
Retreatment with HBV siRNA Results in Additional Reduction in HBV Antigenemia and Immune Stimulation in the AAV-HBV Mouse Model

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement and Animal Experimentation
2.2. Compounds
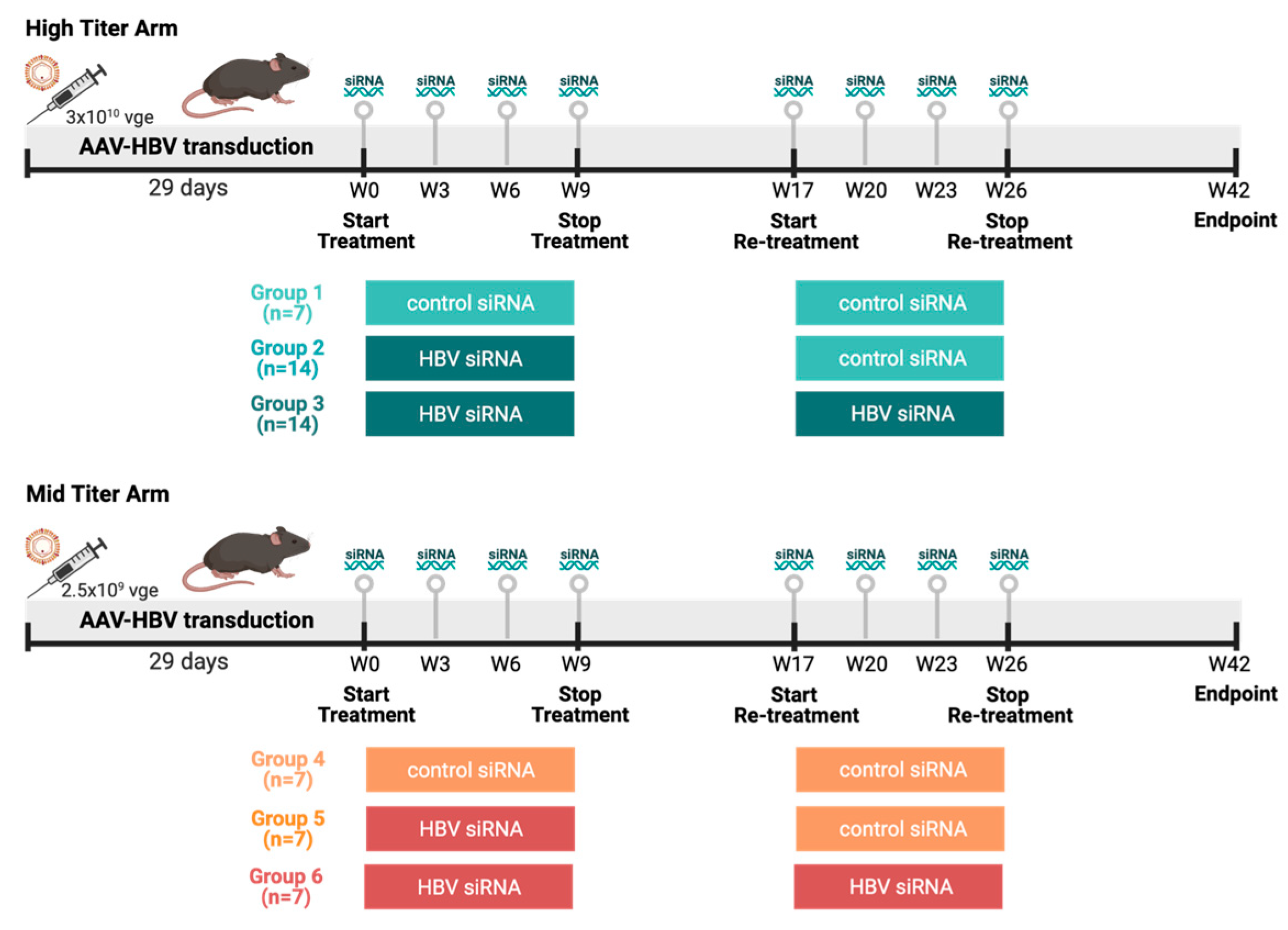
2.3. Animal Models and Study Designs
2.4. Viral Parameters and Alanine Aminotransferase (ALT) Analyses
2.5. Histology and Immunohistochemistry (IHC)
2.6. Image Analysis
2.7. Isolation Intrahepatic Lymphocytes (IHLs)
2.8. Bulk Liver RNA-Sequencing and Data Analysis
2.9. Flow Cytometry
2.10. Statistical Analysis
3. Results
3.1. Two Cycles (But Not One) of HBV-siRNA Treatment Achieve Undetectable HbsAg and HbeAg Levels in a Baseline HbsAg Level Dependent Manner
3.2. Bulk RNA-Sequencing Shows Modest Changes in the Liver after siRNA Treatment
3.3. HBV-siRNA Retreatment Enhances Expression of Checkpoint Receptors and Upregulates CD4 Regulatory T-Cells (Treg) Compared to Control Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. W.H.O. Hepatitis B. 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-b (accessed on 12 March 2023).
- European Association for the Study of the Liver; Lampertico, P.; Agarwal, K.; Berg, T.; Buti, M.; Janssen, H.L.; Papatheodoridis, G.; Zoulim, F.; Tacke, F. EASL 2017 Clinical Practice Guidelines on the management of hepatitis B virus infection. J. Hepatol. 2017, 67, 370–398. [Google Scholar] [CrossRef] [PubMed]
- Terrault, N.A.; Lok, A.S.F.; McMahon, B.J.; Chang, K.-M.; Hwang, J.P.; Jonas, M.M.; Brown, R.S., Jr.; Bzowej, N.H.; Wong, J.B. Update on prevention, diagnosis, and treatment of chronic hepatitis B: AASLD 2018 hepatitis B guidance. Hepatology 2018, 67, 1560–1599. [Google Scholar] [CrossRef] [PubMed]
- Lok, A.S.; Zoulim, F.; Dusheiko, G.; Ghany, M.G. Hepatitis B cure: From discovery to regulatory approval. Hepatology 2017, 66, 1296–1313. [Google Scholar] [CrossRef] [PubMed]
- Yuen, M.; Wong, D.K.; Fung, J.; Ip, P.; But, D.; Hung, I.; Lau, K.; Yuen, J.C.; Lai, C. HBsAg Seroclearance in chronic hepatitis B in Asian patients: Replicative level and risk of hepatocellular carcinoma. Gastroenterology 2008, 135, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Mak, L.Y.; Seto, W.K.; Yuen, M.F. Novel Antivirals in Clinical Development for Chronic Hepatitis B Infection. Viruses 2021, 13, 1169. [Google Scholar] [CrossRef] [PubMed]
- Gane, E.; Lim, Y.-S.; Kim, J.B.; Jadhav, V.; Shen, L.; Bakardjiev, A.I.; Huang, S.A.; Cathcart, A.L.; Lempp, F.A.; Janas, M.M.; et al. Evaluation of RNAi therapeutics VIR-2218 and ALN-HBV for chronic hepatitis B: Results from randomized clinical trials. J. Hepatol. 2023, 79, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Yuen, M.-F.; Locarnini, S.; Lim, T.H.; Strasser, S.I.; Sievert, W.; Cheng, W.; Thompson, A.J.; Given, B.D.; Schluep, T.; Hamilton, J.; et al. Combination treatments including the small-interfering RNA JNJ-3989 induce rapid and sometimes prolonged viral responses in patients with CHB. J. Hepatol. 2022, 77, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Yuen, M.-F.; Lim, S.-G.; Plesniak, R.; Tsuji, K.; Janssen, H.L.; Pojoga, C.; Gadano, A.; Popescu, C.P.; Stepanova, T.; Asselah, T.; et al. Efficacy and Safety of Bepirovirsen in Chronic Hepatitis B Infection. N. Engl. J. Med. 2022, 387, 1957–1968. [Google Scholar] [CrossRef]
- Hui, R.W.-H.; Mak, L.-Y.; Seto, W.-K.; Yuen, M.-F. RNA interference as a novel treatment strategy for chronic hepatitis B infection. Clin. Mol. Hepatol. 2022, 28, 408–424. [Google Scholar] [CrossRef]
- Yeo, Y.H.; Ho, H.J.; Yang, H.I.; Tseng, T.C.; Hosaka, T.; Trinh, H.N.; Kwak, M.S.; Park, Y.M.; Fung, J.Y.; Buti, M.; et al. Factors Associated with Rates of HBsAg Seroclearance in Adults with Chronic HBV Infection: A Systematic Review and Meta-analysis. Gastroenterology 2019, 156, 635–646.e9. [Google Scholar] [CrossRef]
- Springer, A.D.; Dowdy, S.F. GalNAc-siRNA Conjugates: Leading the Way for Delivery of RNAi Therapeutics. Nucleic Acid. Ther. 2018, 28, 109–118. [Google Scholar] [CrossRef]
- Rensen, P.C.N.; van Leeuwen, S.H.; Sliedregt, L.A.J.M.; van Berkel, T.J.C.; Biessen, E.A.L. Design and synthesis of novel N-acetylgalactosamine-terminated glycolipids for targeting of lipoproteins to the hepatic asialoglycoprotein receptor. J. Med. Chem. 2004, 47, 5798–5808. [Google Scholar] [CrossRef]
- Konishi, M.; Wu, C.H.; Wu, G.Y. Inhibition of HBV replication by siRNA in a stable HBV-producing cell line. Hepatology 2003, 38, 842–850. [Google Scholar] [CrossRef]
- Fisicaro, P.; Barili, V.; Rossi, M.; Montali, I.; Vecchi, A.; Acerbi, G.; Laccabue, D.; Zecca, A.; Penna, A.; Missale, G.; et al. Pathogenetic Mechanisms of T Cell Dysfunction in Chronic HBV Infection and Related Therapeutic Approaches. Front. Immunol. 2020, 11, 849. [Google Scholar] [CrossRef]
- Lopes, A.R.; Kellam, P.; Das, A.; Dunn, C.; Kwan, A.; Turner, J.; Peppa, D.; Gilson, R.J.; Gehring, A.; Bertoletti, A.; et al. Bim-mediated deletion of antigen-specific CD8 T cells in patients unable to control HBV infection. J. Clin. Investig. 2008, 118, 1835–1845. [Google Scholar] [CrossRef] [PubMed]
- Maini, M.K.; Burton, A.R. Restoring, releasing or replacing adaptive immunity in chronic hepatitis B. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 662–675. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Chen, Y.; Lu, M. Advances in Targeting the Innate and Adaptive Immune Systems to Cure Chronic Hepatitis B Virus Infection. Front. Immunol. 2019, 10, 3127. [Google Scholar] [CrossRef] [PubMed]
- Marraco, S.A.F.; Neubert, N.J.; Verdeil, G.; Speiser, D.E. Inhibitory Receptors Beyond T Cell Exhaustion. Front. Immunol. 2015, 6, 310. [Google Scholar]
- Yuen, M.F.; Asselah, T.; Jacobson, I.M.; Brunetto, M.R.; Janssen, H.L.; Takehara, T.; Hou, J.L.; Kakuda, T.N.; Lambrecht, T.; Beumont, M.; et al. Efficacy and safety of the siRNA JNJ-73763989 and the capsid assembly modulator JNJ-56136379 (bersacapavir) with nucleos(t)ide analogues for the treatment of chronic hepatitis B virus infection (REEF-1): A multicentre, double-blind, active-controlled, randomised, phase 2b trial. Lancet Gastroenterol. Hepatol. 2023, 8, 790–802. [Google Scholar] [PubMed]
- Yuen, M.-F. Repeat dosing of the GalNAc-siRNA AB-729 in subjects with chronic hepatitis B results in robust and sustained HBsAg suppression. J. Hepatol. 2021, 75, S201–S293. [Google Scholar]
- Sandra, L.; T’jollyn, H.; Goeyvaerts, N.; Vermeulen, A.; Dosne, A.-G.; Perez-Ruixo, J.-J. Plasma and Liver Pharmacokinetics of the N-Acetylgalactosamine Short Interfering RNA JNJ-73763989 in Recombinant Adeno-Associated-Hepatitis B Virus-Infected Mice. J. Pharmacol. Exp. Ther. 2022, 383, 70–79. [Google Scholar] [CrossRef]
- Xu, R.; Njumbe Ediage, E.; Laurijssen, G.; Verhaeghe, T.; Snoeys, J.; Dillen, D. RISC-PK Profile of 5′p-Antisense Strand in Wildtype Mouse Shows Good Correlation with RISC-PK/PD in AAV-HBV Mouse Model; Oligonucleotide Therapeutics Society: Barcelona, Spain, 2023. [Google Scholar]
- Heim, K.; Binder, B.; Sagar; Wieland, D.; Hensel, N.; Llewellyn-Lacey, S.; Gostick, E.; Price, D.A.; Emmerich, F.; Vingerhoet, H.; et al. TOX defines the degree of CD8+ T cell dysfunction in distinct phases of chronic HBV infection. Gut 2020, 70, 1550–1560. [Google Scholar] [CrossRef] [PubMed]
- Gane, E. The roadmap towards cure of chronic hepatitis B virus infection. J. R. Soc. N. Z. 2022, 52, 129–148. [Google Scholar] [CrossRef]
- Pan, Y.; Xia, H.; He, Y.; Zeng, S.; Shen, Z.; Huang, W. The progress of molecules and strategies for the treatment of HBV infection. Front. Cell. Infect. Microbiol. 2023, 13, 1128807. [Google Scholar] [CrossRef] [PubMed]
- Gane, E.; Yuen, M.; Kim, D.J.; Chan, H.L.; Surujbally, B.; Pavlovic, V.; Das, S.; Triyatni, M.; Kazma, R.; Grippo, J.F.; et al. Clinical study of single-stranded oligonucleotide RO7062931 in healthy volunteers and patients with chronic hepatitis B. Hepatology 2021, 74, 1795–1808. [Google Scholar] [CrossRef] [PubMed]
- Paratala, B.; Park, J.; Ganchua, S.; Gane, E.; Yuen, M.F.; Lee, A.; Moore, C.; Lam, A.M.; Sevinsky, H.; Sims, K. Inhibition of hepatitis B surface antigen in chronic hepatitis B subjects by RNA interference therapeutic AB-729 is accompanied by upregulation of HBV-specific T cell activation markers. In Proceedings of the International Liver Congress 2021 (ILC 2021), Virtual, 23–26 June 2021; Elsevier BV: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Streinu-Cercel, A.; Gane, E.; Cheng, W.; Sievert, W.; Roberts, S.; Ahn, S.; Kim, Y.; Agarwal, K.; Niforos, D.; Symonds, B.; et al. A phase 2a study evaluating the multi-dose activity of ARB-1467 in HBeAg positive and negative virally suppressed subjects with hepatitis B. J. Hepatol. 2017, 66, S688–S689. [Google Scholar] [CrossRef]
- Thi, E.P.; Yuen, R.M.F.; Gane, E.; Sevinsky, H.; Sims, K.; Anderson, M.; Lam, A.M.; Sofia, M.J.; Cloherty, G.; Picchio, G. Inhibition of hepatitis B surface antigen by RNA interference therapeutic AB-729 in chronic hepatitis B patients correlates with suppression of all HBsAg isoforms and HBV RNA. In Proceedings of the International Liver Congress 2021 (ILC 2021), Virtual, 23–26 June 2021; Elsevier BV: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Yuen, M.F.; Lim, T.H.; Kim, W.; Tangkijvanich, P.; Yoon, J.H.; Sievert, W. HBV RNAi inhibitor RG6346 in Phase 1b-2a trial was safe, well-tolerated, and resulted in substantial and durable reductions in serum HBsAg levels. In Proceedings of the Liver Meeting Digital Experience™, Virtual, 13–16 November 2020. [Google Scholar]
- Bertoletti, A.; Ferrari, C. Adaptive immunity in HBV infection. J. Hepatol. 2016, 64 (Suppl. S1), S71–S83. [Google Scholar] [CrossRef] [PubMed]
- Burton, A.R.; Pallett, L.J.; McCoy, L.E.; Suveizdyte, K.; Amin, O.E.; Swadling, L.; Alberts, E.; Davidson, B.R.; Kennedy, P.T.; Gill, U.S.; et al. Circulating and intrahepatic antiviral B cells are defective in hepatitis B. J. Clin. Investig. 2018, 128, 4588–4603. [Google Scholar] [CrossRef]
- Isogawa, M.; Chung, J.; Murata, Y.; Kakimi, K.; Chisari, F.V. CD40 activation rescues antiviral CD8(+) T cells from PD-1-mediated exhaustion. PLoS Pathog. 2013, 9, e1003490. [Google Scholar] [CrossRef]
- Maini, M.K.; Gehring, A.J. The role of innate immunity in the immunopathology and treatment of HBV infection. J. Hepatol. 2016, 64 (Suppl. S1), S60–S70. [Google Scholar] [CrossRef]
- Mueller, S.N.; Ahmed, R. High antigen levels are the cause of T cell exhaustion during chronic viral infection. Proc. Natl. Acad. Sci. USA 2009, 106, 8623–8628. [Google Scholar] [CrossRef]
- Richter, K.; Brocker, T.; Oxenius, A. Antigen amount dictates CD8+ T-cell exhaustion during chronic viral infection irrespective of the type of antigen presenting cell. Eur. J. Immunol. 2012, 42, 2290–2304. [Google Scholar] [CrossRef]
- Schietinger, A.; Greenberg, P.D. Tolerance and exhaustion: Defining mechanisms of T cell dysfunction. Trends Immunol. 2014, 35, 51–60. [Google Scholar] [CrossRef]
- Tay, S.S.; Wong, Y.C.; McDonald, D.M.; Wood, N.A.W.; Roediger, B.; Sierro, F.; Mcguffog, C.; Alexander, I.E.; Bishop, G.A.; Gamble, J.R.; et al. Antigen expression level threshold tunes the fate of CD8 T cells during primary hepatic immune responses. Proc. Natl. Acad. Sci. USA 2014, 111, E2540–E2549. [Google Scholar] [CrossRef]
- Wherry, E.J. T cell exhaustion. Nat. Immunol. 2011, 12, 492–499. [Google Scholar] [CrossRef]
- Hendrikx, T.; Porsch, F.; Kiss, M.G.; Rajcic, D.; Papac-Miličević, N.; Hoebinger, C.; Goederle, L.; Hladik, A.; Shaw, L.E.; Horstmann, H.; et al. Soluble TREM2 levels reflect the recruitment and expansion of TREM2(+) macrophages that localize to fibrotic areas and limit NASH. J. Hepatol. 2022, 77, 1373–1385. [Google Scholar] [CrossRef]
- Coelho, I.; Duarte, N.; Barros, A.; Macedo, M.P.; Penha-Gonçalves, C. Trem-2 Promotes Emergence of Restorative Macrophages and Endothelial Cells During Recovery from Hepatic Tissue Damage. Front. Immunol. 2020, 11, 616044. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group A | Group B | # Significant Genes (Adjusted p-Value < 0.05) |
|---|---|---|
| G1 High titer control: Control siRNA + Control siRNA (n = 6) | G4 Mid titer control: Control siRNA + Control siRNA (n = 5) | 0 |
| G1 High titer control: Control siRNA + Control siRNA (n = 6) | G2 High titer single treatment: Control siRNA + HBV siRNA (n = 13) | 5 |
| G1 High titer control: Control siRNA + Control siRNA (n = 6) | G3 High titer retreatment: HBV siRNA + HBV siRNA (n = 12) | 72 |
| G2 High titer single treatment: Control siRNA + HBV siRNA (n = 13) | G3 High titer retreatment: HBV siRNA + HBV siRNA (n = 12) | 0 |
| G4 Mid titer control: Control siRNA + Control siRNA (n = 5) | G5 Mid titer single treatment: Control siRNA + HBV siRNA (n = 6) | 3 |
| G4 Mid titer control: Control siRNA + Control siRNA (n = 5) | G6 Mid titer retreatment: HBV siRNA + HBV siRNA (n = 6) | 425 |
| G5 Mid titer single treatment: Control siRNA + HBV siRNA (n = 6) | G6 Mid titer retreatment: HBV siRNA + HBV siRNA (n = 6) | 196 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Gulck, E.; Conceição-Neto, N.; Aerts, L.; Pierson, W.; Verschueren, L.; Vleeschouwer, M.; Krishna, V.; Nájera, I.; Pauwels, F. Retreatment with HBV siRNA Results in Additional Reduction in HBV Antigenemia and Immune Stimulation in the AAV-HBV Mouse Model. Viruses 2024, 16, 347. https://doi.org/10.3390/v16030347
Van Gulck E, Conceição-Neto N, Aerts L, Pierson W, Verschueren L, Vleeschouwer M, Krishna V, Nájera I, Pauwels F. Retreatment with HBV siRNA Results in Additional Reduction in HBV Antigenemia and Immune Stimulation in the AAV-HBV Mouse Model. Viruses. 2024; 16(3):347. https://doi.org/10.3390/v16030347
Chicago/Turabian StyleVan Gulck, Ellen, Nádia Conceição-Neto, Liese Aerts, Wim Pierson, Lore Verschueren, Mara Vleeschouwer, Vinod Krishna, Isabel Nájera, and Frederik Pauwels. 2024. "Retreatment with HBV siRNA Results in Additional Reduction in HBV Antigenemia and Immune Stimulation in the AAV-HBV Mouse Model" Viruses 16, no. 3: 347. https://doi.org/10.3390/v16030347
APA StyleVan Gulck, E., Conceição-Neto, N., Aerts, L., Pierson, W., Verschueren, L., Vleeschouwer, M., Krishna, V., Nájera, I., & Pauwels, F. (2024). Retreatment with HBV siRNA Results in Additional Reduction in HBV Antigenemia and Immune Stimulation in the AAV-HBV Mouse Model. Viruses, 16(3), 347. https://doi.org/10.3390/v16030347