


First Detection of Benign Rabbit Caliciviruses in Chile
 , , , , , and
, , , , , and 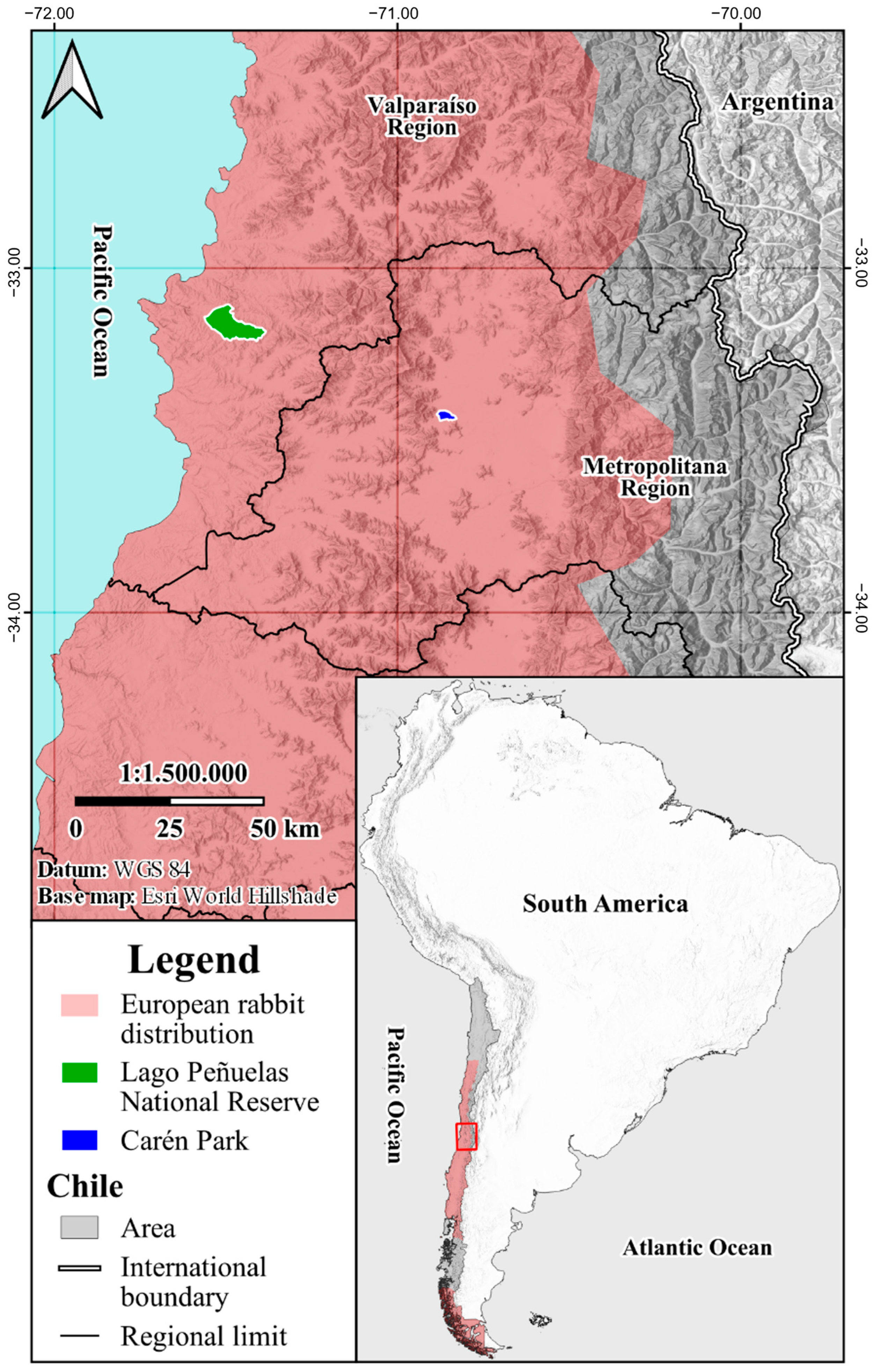{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Sample Collection
2.2. RNA Extraction and Virus Detection
2.3. Sequencing and Data Analysis
2.4. Phylogenetic Analysis
2.5. Phylogenetic Molecular Clock Analysis
3. Results
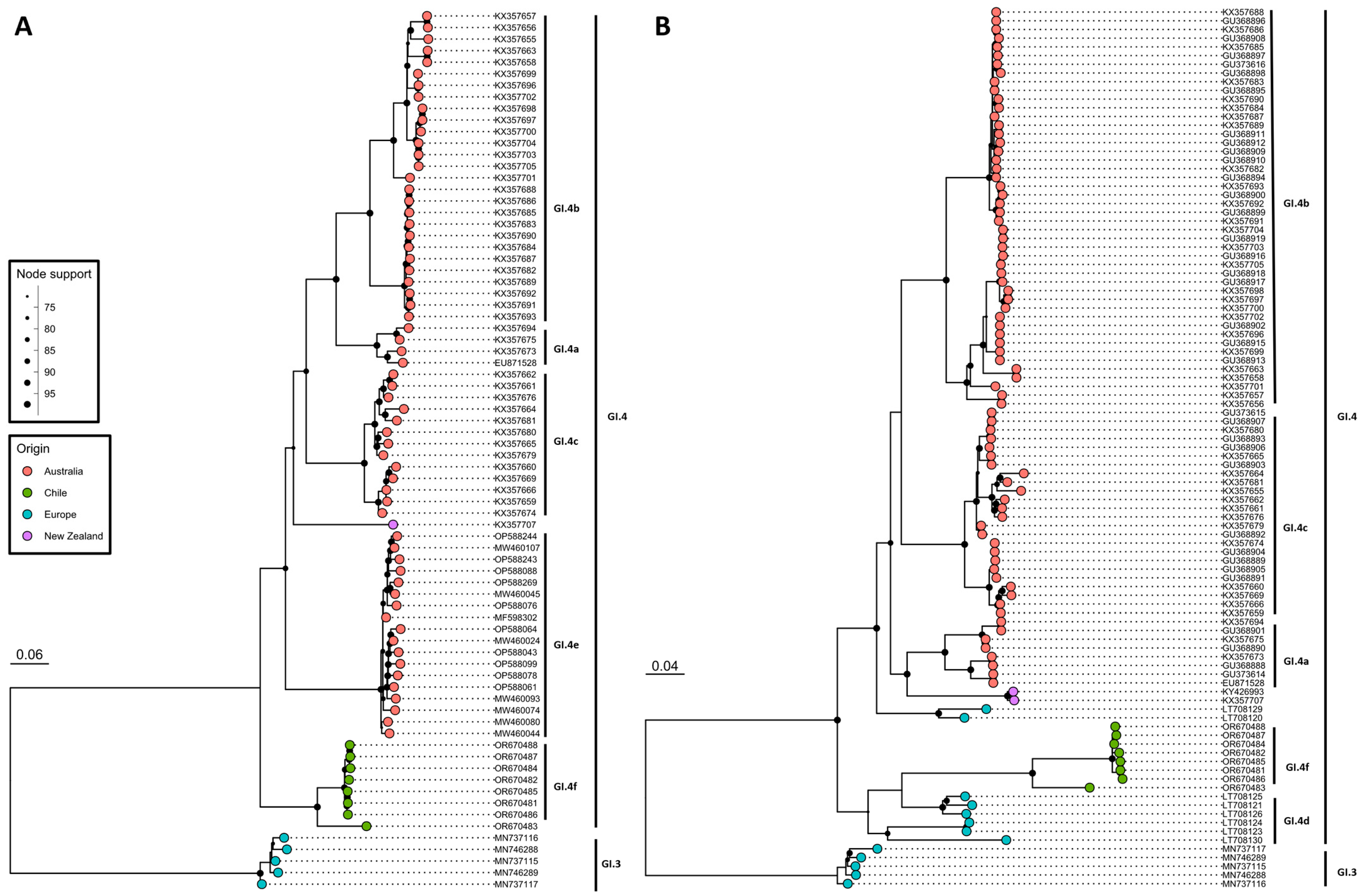
3.1. Detection and Phylogenetic Analysis of Benign Rabbit Caliciviruses
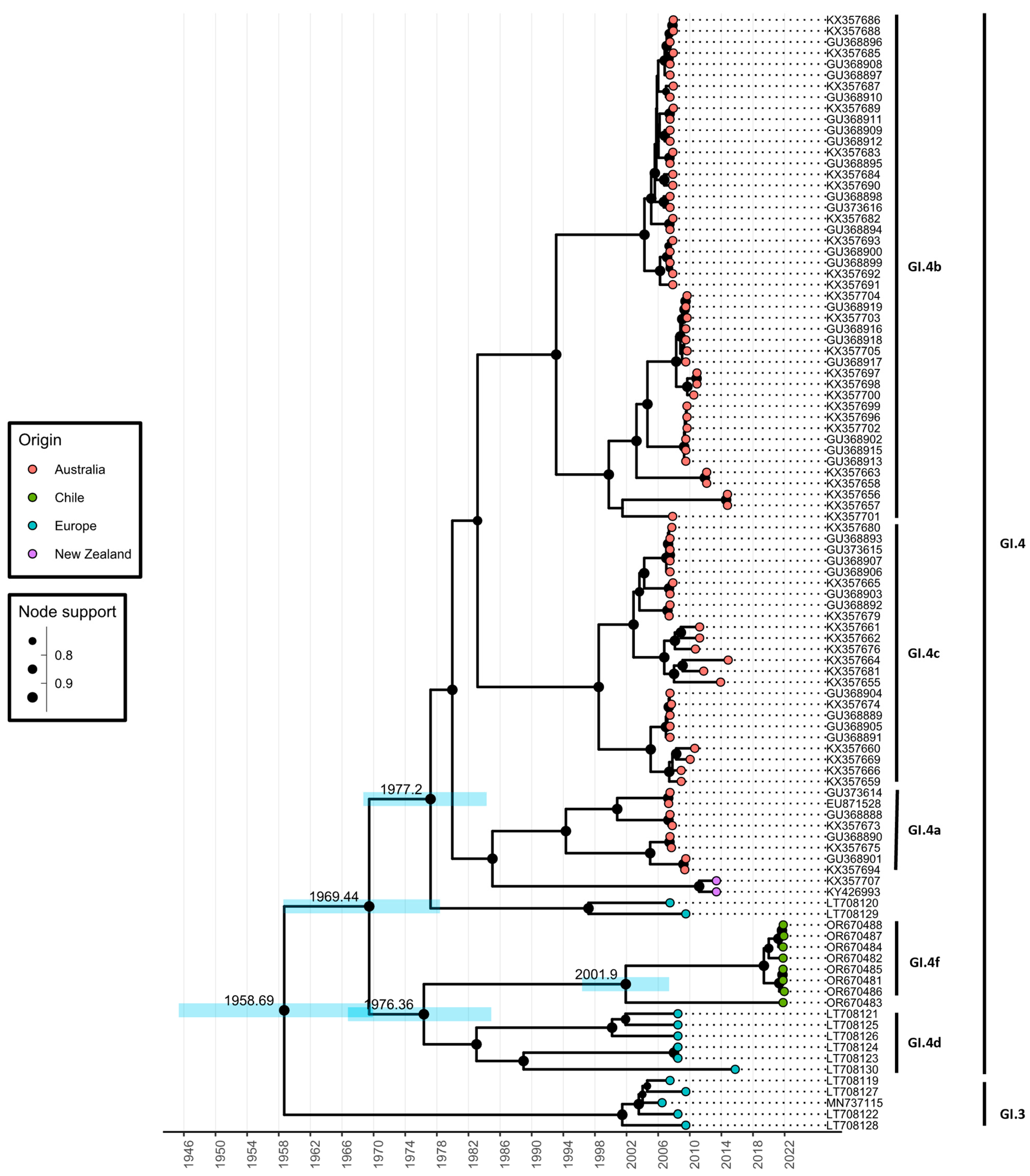
3.2. Time Scale Estimate of GI.4f Evolution
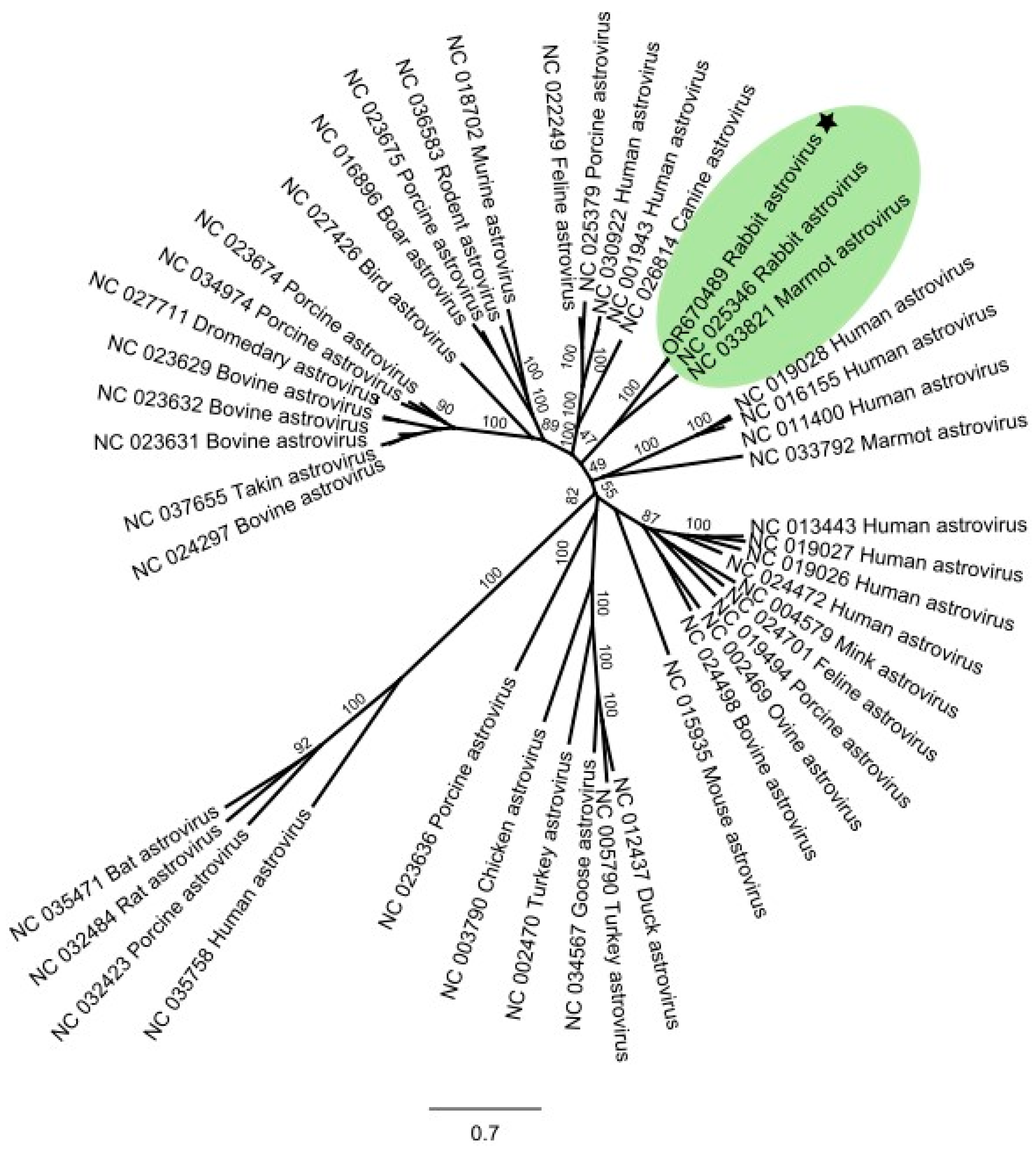
3.3. Coincidental Find of a Rabbit Astrovirus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Capucci, L.; Fusi, P.; Lavazza, A.; Pacciarini, M.L.; Rossi, C. Detection and Preliminary Characterization of a New Rabbit Calicivirus Related to Rabbit Hemorrhagic Disease Virus but Nonpathogenic. J. Virol. 1996, 70, 8614–8623. [Google Scholar] [CrossRef] [PubMed]
- Strive, T.; Wright, J.D.; Robinson, A.J. Identification and Partial Characterisation of a New Lagovirus in Australian Wild Rabbits. Virology 2009, 384, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Strive, T.; Wright, J.; Kovaliski, J.; Botti, G.; Capucci, L. The Non-Pathogenic Australian Lagovirus RCV-A1 Causes a Prolonged Infection and Elicits Partial Cross-Protection to Rabbit Haemorrhagic Disease Virus. Virology 2010, 398, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Hall, R.N.; Strive, T. Viruses for Landscape-Scale Therapy: Biological Control of Rabbits in Australia. Methods Mol. Biol. 2021, 2225, 1–23. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Zwingelstein, F.; Fages, M.P.; Bertagnoli, S.; Gelfi, J.; Aubineau, J.; Roobrouck, A.; Botti, G.; Lavazza, A.; Marchandeau, S. Characterisation of a Non-Pathogenic and Non-Protective Infectious Rabbit Lagovirus Related to RHDV. Virology 2011, 410, 395–402. [Google Scholar] [CrossRef]
- Lemaitre, E.; Zwingelstein, F.; Marchandeau, S.; Le Gall-Reculé, G. First Complete Genome Sequence of a European Non-Pathogenic Rabbit Calicivirus (Lagovirus GI.3). Arch. Virol. 2018, 163, 2921–2924. [Google Scholar] [CrossRef]
- Liu, J.; Fordham, D.A.; Cooke, B.D.; Cox, T.; Mutze, G.; Strive, T. Distribution and Prevalence of the Australian Non-Pathogenic Rabbit Is Correlated with Rainfall and Temperature. PLoS ONE 2014, 9, e113976. [Google Scholar] [CrossRef]
- Nicholson, L.J.; Mahar, J.E.; Strive, T.; Zheng, T.; Holmes, E.C.; Ward, V.K.; Duckworth, J.A. Benign Rabbit Calicivirus in New Zealand. Appl. Environ. Microbiol. 2017, 83, e00090-17. [Google Scholar] [CrossRef] [PubMed]
- Ehresmann, D.W.; Schaffer, F.L. RNA Synthesized in Calicivirus Infected Cells Is Atypical of Picornaviruses. J. Virol. 1977, 22, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Meyers, G.; Wirblich, C.; Thiel, H.-J. Rabbit Hemorrhagic Disease Virus—Molecular Cloning and Nucleotide Sequencing of a Calicivirus Genome. Virology 1991, 184, 664–676. [Google Scholar] [CrossRef]
- Meyers, G.; Wirblich, C.; Thiel, H.J. Genomic and Subgenomic RNAs of Rabbit Hemorrhagic Disease Virus Are Both Protein-Linked and Packaged into Particles. Virology 1991, 184, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Parra, F.; Prieto, M. Purification and Characterization of a Calicivirus as the Causative Agent of a Lethal Hemorrhagic Disease in Rabbits. J. Virol. 1990, 64, 4013–4015. [Google Scholar] [CrossRef]
- Conley, M.J.; McElwee, M.; Azmi, L.; Gabrielsen, M.; Byron, O.; Goodfellow, I.G.; Bhella, D. Calicivirus VP2 Forms a Portal-like Assembly Following Receptor Engagement. Nature 2019, 565, 377–381. [Google Scholar] [CrossRef]
- Black, D.N.; Burroughs, J.N.; Harris, T.J.R.; Brown, F. The Structure and Replication of Calicivirus RNA. Nature 1978, 274, 614–615. [Google Scholar] [CrossRef]
- Burroughs, J.N.; Brown, F. Presence of a Covalently Linked Protein on Calicivirus RNA. J. Gen. Virol. 1978, 41, 443–446. [Google Scholar] [CrossRef]
- Le Pendu, J.; Abrantes, J.; Bertagnoli, S.; Guitton, J.S.; Le Gall-Reculé, G.; Lopes, A.M.; Marchandeau, S.; Alda, F.; Almeida, T.; Célio, A.P.; et al. Proposal for a Unified Classification System and Nomenclature of Lagoviruses. J. Gen. Virol. 2017, 98, 1658–1666. [Google Scholar] [CrossRef]
- Mahar, J.E.; Jenckel, M.; Huang, N.; Smertina, E.; Holmes, E.C.; Strive, T.; Hall, R.N. Frequent Intergenotypic Recombination between the Non-Structural and Structural Genes Is a Major Driver of Epidemiological Fitness in Caliciviruses. Virus Evol. 2021, 7, veab080. [Google Scholar] [CrossRef]
- Lopes, A.M.; Dalton, K.P.; Magalhães, M.J.; Parra, F.; Esteves, P.J.; Holmes, E.C.; Abrantes, J. Full Genomic Analysis of New Variant Rabbit Hemorrhagic Disease Virus Revealed Multiple Recombination Events. J. Gen. Virol. 2015, 96, 1309–1319. [Google Scholar] [CrossRef]
- Abrantes, J.; Lopes, A.M.; Lemaitre, E.; Ahola, H.; Banihashem, F.; Droillard, C.; Marchandeau, S.; Esteves, P.J.; Neimanis, A.; Gall-Reculé, G. Le Retrospective Analysis Shows That Most Rhdv Gi.1 Strains Circulating since the Late 1990s in France and Sweden Were Recombinant Gi.3p–Gi.1d Strains. Genes 2020, 11, 910. [Google Scholar] [CrossRef]
- Abrantes, J.; Droillard, C.; Lopes, A.M.; Lemaitre, E.; Lucas, P.; Blanchard, Y.; Marchandeau, S.; Esteves, P.J.; Le Gall-Reculé, G. Recombination at the Emergence of the Pathogenic Rabbit Haemorrhagic Disease Virus Lagovirus Europaeus/GI.2. Sci. Rep. 2020, 10, 14502. [Google Scholar] [CrossRef]
- Urakova, N.; Hall, R.; Strive, T.; Frese, M. Restricted Host Specificity of Rabbit Hemorrhagic Disease Virus Is Supported by Challenge Experiments in Immune-Compromised Mice (Mus musculus). J. Wildl. Dis. 2019, 55, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Cooke, B.D.; Duncan, R.P.; McDonald, I.; Liu, J.; Capucci, L.; Mutze, G.J.; Strive, T. Prior Exposure to Non-Pathogenic Calicivirus RCV-A1 Reduces Both Infection Rate and Mortality from Rabbit Haemorrhagic Disease in a Population of Wild Rabbits in Australia. Transbound. Emerg. Dis. 2018, 65, e470–e477. [Google Scholar] [CrossRef]
- Strive, T.; Elsworth, P.; Liu, J.; Wright, J.D.; Kovaliski, J.; Capucci, L. The Non-Pathogenic Australian Rabbit Calicivirus RCV-A1 Provides Temporal and Partial Cross Protection to Lethal Rabbit Haemorrhagic Disease Virus Infection Which Is Not Dependent on Antibody Titres. Vet. Res. 2013, 44, 51. [Google Scholar] [CrossRef]
- Taggart, P.L.; Hall, R.N.; Cox, T.E.; Kovaliski, J.; McLeod, S.R.; Strive, T. Changes in Virus Transmission Dynamics Following the Emergence of RHDV2 Shed Light on Its Competitive Advantage over Previously Circulating Variants. Transbound. Emerg. Dis. 2022, 69, 1118–1130. [Google Scholar] [CrossRef]
- Camus, P.; Castro, S.; Jaksic, F. European Rabbits in Chile: The History of a Biological Invasion. Hist. Santiago 2008, 41, 305–339. [Google Scholar] [CrossRef]
- Camus, P.; Castro, S.A.; Jaksic, F.M. European Rabbit (Oryctolagus cuniculus L.) in Chile: The Human Dimension behind a Biological Invasion. In Biological Invasions in the South American Anthropocene: Global Causes and Local Impacts; Springer: Cham, Switzerland, 2021; pp. 171–185. [Google Scholar] [CrossRef]
- Correa-Cuadros, J.P.; Flores-Benner, G.; Muñoz-Rodríguez, M.A.; Briceño, C.; Díaz, M.; Strive, T.; Vásquez, F.; Jaksic, F.M. History, Control, Epidemiology, Ecology, and Economy of the Invasion of European Rabbits in Chile: A Comparison with Australia. Biol. Invasions 2023, 25, 309–338. [Google Scholar] [CrossRef]
- Jaksic, F.M. Vertebrate Invaders and Their Ecological Impacts in Chile. Biodivers. Conserv. 1998, 7, 1427–1445. [Google Scholar] [CrossRef]
- PNUD Valoración Económica Del Impacto de Siete Especies Exóticas Invasoras Sobre Los Sectores Productivos y La Biodiversidad En Chile. Available online: https://especies-exoticas.mma.gob.cl/wp-content/uploads/2018/12/1.-LIBRO-Valoracion-economica-EEI-FINAL.pdf (accessed on 15 January 2024).
- Kerr, P.J.; Cattadori, I.M.; Rogers, M.B.; Fitch, A.; Geber, A.; Liu, J.; Sim, D.G.; Boag, B.; Eden, J.S.; Ghedin, E.; et al. Genomic and Phenotypic Characterization of Myxoma Virus from Great Britain Reveals Multiple Evolutionary Pathways Distinct from Those in Australia. PLoS Pathog. 2017, 13, e1006252. [Google Scholar] [CrossRef]
- Jaksic, F.M.; Yañez, J.L. Rabbit and Fox Introductions in Tierra Del Fuego: History and Assessment of the Attempts at Biological Control of the Rabbit Infestation. Biol. Conserv. 1983, 26, 367–374. [Google Scholar] [CrossRef]
- Kerr, P.J.; Best, S.M. Myxoma Virus in Rabbits. Rev. Sci. Tech. Off. Int. Epiz 1998, 17, 256–268. [Google Scholar] [CrossRef]
- Alves, J.M.; Carneiro, M.; Cheng, J.Y.; Lemos De Matos, A.; Rahman, M.M.; Loog, L.; Campos, P.F.; Wales, N.; Eriksson, A.; Manica, A.; et al. Parallel Adaptation of Rabbit Populations to Myxoma Virus. Science (1979) 2019, 363, 1319–1326. [Google Scholar] [CrossRef]
- Cooke, B.D.; Fenner, F. Rabbit Haemorrhagic Disease and the Biological Control of Wild Rabbits, Oryctolagus cuniculus, in Australia and New Zealand. Wildl. Res. 2002, 29, 689–706. [Google Scholar] [CrossRef]
- Cooke, B.; Chudleigh, P.; Simpson, S.; Saunders, G. The Economic Benefits of the Biological Control of Rabbits in Australia, 1950–2011. Aust. Econ Hist. Rev. 2013, 53, 91–107. [Google Scholar] [CrossRef]
- Ramsey, D.S.; Patel, K.K.; Campbell, S.; Hall, R.N.; Taggart, P.L.; Strive, T. Sustained Impact of RHDV2 on Wild Rabbit Populations across Australia Eight Years after Its Initial Detection. Viruses 2023, 15, 1159. [Google Scholar] [CrossRef]
- Ramsey, D.S.L.; Cox, T.; Strive, T.; Forsyth, D.M.; Stuart, I.; Hall, R.; Elsworth, P.; Campbell, S. Emerging RHDV2 Suppresses the Impact of Endemic and Novel Strains of RHDV on Wild Rabbit Populations. J. Appl. Ecol. 2020, 57, 630–641. [Google Scholar] [CrossRef]
- Servicio Agrícola y Ganadero (SAG). Lista de enfermedades de denuncia obligatoria (EDO) al SAG. Available online: https://www.sag.gob.cl/sites/default/files/enfermedades_denuncia_obligatoria_sag_9-10-2019.pdf (accessed on 15 January 2024).
- Servicio Agrícola y Ganadero (SAG), Ord. No 274/2015.v1. Letter: Pronunciamiento Sobre La Aplicación de Calicivirus y Virus Mixoma Para Control de Conejo Europeo En Islas de Áreas Silvestres Protegidas Del Estado. Santiago, Chile, 2015. CSIRO Data Collection. Available online: https://data.csiro.au/collection/csiro%3A61523v1 (accessed on 4 February 2024). [CrossRef]
- Donato, C.; Vijaykrishna, D. The Broad Host Range and Genetic Diversity of Mammalian and Avian Astroviruses. Viruses 2017, 9, 102. [Google Scholar] [CrossRef]
- Zhao, Q.; Tian, Y.; Liu, L.; Jiang, Y.; Sun, H.; Tan, S.; Huang, B. The Genomic and Genetic Evolution Analysis of Rabbit Astrovirus. Vet. Sci. 2022, 9, 603. [Google Scholar] [CrossRef]
- American Veterinary Medical Association. AVMA Guidelines for the Euthanasia of Animals: 2020 Edition; American Veterinary Medical Association: Schaumburg, IL, USA, 2020. [Google Scholar]
- Hall, R.N.; Mahar, J.E.; Read, A.J.; Mourant, R.; Piper, M.; Huang, N.; Strive, T. A Strain-Specific Multiplex RT-PCR for Australian Rabbit Haemorrhagic Disease Viruses Uncovers a New Recombinant Virus Variant in Rabbits and Hares. Transbound. Emerg. Dis. 2018, 65, e444–e456. [Google Scholar] [CrossRef]
- Jenckel, M.; Hall, R.N.; Strive, T. Pathogen Profiling of Australian Rabbits by Metatranscriptomic Sequencing. Transbound. Emerg. Dis. 2022, 69, e2629–e2640. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High throughput Sequence Data [Online]. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 4 February 2024).
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Li, D.; Liu, C.M.; Luo, R.; Sadakane, K.; Lam, T.W. MEGAHIT: An Ultra-Fast Single-Node Solution for Large and Complex Metagenomics Assembly via Succinct de Bruijn Graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y. Ggtree: An R Package for Visualization and Annotation of Phylogenetic Trees with Their Covariates and Other Associated Data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Wang, L.G.; Lam, T.T.Y.; Xu, S.; Dai, Z.; Zhou, L.; Feng, T.; Guo, P.; Dunn, C.W.; Jones, B.R.; Bradley, T.; et al. Treeio: An R Package for Phylogenetic Tree Input and Output with Richly Annotated and Associated Data. Mol. Biol. Evol. 2020, 37, 599–603. [Google Scholar] [CrossRef]
- Wickham, H.; François, R.; Henry, L.; Müller, K.; Vaughan, D. Dplyr: A Grammar of Data Manipulation, R Package Version 1.1.2; The R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Wickham, H.; Henry, L.; Pedersen, T.L.; Luciani, T.J.; Decorde, M.; Lise, V. Svglite: An “SVG” Graphics Device, R Package Version 2.1.1; The R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Neuwirth, E. RColorBrewer: ColorBrewer Palettes; RStudio, Inc.: Vienna, Austria, 2022. [Google Scholar]
- Campitelli, E. Ggnewscale: Multiple Fill and Colour Scales in “Ggplot2”; RStudio, Inc.: Vienna, Austria, 2023. [Google Scholar]
- Arnold, J.B. Ggthemes: Extra Themes, Scales and Geoms for “Ggplot2”; RStudio, Inc.: Vienna, Austria, 2023. [Google Scholar]
- Xu, S.; Dai, Z.; Guo, P.; Fu, X.; Liu, S.; Zhou, L.; Tang, W.; Feng, T.; Chen, M.; Zhan, L.; et al. GgtreeExtra: Compact Visualization of Richly Annotated Phylogenetic Data. Mol. Biol. Evol. 2021, 38, 4039–4042. [Google Scholar] [CrossRef]
- Sievert, C. Interactive Web-Based Data Visualization with R, Plotly, and Shiny; CRC Press: Boca Raton, FL, USA, 2020; ISBN 9781138331457. [Google Scholar]
- Barrett, T.; Dowle, M.; Srinivasan, A.; Gorecki, J.; Chirico, M.; Hocking, T. Data.Table: Extension of ‘data.Frame’. R Package Version 1.14.8; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Wickham, H.; Pedersen, T.L.; Seidel, D. Scales: Scale Functions for Visualization; R Package Version 1.2.1; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Rambaut, A.; Lam, T.T.; Carvalho, L.M.; Pybus, O.G. Exploring the Temporal Structure of Heterochronous Sequences Using TempEst (Formerly Path-O-Gen). Virus Evol. 2016, 2, vew007. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian Evolutionary Analysis by Sampling Trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [PubMed]
- Stenglein, M.D.; Velazquez, E.; Greenacre, C.; Wilkes, R.P.; Ruby, J.G.; Lankton, J.S.; Ganem, D.; Kennedy, M.A.; Derisi, J.L. Complete Genome Sequence of an Astrovirus Identified in a Domestic Rabbit (Oryctolagus cuniculus) with Gastroenteritis. Virol. J. 2012, 9, 216. [Google Scholar] [CrossRef]
- Mahar, J.E.; Read, A.J.; Gu, X.; Urakova, N.; Mourant, R.; Piper, M.; Haboury, S.; Holmes, E.C.; Strive, T.; Hall, R.N. Detection and Circulation of a Novel Rabbit Hemorrhagic Disease Virus in Australia. Emerg. Infect. Dis. 2018, 24, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Peng, N.Y.G.; Hall, R.N.; Huang, N.; West, P.; Cox, T.E.; Mahar, J.E.; Mason, H.; Campbell, S.; O’Connor, T.; Read, A.J.; et al. Utilizing Molecular Epidemiology and Citizen Science for the Surveillance of Lagoviruses in Australia. Viruses 2023, 15, 2348. [Google Scholar] [CrossRef] [PubMed]
- Mahar, J.E.; Nicholson, L.; Eden, J.-S.; Duchêne, S.; Kerr, P.J.; Duckworth, J.; Ward, V.K.; Holmes, E.C.; Strive, T. Benign Rabbit Caliciviruses Exhibit Evolutionary Dynamics Similar to Those of Their Virulent Relatives. J. Virol. 2016, 90, 9317–9329. [Google Scholar] [CrossRef]
- Correa-Cuadros, J.P.; Flores-Benner, G.; María, P.G.; Ávila-Thieme, I.; Muñoz, M.; Duclos, M.; Soto, N.; Briceño, C.; Vásquez, F.; Díaz, M.; et al. La Invasión del Conejo Europeo en Chile; Ediciones CAPES-UC: Geneve, Switzerland, 2023; pp. 1–126. [Google Scholar] [CrossRef]
- Arentsen, P. Control Biológico Del Conejo: Difusión Del Virus Mixomatosis cuniculus, Por Contagio Directo, En La Isla Grande de Tierra Del Fuego. Boletín Ganad. (Punta Arenas) 1954, 43, 3–25. [Google Scholar]
- Liu, J.; Kerr, P.J.; Strive, T. A Sensitive and Specific Blocking ELISA for the Detection of Rabbit Calicivirus RCV-A1 Antibodies. Virol. J. 2012, 9, 182. [Google Scholar] [CrossRef]
- Martella, V.; Moschidou, P.; Pinto, P.; Catella, C.; Desario, C.; Larocca, V.; Circella, E.; Bànyai, K.; Lavazza, A.; Magistrali, C.; et al. Astroviruses in Rabbits. Emerg. Infect. Dis. 2011, 17, 2287–2293. [Google Scholar] [CrossRef]
- Esteves, P.J.; Abrantes, J.; Bertagnoli, S.; Cavadini, P.; Gavier-Widén, D.; Guitton, J.S.; Lavazza, A.; Lemaitre, E.; Letty, J.; Lopes, A.M.; et al. Emergence of Pathogenicity in Lagoviruses: Evolution from Pre-Existing Nonpathogenic Strains or through a Species Jump? PLoS Pathog. 2015, 11, e1005087. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smertina, E.; Keller, L.M.; Huang, N.; Flores-Benner, G.; Correa-Cuadros, J.P.; Duclos, M.; Jaksic, F.M.; Briceño, C.; Ramirez, V.N.; Díaz-Gacitúa, M.; et al. First Detection of Benign Rabbit Caliciviruses in Chile. Viruses 2024, 16, 439. https://doi.org/10.3390/v16030439
Smertina E, Keller LM, Huang N, Flores-Benner G, Correa-Cuadros JP, Duclos M, Jaksic FM, Briceño C, Ramirez VN, Díaz-Gacitúa M, et al. First Detection of Benign Rabbit Caliciviruses in Chile. Viruses. 2024; 16(3):439. https://doi.org/10.3390/v16030439
Chicago/Turabian StyleSmertina, Elena, Luca M. Keller, Nina Huang, Gabriela Flores-Benner, Jennifer Paola Correa-Cuadros, Melanie Duclos, Fabian M. Jaksic, Cristóbal Briceño, Victor Neira Ramirez, Miguel Díaz-Gacitúa, and et al. 2024. "First Detection of Benign Rabbit Caliciviruses in Chile" Viruses 16, no. 3: 439. https://doi.org/10.3390/v16030439
APA StyleSmertina, E., Keller, L. M., Huang, N., Flores-Benner, G., Correa-Cuadros, J. P., Duclos, M., Jaksic, F. M., Briceño, C., Ramirez, V. N., Díaz-Gacitúa, M., Carrasco-Fernández, S., Smith, I. L., Strive, T., & Jenckel, M. (2024). First Detection of Benign Rabbit Caliciviruses in Chile. Viruses, 16(3), 439. https://doi.org/10.3390/v16030439