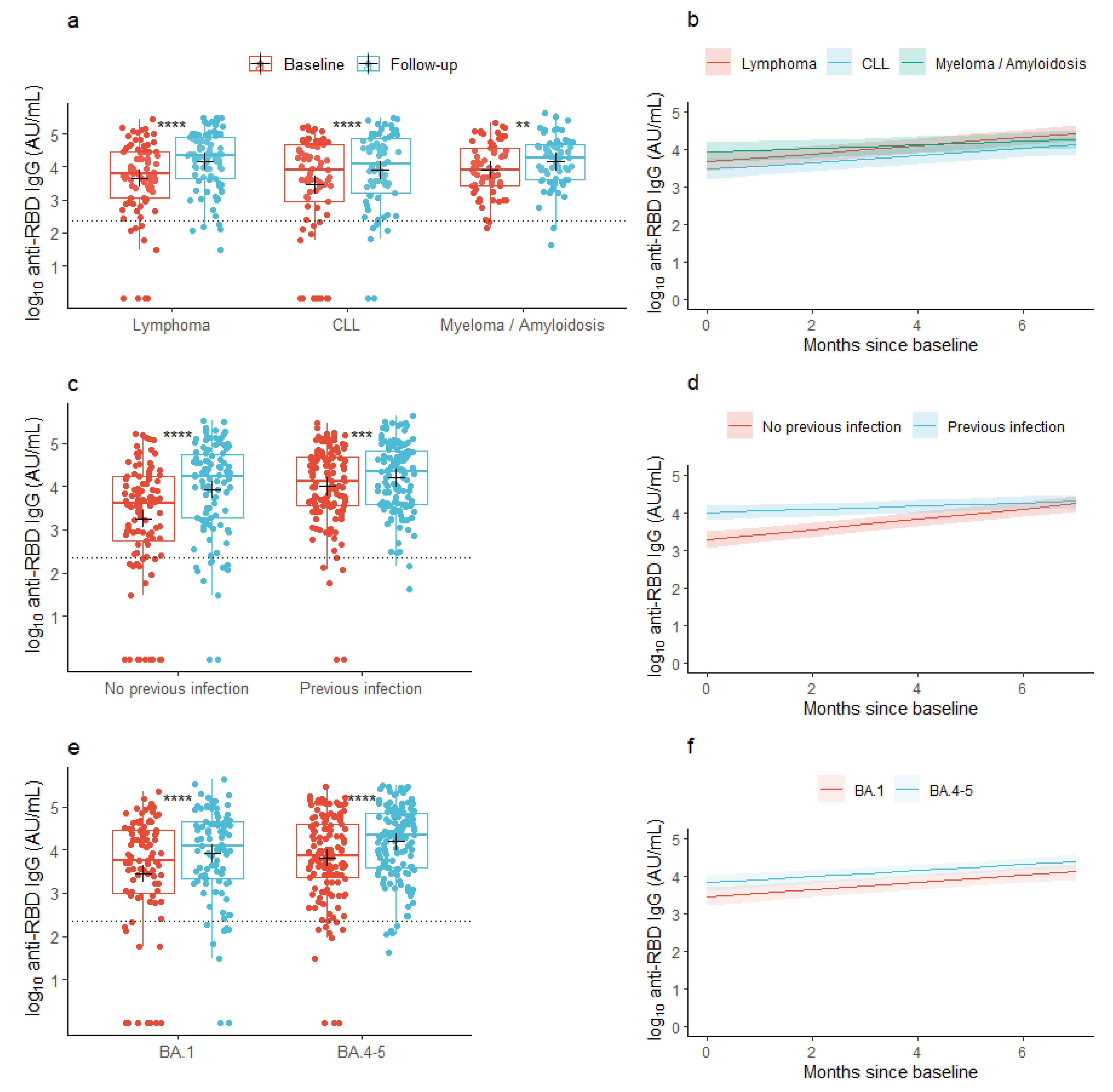
3.1. Anti-RBD IgG Concentrations
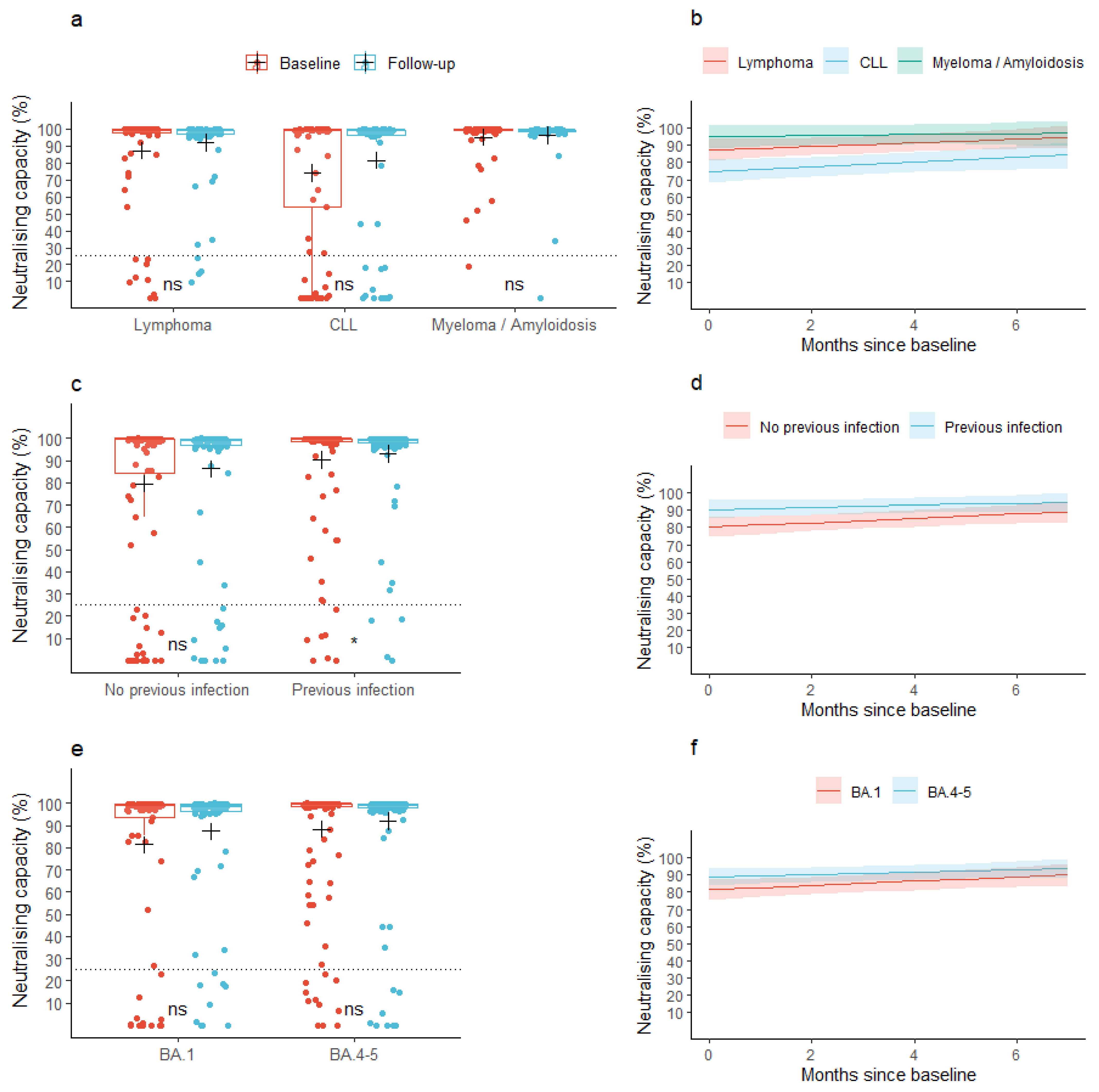
3.2. Neutralizing Capacity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Statement on the Fifteenth Meeting of the IHR (2005) Emergency Committee on the COVID-19 Pandemic. Available online: https://www.who.int/news/item/05-05-2023-statement-on-the-fifteenth-meeting-of-the-international-health-regulations-(2005)-emergency-committee-regarding-the-coronavirus-disease-(covid-19)-pandemic?adgroupsurvey=%7Badgroupsurvey%7Dgclid=Cj0KCQjwj_ajBhCqARIsAA37s0z3Y6LqRWqMyRh_AAF45q23xZdgQ1OUcWPkm-lUuN7Ic34MHYQGF5saAkU7EALw_wcB (accessed on 5 June 2023).
- Nab, L.; Parker, E.P.K.; Andrews, C.D.; Hulme, W.J.; Fisher, L.; Morley, J.; Mehrkar, A.; MacKenna, B.; Inglesby, P.; Morton, C.E.; et al. Changes in COVID-19-related mortality across key demographic and clinical subgroups in England from 2020 to 2022: A retrospective cohort study using the OpenSAFELY platform. Lancet Public Health 2023, 8, e364–e377. Available online: https://pubmed.ncbi.nlm.nih.gov/37120260/ (accessed on 26 June 2023). [CrossRef] [PubMed]
- Pagano, L.; Salmanton-García, J.; Marchesi, F.; López-García, A.; Lamure, S.; Itri, F.; Gomes-Silva, M.; Dragonetti, G.; Falces-Romero, I.; van Doesum, J.; et al. COVID-19 in vaccinated adult patients with hematological malignancies: Preliminary results from EPICOVIDEHA. Blood 2022, 139, 1588. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8577877/ (accessed on 27 June 2023). [CrossRef] [PubMed]
- Krekeler, C.; Reitnauer, L.; Bacher, U.; Khandanpour, C.; Steger, L.; Boeckel, G.R.; Klosner, J.; Tepasse, P.R.; Kemper, M.; Hennies, M.T.; et al. Efficacy of COVID-19 Booster Vaccines in Patients with Hematologic Malignancies: Experiences in a Real-World Scenario. Cancers 2022, 14, 5512. Available online: https://pubmed.ncbi.nlm.nih.gov/36428605/ (accessed on 27 June 2023). [CrossRef] [PubMed]
- Heftdal, L.D.; Hamm, S.R.; Pérez-Alós, L.; Madsen, J.R.; Armenteros, J.J.A.; Fogh, K.; Kronborg, C.C.; Vallentin, A.P.; Hasselbalch, R.B.; Møller, D.L.; et al. Humoral and cellular immune responses after three or four doses of BNT162b2 in patients with hematological malignancies. Eur. J. Haematol. 2023, 111, 229–239. Available online: https://pubmed.ncbi.nlm.nih.gov/37151174/ (accessed on 9 September 2023). [CrossRef] [PubMed]
- Menegale, F.; Manica, M.; Zardini, A.; Guzzetta, G.; Marziano, V.; d’Andrea, V.; Trentini, F.; Ajelli, M.; Poletti, P.; Merler, S. Evaluation of Waning of SARS-CoV-2 Vaccine–Induced Immunity: A Systematic Review and Meta-analysis. JAMA Net. Open 2023, 6, e2310650. Available online: https://jamanetwork.com/journals/jamanetworkopen/fullarticle/2804451 (accessed on 27 June 2023). [CrossRef] [PubMed]
- Kozlov, M. Should COVID vaccines be given yearly? Proposal divides US scientists. Nature 2023. ahead of print. Available online: https://pubmed.ncbi.nlm.nih.gov/36707704/ (accessed on 9 September 2023).
- Pérez-Alós, L.; Armenteros, J.J.A.; Madsen, J.R.; Hansen, C.B.; Jarlhelt, I.; Hamm, S.R.; Heftdal, L.D.; Pries-Heje, M.M.; Møller, D.L.; Fogh, K.; et al. Modeling of waning immunity after SARS-CoV-2 vaccination and influencing factors. Nat. Commun. 2022, 131, 1614. Available online: https://www.nature.com/articles/s41467-022-29225-4 (accessed on 7 July 2022). [CrossRef] [PubMed]
- Voldstedlund, M.; Haarh, M.; Mølbak, K. The danish microbiology database (MIBA) 2010 to 2013. Eurosurveillance 2014, 19, 20667. Available online: https://www.eurosurveillance.org/content/10.2807/1560-7917.ES2014.19.1.20667 (accessed on 21 June 2023). [CrossRef] [PubMed]
- Grove Krause, T.; Jakobsen, S.; Haarh, M.; Mølbak, K. The Danish vaccination register. Eurosurveillance 2012, 17, 2. Available online: https://www.eurosurveillance.org/content/10.2807/ese.17.17.20155-en (accessed on 6 September 2022). [CrossRef] [PubMed]
- Hansen, C.B.; Jarlhelt, I.; Hasselbalch, R.B.; Hamm, S.R.; Fogh, K.; Pries-Heje, M.M.; Møller, D.L.; Heftdal, L.D.; Pérez-Alós, L.; Sørensen, E.; et al. Antibody-dependent neutralizing capacity of the SARS-CoV-2 vaccine BNT162b2 with and without previous COVID-19 priming. J. Intern. Med. 2021, 290, 1272–1274. Available online: https://onlinelibrary.wiley.com/doi/10.1111/joim.13366 (accessed on 9 July 2021). [CrossRef] [PubMed]
- R Core Team, R A Language and Environment for Statistical Computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2019; Available online: https://www.scirp.org/(S(lz5mqp453edsnp55rrgjct55))/reference/ReferencesPapers.aspx?ReferenceID=2631126 (accessed on 14 December 2023).
- A Grammar of Data Manipulation [R Package Dplyr Version 1.0.7]. 2021. Available online: https://cran.r-project.org/package=dplyr (accessed on 4 November 2021).
- Tests in Linear Mixed Effects Models [R Package lmerTest Version 3.1-3]. 2020. Available online: https://cran.r-project.org/package=lmerTest (accessed on 22 June 2023).
- Xiao, N. Scientific Journal and Sci-Fi Themed Color Palettes for ‘ggplot2’ [R Package Ggsci Version 2.9]. 2018. Available online: https://cran.r-project.org/package=ggsci (accessed on 27 August 2022).
- Chemaitelly, H.; Ayoub, H.H.; Tang, P.; Coyle, P.; Yassine, H.M.; Al Thani, P.A.A.; Al-Khatib, H.A.; Hasan, M.R.; Al-Kanaani, Z.; Al-Kuwari, E.; et al. Long-term COVID-19 booster effectiveness by infection history and clinical vulnerability and immune imprinting: A retrospective population-based cohort study. Lancet Infect Dis. 2023, 23, 816–827. Available online: https://pubmed.ncbi.nlm.nih.gov/36913963/ (accessed on 22 April 2023). [CrossRef] [PubMed]
- King, S.M.; Bryan, S.P.; Hilchey, S.P.; Wang, J.; Zand, M.S. First Impressions Matter: Immune Imprinting and Antibody Cross-Reactivity in Influenza and SARS-CoV-2. Pathogens 2023, 12, 169. Available online: https://pubmed.ncbi.nlm.nih.gov/36839441/ (accessed on 22 April 2023). [CrossRef] [PubMed]
- Pérez-Alós, L.; Hansen, C.B.; Almagro Armenteros, J.J.; Madsen, J.R.; Heftdal, L.D.; Hasselbalch, R.B.; Pries-Heje, M.M.; Bayarri-Olmos, R.; Jarlhelt, I.; Hamm, S.R.; et al. Previous immunity shapes immune responses to SARS-CoV-2 booster vaccination and Omicron breakthrough infection risk. Nat. Commun 2023, 14, 5624. Available online: https://pubmed.ncbi.nlm.nih.gov/37699890/ (accessed on 2 October 2023). [CrossRef] [PubMed]
- Wang, W.; Goguet, E.; Padilla, S.P.; Vassell, R.; Pollett, S.; Mitre, E.; Weiss, C.D. The Journal of Infectious Diseases BRIEF REPORT Bivalent COVID-19 vaccine antibody responses to Omicron variants suggest that responses to divergent variants would be improved with matched vaccine antigens. J. Infect Dis. 2023, 228, 439–443. Available online: https://academic.oup.com/jid/advance-article/doi/10.1093/infdis/jiad111/7190664 (accessed on 19 June 2023). [CrossRef] [PubMed]
- Baerends, E.A.M.; Reekie, J.; Andreasen, S.R.; Stærke, N.B.; Raben, D.; Nielsen, H.; Petersen, K.T.; Johansen, I.S.; Lindvig, S.O.; Madsen, L.W.; et al. Omicron variant-specific serological imprinting following BA.1 or BA.4/5 bivalent vaccination and previous SARS-CoV-2 infection: A cohort study. Clin. Infect Dis. 2023, 77, 1511–1520. Available online: https://pubmed.ncbi.nlm.nih.gov/37392436/ (accessed on 6 July 2023). [CrossRef] [PubMed]
- Available online: https://www.sst.dk/-/media/Udgivelser/2022/Corona/Vaccination/Efteraar/Anbefalinger-vaccinationsprogram-efteraar-2022.ashx (accessed on 3 October 2023).



{kind=link}
{kind=link}
| All Patients n = 233 | Chronic Lymphatic B-Cell Leukemia n = 73 (31.3%) | Lymphoma n = 89 (38.2%) | Multiple Myeloma/Amyloidosis n = 71 (30.5%) | p-Value | ||
|---|---|---|---|---|---|---|
| Age; years, median [IQR] | 69 [63, 75] | 71 [66, 75] | 68 [62, 74] | 69 [62, 76] | 0.172 | |
| Male sex, n (%) | 116 (49.8) | 46 (63.0) | 40 (44.9) | 30 (42.3) | 0.023 | |
| BMI, mean (SD) | 25.4 (4.1) Missing: 5 | 25.3 (4.1) Missing: 2 | 25.6 (4) Missing: 2 | 25.4 (4.1) Missing: 1 | 0.739 | |
| No. of COVID-19 vaccines at follow-up, n (%) | 4 doses, n (%) | 38 (16.3) | 13 (17.8) | 21 (23.6) | 4 (5.6) | - |
| 5 doses, n (%) | 195 (83.7) | 60 (82.2) | 68 (76.4) | 67 (94.4) | 0.009 | |
| Vaccine variant | BA. 1 | 93 (39.9) | 28 (38.4) | 37 (41.6) | 28 (39.4) | - |
| BA. 4-5 | 140 (60.1) | 45 (61.6) | 52 (58.4) | 43 (60.6) | 0.913 | |
| Months between samples, median [IQR] | 4.6 [4.0, 5.1] | 4.4 [4.1, 5.0] | 4.5 [4.1, 5.1] | 4.6 [3.9, 5.1] | 0.820 | |
| Months from baseline to vaccination, median [IQR] | 0.8 [0.5, 1.2] | 0.7 [0.3, 1.2] | 0.8 [0.5, 1.2] | 0.9 [0.5, 1.2] | 0.513 | |
| Months from vaccination to follow-up, median [IQR] | 3.7 [3.2, 4.2] | 3.8 [3.3, 4.1] | 3.8 [3.2, 4.2] | 3.6 [3.0, 4.1] | 0.237 | |
| Days between doses, median [IQR] | 1st to 2nd dose | 23 [21, 24] | 23 [21, 25] | 23 [22, 24] | 23 [21, 24] | 0.372 |
| 2nd to 3rd dose | 183 [175, 203] | 179 [172, 202] | 184 [176, 212] | 184 [176, 201] | 0.150 | |
| 3rd to 4th dose | 141 [136, 179] | 142 [137, 179] | 141 [138, 285] | 140 [133, 147.5] | 0.012 | |
| 4th to 5th dose | 248 [238, 254.5] | 248 [242, 254.2] | 248 [240.5, 252] | 249 [237, 257] | 0.737 | |
| COVID-19 infection (PCR or N-Ab), n (%) | At baseline | 129 (55.4) | 40 (54.8) | 46 (51.7) | 43 (60.6) | 0.529 |
| At follow-up | 157 (67.4) | 49 (67.1) | 57 (64.0) | 51 (71.8) | 0.579 | |
| Treatment with anti-SARS-CoV-2 immunoglobulins | <1 year prior to baseline | 38 (16.3) | 16 (21.9) | 8 (9.0) | 14 (19.7) | 0.056 |
| By time of follow-up | 41 (17.6) | 17 (23.3) | 9 (10.1) | 15 (21.1) | 0.058 | |
| Treatment with human immunoglobulin | <1 year prior to baseline | 26 (11.2) | 12 (16.4) | 7 (7.9) | 7 (9.9) | 0.207 |
| By time of follow-up | 32 (13.7) | 12 (16.4) | 10 (11.2) | 10 (14.1) | 0.629 | |
| B-cell- or plasma-cell-targeted treatment and immunomodulatory treatment, n (% of patient population) <1 year prior to baseline | Anti-CD20 therapy | 9 (3.9) | 3 (4.1) | 6 (6.7) | - | 0.088 |
| Anti-CD30 therapy | 1 (0.4) | - | 1 (1.1) | - | 0.444 | |
| Anti-CD38 therapy | 25 (10.7) | - | - | 25 (35.2) | <0.001 | |
| Proteasome inhibitors | 7 (3.0) | - | - | 7 (9.9) | <0.001 | |
| Proteinkinase, PI3K, or BCL2 inhibitors | 24 (10.3) | 17 (23.3) | 6 (6.7) | 1 (1.4) | <0.001 | |
| Immunomodulatory imide drugs (IMiDs) | 25 (10.7) | - | 1 (1.1) | 24 (33.8) | <0.001 | |
| (a) |
SARS-CoV-2 anti-RBD IgG Partially Adjusted Model * |
SARS-CoV-2 anti-RBD IgG Fully Adjusted Model # | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Difference in Baseline log10 AU/mL (95% CI) | p-Value | Change in log10 AU/mL/Month (95% CI) | p-Value | Difference in Baseline log10 AU/mL (95% CI) | p-Value | Change in log10 AU/mL/Month (95% CI) | p-Value | ||
| Malignancy type | Lymphoma | Ref | Ref | Ref | Ref | Ref | Ref | Ref | Ref |
| Myeloma/Amyloidosis | 0.255 (−0.117, 0.627) | 0.181 | −0.059 (−0.109; −0.009) | 0.022 | 0.267 (−0.088; 0.623) | 0.147 | −0.058 (−0.108; −0.008) | 0.025 | |
| CLL | −0.198 (−0.567; 0.171) | 0.295 | −0.017 (−0.066; 0.033) | 0.515 | −0.159 (−0.513; 0.195) | 0.385 | −0.014 (−0.064; 0.036) | 0.573 | |
| Age, per 10 years | −0.209 (−0.378; −0.040) | 0.016 | 0.018 (−0.005; 0.041) | 0.120 | −0.067 (−0.203; 0.070) | 0.345 | - | - | |
| Sex | Female | Ref | Ref | Ref | Ref | Ref | Ref | - | - |
| Male | 0.057 (−0.253; 0.366) | 0.721 | −0.009 (−0.051; 0.033) | 0.680 | 0.067 (−0.173; 0.308) | 0.592 | - | - | |
| No. of COVID-19 vaccines at follow-up | 4 doses | Ref | Ref | Ref | Ref | Ref | Ref | - | - |
| 5 doses | −0.159 (−0.578; 0.259) | 0.456 | −0.027 (−0.085; 0.030) | 0.355 | −0.197 (−0.526; 0.132) | 0.249 | - | - | |
| Vaccine variant | BA.1 | Ref | Ref | Ref | Ref | Ref | Ref | - | - |
| BA.4-5 | 0.363 (0.051; 0.675) | 0.024 | −0.012 (−0.055; −0.030) | 0.566 | 0.252 (0.004; 0.500) | 0.052 | - | - | |
| COVID-19 infection (verified using PCR and/or N-Ab) < 1 year prior to baseline | 0.723 (0.426; 1.020) | <0.001 | −0.094 (−0.135; −0.054) | < 0.001 | 0.358 (0.103; 0.612) | 0.005 | - | - | |
| Treatment with human immunoglobulin < 1 year prior to baseline | −0.494 (−0.982; −0.007) | 0.048 | 0.016 (−0.052; 0.083) | 0.644 | −0.498 (−0.884; −0.111) | 0.013 | - | - | |
| (b) | Neutralizing Capacity Partially Adjusted Model * | Neutralizing Capacity Fully Adjusted Model # | |||||||
| Difference in Baseline% (95% CI) | p-Value | Change in pp/Month (95% CI) | p-Value | Difference in Baseline% (95% CI) | p-Value | Change in pp/Month (95% CI) | p-Value | ||
| Malignancy type | Lymphoma | Ref | Ref | Ref | Ref | Ref | Ref | Ref | Ref |
| Myeloma/Amyloidosis | 7.8 (−1.3; 16.9) | 0.094 | −0.8 (−2.5; 1.0) | 0.388 | −8.4 (−0.6; 17.4) | 0.073 | −0.8 (−2.5; 1.0) | 0.389 | |
| CLL | −12.4 (−21.4; −3.4) | 0.008 | 0.3 (−1.4; 2.0) | 0.751 | −12.0 (−21.0; −3.0) | 0.010 | 0.3 (−1.4; 2.0) | 0.708 | |
| Age, per 10 years | −4.2 (−8.4; 0.1) | 0.057 | 0.2 (−0.5; 1.0) | 0.540 | −1.5 (−4.9; 1.9) | 0.403 | - | - | |
| Sex | Female | Ref | Ref | Ref | Ref | Ref | Ref | - | - |
| Male | −1.5 (−9.2; 6.3) | 0.715 | −0.5 (−1.9; 1.0 | 0.524 | −0.5 (−6.5; 5.5) | 0.874 | - | - | |
| No. of COVID-19 vaccines at follow-up | 4 doses | Ref | Ref | Ref | Ref | Ref | Ref | - | - |
| 5 doses | −7.5 (−18.0; 2.9) | 0.160 | 0.6 (−1.4; 2.6) | 0.546 | −6.5 (−14.7; 1.8) | 0.130 | - | - | |
| Vaccine variant | BA.1 | Ref | Ref | Ref | Ref | Ref | Ref | - | - |
| BA.4-5 | 7.1 (−0.7; 15.0) | 0.078 | −0.5 (−1.9; 1.0) | 0.512 | 4.2 (−2.0; 10.5) | 0.190 | - | - | |
| COVID-19 infection (verified using PCR and/or N-Ab) < 1 year prior to baseline | 10.3 (2.6; 18.0) | 0.010 | −0.7 (−2.1; 0.8) | 0.357 | 6.7 (0.6; 12.8) | 0.035 | - | - | |
| Treatment with human immunoglobulin < 1 year prior to baseline | −5.5 (−17.8; 6.8) | 0.385 | 1.9 (−0.4; 4.1) | 0.108 | 0.3 (−9.4; 10.0) | 0.954 | - | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heftdal, L.D.; Hansen, C.B.; Hamm, S.R.; Pérez-Alós, L.; Fogh, K.; Pries-Heje, M.; Hasselbalch, R.B.; Møller, D.L.; Gang, A.O.; Ostrowski, S.R.; et al. Humoral Immune Responses after an Omicron-Adapted Booster BNT162b2 Vaccination in Patients with Lymphoid Malignancies. Viruses 2024, 16, 11. https://doi.org/10.3390/v16010011
Heftdal LD, Hansen CB, Hamm SR, Pérez-Alós L, Fogh K, Pries-Heje M, Hasselbalch RB, Møller DL, Gang AO, Ostrowski SR, et al. Humoral Immune Responses after an Omicron-Adapted Booster BNT162b2 Vaccination in Patients with Lymphoid Malignancies. Viruses. 2024; 16(1):11. https://doi.org/10.3390/v16010011
Chicago/Turabian StyleHeftdal, Line Dam, Cecilie Bo Hansen, Sebastian Rask Hamm, Laura Pérez-Alós, Kamille Fogh, Mia Pries-Heje, Rasmus Bo Hasselbalch, Dina Leth Møller, Anne Ortved Gang, Sisse Rye Ostrowski, and et al. 2024. "Humoral Immune Responses after an Omicron-Adapted Booster BNT162b2 Vaccination in Patients with Lymphoid Malignancies" Viruses 16, no. 1: 11. https://doi.org/10.3390/v16010011
APA StyleHeftdal, L. D., Hansen, C. B., Hamm, S. R., Pérez-Alós, L., Fogh, K., Pries-Heje, M., Hasselbalch, R. B., Møller, D. L., Gang, A. O., Ostrowski, S. R., Frikke-Schmidt, R., Sørensen, E., Hilsted, L., Bundgaard, H., Garred, P., Iversen, K., Sabin, C., Nielsen, S. D., & Grønbæk, K. (2024). Humoral Immune Responses after an Omicron-Adapted Booster BNT162b2 Vaccination in Patients with Lymphoid Malignancies. Viruses, 16(1), 11. https://doi.org/10.3390/v16010011